











 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCCIÓN
El reclutamiento se define como la incorporación de nuevos individuos a una población y es un proceso demográfico esencial vinculado al mantenimiento de las poblaciones silvestres (Caley et al., 1996). En el caso de organismos como los árboles y corales, que son los respectivos componentes estructurales fundamentales de bosques y arrecifes coralinos, el reclutamiento también determina la resiliencia de estos ecosistemas (Hughes y Tanner, 2000). La comprensión del reclutamiento de corales es primordial dado el evidente deterioro global de las últimas décadas en los arrecifes de coral y sus servicios asociados (Richmond, 1997; Babcock et al., 2003; Hughes et al., 2010).
El reclutamiento de corales se ha documentado relativamente bien en las regiones del Indo-Pacífico, Pacífico Central y Caribe, y ha sido menos estudiado en el Pacífico Tropical Oriental - PTO (Glynn et al., 2017a). Los primeros estudios en esta región no encontraron reclutas de corales sobre sustratos naturales en Panamá, y muestreos realizados con placas de asentamiento resultaron en un reclutamiento extremadamente bajo o nulo (Birkeland, 1977; Wellington, 1982; Richmond, 1985, 1987). Estos resultados negativos, además de aquellos de los primeros análisis histológicos, llevaron incluso a algunos investigadores a considerar a los corales del PTO como "un ejemplo de población estéril o expatriada" (Richmond, 1985, 1987), cuya recuperación de las perturbaciones era producto principalmente de la reproducción asexual, especialmente por fragmentación de colonias (Highsmith, 1982; Muñoz et al., 2015). Estudios más recientes han encontrado pocos -si es que han encontrado alguno- reclutas en las placas de asentamiento, por ejemplo: nueve reclutas de Porites en el Pacífico mexicano (Medina- Rosas et al., 2005), 291 reclutas de Porites panamensis, pero sólo un recluta de Pocillopora también en el Pacífico mexicano (López-Pérez et al., 2007), y ningún recluta en la isla Gorgona, Pacífico colombiano (Lozano-Cortés y Zapata, 2014). Además, los nuevos análisis histológicos indicaban claramente que la maduración gonadal era un proceso activo (Glynn et al., 1991, 1994, 2017a y sus referencias), incluso en los corales pocilopóridos de isla Gorgona (Castrillón et al., 2015).
Aunque la isla Gorgona se encuentra dentro de un área marina protegida en la que prácticamente no existen impactos antropogénicos directos, estos arrecifes se ven afectados negativamente por perturbaciones naturales recurrentes como el calentamiento temporal del agua por el fenómeno de El Niño-Oscilación del Sur y las exposiciones al aire durante mareas extremadamente bajas (Zapata y Vargas-Ángel, 2003; Castrillón et al., 2017; Fiedler y Lavín, 2017). Por ejemplo, la cobertura de coral vivo en el arrecife de La Azufrada disminuyó en promedio 26% durante la década 1998-2008, siendo aún más crítico en las áreas someras del arrecife donde pasó de 61% a 15% (Zapata, 2017), a partir de donde ha ido recuperándose lentamente. Para entender apropiadamente estos cambios en el ecosistema, es urgente conocer en qué medida contribuye el reclutamiento de los corales a la resiliencia de estos arrecifes; sin embargo, una dificultad importante para lograrlo ha sido la falta de evidencia de que el reclutamiento coralino producto de la reproducción sexual ocurre en la isla.
En este estudio, se presenta evidencia del reclutamiento de corales producto del asentamiento de larvas en sustratos naturales de arrecifes coralinos y sitios rocosos de la isla Gorgona, registrando al mismo tiempo la ausencia de reclutas de coral en placas de asentamiento de diferentes materiales. También se registra la distribución de frecuencia de tamaños y la densidad poblacional de colonias juveniles de Pocillopora sp. y Porites panamensis.
MÉTODOS
Área de estudio
Isla Gorgona se encuentra ubicada a unos 30 km de la costa (3° 00' 55''N, 78° 14' 30''O) y constituye el territorio insular más grande (13.2 km2) de la plataforma continental del Pacífico colombiano. La isla y sus aguas circundantes son parte del Parque Nacional Natural Gorgona, que alberga algunas de las formaciones de arrecifes de coral más grandes, diversas y desarrolladas del PTO, junto con extensas costas rocosas intermareales y montañas rocosas sumergidas (Zapata y Vargas-Ángel, 2003; Muñoz y Zapata, 2013). Hasta la fecha, se conocen alrededor de 25 especies de corales hermatípicos en la isla, pero debido a las dificultades taxonómicas dentro del género Pocillopora (Schmidt-Roach et al., 2014), el número verdadero aún se desconoce (Glynn et al., 2017b). Los corales pocilopóridos dominan en las aguas poco profundas (< 5 m de profundidad) y son los principales constructores de los verdaderos arrecifes coralinos, mientras que las especies de crecimiento masivo como Pavona varians, Porites lobata y Gardineroseris planulata están relegadas a los bordes exteriores de esos arrecifes pocilopóridos, siendo más comunes en sitios rocosos más profundos. Las especies de coral más frecuentes en isla Gorgona se consideran liberadoras de gametos, excepto por Porites panamensis que es incubadora, y por las especies de Pocillopora, las cuales han mostrado diversas formas de reproducción en todo el mundo (Glynn et al., 1991).
La precipitación anual promedio es alta, oscilando entre 4000 - 7000 mm con dos estaciones climatológicas contrastantes, una lluviosa de mayo a octubre y otra seca de diciembre a marzo (Díaz et al., 2001; Blanco, 2009). Debido a las fuertes lluvias en Gorgona, las cuerpos de agua dulce son abundantes y la turbidez del mar es muy variable y está parcialmente relacionada con la lluvia. El régimen de mareas es semidiurno, con un ámbito máximo de marea de 5.7 m (IDEAM, 2017); las corrientes oceánicas y la acción del oleaje son más fuertes en el lado oeste, barlovento de la isla principal (Giraldo et al., 2008).
Muestreo
Se implementaron tres métodos diferentes para estudiar el reclutamiento coralino producto de la reproducción sexual en Isla Gorgona: A) una amplia búsqueda de corales juveniles en los sustratos naturales alrededor de la isla durante inmersiones exploratorias, B) conteos visuales de los corales juveniles en transectos, y C) el uso de placas de asentamiento para muestrear reclutas de coral.
Inmersiones exploratorias
Durante las inmersiones exploratorias en busca de corales juveniles, se inspeccionaron un total de 19 sitios alrededor de la isla, incluyendo arrecifes de coral y sitios rocosos. Se realizaron al menos tres inmersiones en cada sitio al año; se examinaron 17 sitios durante 2010 - 2015, y dos sitios adicionales durante 2017.
Se utilizaron diferentes criterios (basados en Richmond, 1985 y Glynn et al., 1996) para determinar si las pequeñas colonias de coral eran el resultado del asentamiento de larvas, particularmente en los pocilopóridos ramificados que se fragmentan con frecuencia (Highsmith, 1982; Glynn et al., 1991; Muñoz et al., 2015). Los criterios fueron: 1) La presencia de una base incrustante ancha, plana y casi simétrica, 2) la fijación a sustratos empinados o verticales, como las paredes rocosas, donde no se esperaría encontrar fragmentos de coral, y 3) la ubicación de los corales juveniles suficientemente alejados de las colonias adultas. El cumplimiento simultáneo de algunos criterios para la identificación de corales juveniles ha supuesto un fuerte apoyo frente a las críticas de algunos autores de que la inspección visual de los corales juveniles no es suficiente para establecer el origen sexual (por asentamiento de una larva) o asexual (por supervivencia de un fragmento) de una pequeña colonia de coral (ej., Miller et al., 2008).
Similar a Edmunds (2004), un criterio usado para definir a los corales juveniles fue un tamaño de colonia entre ~ 0.1 cm (1 pólipo) y ≤ 5.0 cm de diámetro, incluyendo por lo tanto reclutas y corales juveniles. Considerando que los corales pocilopóridos alcanzan la madurez sexual alrededor de los 2-3 años y crecen ~ 2.7 cm a-1 (Palacios et al., 2014), una colonia de 5 cm de diámetro tendría 1.5 años de edad. Del mismo modo, los corales masivos como Porites panamensis tendrían aproximadamente 3.5 años de edad, considerando que alcanzan la madurez a los 4-7 años (Richmond, 1997) y crecen ~ 1.5 cm a-1 (Cabral-Tena et al., 2013). Finalmente, debido a que los corales juveniles pocilopóridos fueron difíciles de identificar a nivel de especie, se clasificaron como Pocillopora sp.
Conteo en transectos
Para estimar la abundancia y el tamaño (diámetro máximo) de las especies de corales juveniles, durante noviembre de 2010 se examinaron diez transectos visuales (cada transecto de 20 m2) en cada uno de los dos arrecifes de coral más grandes de la isla (La Azufrada y Playa Blanca); además, durante marzo de 2011 se examinaron nueve transectos similares en la costa rocosa de El Laberinto; y durante junio de 2011, se examinaron tres transectos más pequeños (10 m2) en el arrecife coralino El Arrecifito. Para evaluar la abundancia y las diferencias de tamaño entre los corales juveniles de pocilopóridos de diferentes sitios, se utilizó ANOVA de una vía, después de comprobar que se cumplían los supuestos de homocedasticidad y normalidad.
Placas de asentamiento
En octubre de 2010, se instalaron en total 54 placas de asentamiento de ~ 20 x 20 x 0.5 cm (~ 840 cm2 de superficie total por placa) en la planicie arrecifal (~ 0.5 a 1 m de profundidad en relación con el promedio del nivel del agua en marea baja) del arrecife de La Azufrada (12 placas de cada uno de los cuatro materiales: acrílico, cerámica, concreto y mármol, y seis placas adicionales de hilo de nylon trenzado y entretejido sirviendo de superficie en un marco de tubos de polivinilo). Las placas de los cuatro materiales acrílico, cerámica, concreto y mármol se instalaron en pares, una encima de la otra separadas aproximadamente por 2 cm, para proporcionar entre el par de placas un microhábitat críptico a menudo preferido para el asentamiento de las larvas de coral (Harriott y Fisk, 1987); todos los pares de placas se mantuvieron unidos por una barra de acero inoxidable clavada en el sustrato. Para reducir los efectos potenciales sobre los reclutas de coral debido a la competencia por el espacio con algas y cirrípedos (Birkeland, 1977), estos organismos se eliminaron raspando las superficies de la mitad de las placas (27 placas elegidas al azar y marcadas) después de revisarlas y fotografiarlas, durante visitas al sitio de estudio aproximadamente cada 45 días. Todas las placas fueron examinadas directamente y fotografiadas con una cámara digital de alta resolución con la misma frecuencia por un período de seis meses; las fotografías fueron finalmente examinadas visualmente en un computador. En abril de 2011, se extrajeron las placas del mar y se examinaron directamente en el laboratorio con un microscopio de disección.
RESULTADOS
Inmersiones exploratorias
Encontramos corales juveniles producto de la reproducción sexual en diez de un total de 19 sitios inspeccionados en isla Gorgona entre 2010 y 2017. Se observaron corales juveniles en todos los arrecifes de coral y en algunos de los sitios rocosos. Los Farallones y El Arrecifito fueron los sitios con más especies (seis y cinco especies respectivamente, Figura 1), mientras que en los principales arrecifes de coral (La Azufrada, Playa Blanca, La Ventana, El Muelle) sólo se encontraron dos especies. Los corales juveniles pertenecían a por lo menos diez especies diferentes y cuatro familias (Tabla 1, Figura 2).
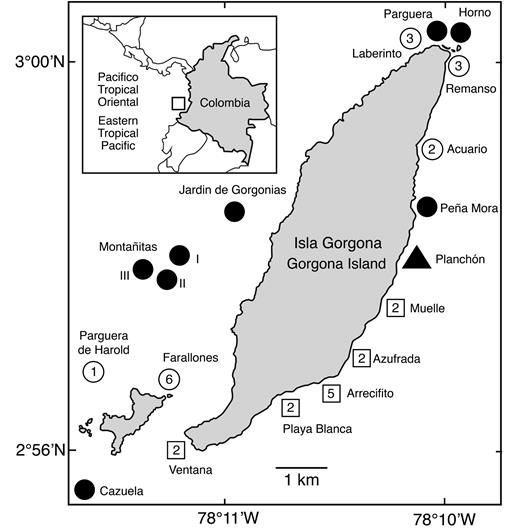
Figura 1 Número de especies de corales juveniles por sitio. Cuadrado = arrecife de coral; círculo = sitio rocoso; triángulo = naufragio en fondo arenoso. Negro = no se encontraron corales juveniles.
Tabla 1 Presencia de los corales juveniles observados en el Parque Nacional Natural Gorgona (Colombia, Pacífico Tropical Oriental) durante 2010 - 2017.
| Agariciidae | Pocilloporidae | Poritidae | Psammocoridae | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sitio Site | Ámbito de profundidad (m) Depth Range (m) | Cobertura de coral (~ %) Coral cover (~ %) | Años del estudio Study years | Gardineroseris planulata | Pavona chiriquiensis | Pavona clavus | Pavona frondifera | Pavona gigantea | Pavona varians | Pocillopora sp. | Porites panamensis | Porites lobata | Psammocora stellata |
| Arrecifes de coral Coral Reefs | |||||||||||||
| Arrecifito | 0-10 | 70.0% | 2010-2012 | - | - | - | - | X | X | X | X | - | X |
| Azufrada | 0-10 | 50.0% | 2010-2015 | - | - | - | - | - | X | X | - | - | - |
| Muelle | 0-10 | 70.0% | 2010-2015 | - | - | - | - | X | X | - | - | - | - |
| Playa Blanca | 0-10 | 70.0% | 2010-2015 | - | - | - | - | - | - | X | X | - | - |
| Ventana | 2-10 | 85.0% | 2010-2015 | X | - | - | - | - | - | X | - | - | - |
| Montañas rocosas/peñascos | |||||||||||||
| Rocky mounts / boulders | |||||||||||||
| Cazuela | 16-30 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Farallones | 0-12 | 5.0% | 2010 | - | X | X | X | - | - | X | - | X | X |
| Horno | 0-35 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Jardín de Gorgonias | 10-25 | 1.0% | 2017 | - | - | - | - | - | - | - | - | - | - |
| Parguera de Harold | 10-20 | 3.5% | 2017 | - | - | - | - | - | - | X | - | - | - |
| Montañita I | 7-25 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Montañita II | 3-25 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Montañita III | 20-40 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Peña Mora | 7-20 | 5.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Costas rocosas | |||||||||||||
| Rocky shores | |||||||||||||
| Acuario | 0-5 | 40.0% | 2010-2014 | - | - | - | - | X | - | X | - | - | - |
| Laberinto | 0-15 | 25.0% | 2010-2014 | X | - | - | - | - | - | X | - | X | - |
| Parguera | 0-12 | 35.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Remanso | 0-12 | 45.0% | 2010-2014 | - | - | - | - | X | - | X | - | X | - |
| Naufragio | |||||||||||||
| Shipwreck | |||||||||||||
| Planchón | 5-17 | 2.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |

Figura 2 Corales juveniles producto de reproducción sexual asentados sobre sustratos naturales en Isla Gorgona (Pacífico Tropical Oriental). Pocillopora sp. asentado sobre escombros de coral con algas calcáreas (A, B); Pocillopora sp. asentado sobre una pared rocosa con algas filamentosas (C); Porites cf. panamensis asentado sobre escombros de coral con algas calcáreas (D); Pavona varians asentado sobre escombros de coral con algas calcáreas (E); Pavona gigantea asentado sobre una pared rocosa con algas filamentosas (F). Las flechas blancas señalan la base incrustante en los ejemplares de pocilopóridos.
Conteo en transectos
Se observaron corales juveniles de al menos dos especies asentados sobre sustratos naturales en los transectos realizados en isla Gorgona. La especie Pocillopora sp. se encontró en los arrecifes de coral de La Azufrada y Playa Blanca, y en la costa rocosa de El Laberinto (Figuras 2A, 2B y 2C; Tabla 1), mientras que Porites panamensis se encontró en el arrecife coralino El Arrecifito (Figura 2D, Tabla 1).
Encontramos un total de 41 ejemplares de corales juveniles pocilopóridos, estimándose una densidad media de 0.07 ± 0.11 colonias m-2 (± DE). El mayor reclutamiento de corales se observó en La Azufrada (0.09 ± 0.12 colonias m-2), seguido de El Laberinto (0.08 ± 0.11 colonias m-2) y Playa Blanca (0.05 ± 0.08 colonias m-2); y no se encontraron diferencias significativas en la densidad de corales juveniles de Pocillopora sp. entre los tres sitios (F2, 26 = 0.28, p > 0.1). El tamaño medio de colonia de estos corales juveniles fue de 3.5 ± 1.0 cm, sin diferencias significativas entre los tres sitios (F2, 38 = 0.31; p > 0.1; Figura 3A, 3B, 3C). El juvenil pocilopórido de menor tamaño medía ~ 1.0 cm de diámetro y se observó en El Laberinto, asentado sobre una superficie rocosa vertical (similar a la Figura 2C).
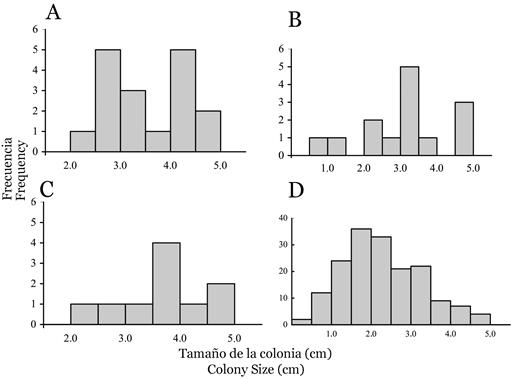
Figura 3 Distribuciones de frecuencia del tamaño de los corales juveniles (diámetro más largo de una colonia vista desde arriba) en Isla Gorgona (Pacífico Tropical Oriental). Las distribuciones se truncan a 5,0 cm, ya que los corales juveniles fueron definidos como colonias ≤ 5,0 cm. Distribución para los corales juveniles pocilopóridos (N = 41) en tres sitios: La Azufrada (A), El Laberinto (B) y Playa Blanca (C). Distribución para corales juveniles de Porites panamensis (N = 170) en El Arrecifito (D).
En El Arrecifito, se encontró un total de 170 corales juveniles de Porites panamensis, con una densidad media de 5.7 ± 4.7 colonias m-2 y un tamaño medio de colonia de 2.3 ± 0.9 cm. La colonia más pequeña de P. panamensis medía ~ 0.5 cm de diámetro, y tenía entre 15 y 20 pólipos con un diámetro cada uno ~ 0.1 cm (Figura 2D; 3D).
Placas de asentamiento
En contraste con los sustratos naturales, no se encontraron corales juveniles o reclutas asentados sobre las placas de asentamiento durante las visitas al sitio de estudio, ni al completar el análisis de las fotografías, ni tampoco después de un minucioso examen directo de las placas con un microscopio de disección.
DISCUSIÓN
Este estudio registra por primera vez en el Pacífico colombiano, evidencia del reclutamiento de corales provenientes de reproducción sexual. Sin embargo, se obtuvieron dos resultados contradictorios similares a los descritos en estudios previos sobre el reclutamiento de corales en el PTO: a pesar de las bajas densidades, se encontraron corales juveniles en los sustratos naturales de arrecifes de coral y sitios rocosos, pero el método de muestreo habitual de las placas de asentamiento no detectó reclutamiento coralino (Birkeland, 1977; Wellington, 1982; Richmond, 1985; Medina-Rosas et al., 2005; López-Pérez et al., 2007; Lozano- Cortés y Zapata, 2014). En los arrecifes de coral, los corales juveniles se encontraron asentados sobre los escombros de coral o en la matriz coralina consolidada cubierta por algas calcáreas, mientras que, en los sitios rocosos, los corales juveniles se encontraron asentados sobre paredes rocosas, desnudas o parcialmente cubiertas por algas filamentosas.
Además de constituir nuevos sitios del PTO en los que han sido observadas colonias juveniles de Pocillopora sp. y Porites panamensis producto de la reproducción sexual sobre sustratos naturales (Richmond, 1985; Medina-Rosas et al., 2005; Glynn et al., 1991, 1994, 2017a), adicionalmente, este estudio describe la distribución de la abundancia y del tamaño de los juveniles para estas especies en el Pacífico colombiano. Los valores del reclutamiento de corales sobre sustratos naturales en los arrecifes del PTO han sido muy bajos, indicando que la abundancia aquí registrada de juveniles de P. panamensis es una de las cifras más altas conocidas hasta el momento en la región, siendo superado solo por la abundancia de juveniles de Pavona clavus en Costa Rica (Glynn et al., 2017a) y seguramente resultado de un pulso de reclutamiento de un modo reproductivo incubador (Glynn et al., 1994).
La implicación de los resultados del reclutamiento de corales obtenidos hasta ahora en el PTO es doble. Primero, los corales sí pueden reproducirse sexualmente en la región, lo cual ya había sido previamente discutido por varios estudios histológicos sobre la reproducción del coral a lo largo del PTO (Glynn et al., 2017a y referencias incluidas). Estos resultados rechazan la hipótesis de que los corales sean estériles en la región (Richmond, 1985, 1987; Glynn et al., 1991), y al mismo tiempo rebaten la idea de que el reclutamiento derive principalmente de la fragmentación de las colonias (Birkeland, 1977; Highsmith, 1982; Wellington, 1982; Richmond, 1985).
En segundo lugar, la escasez del reclutamiento de corales en sustratos artificiales en el PTO puede ser debido a algún factor desconocido que impide que las larvas de coral se asienten o sobrevivan sobre los sustratos artificiales muestreados. Los resultados negativos obtenidos con las placas de asentamiento podrían estar relacionados con: 1) el uso de materiales de sustrato inapropiados, 2) tiempo insuficiente después de la instalación para un adecuado acondicionamiento del sustrato (López-Pérez et al., 2007), y 3) competencia, depredación o herbivoría en las placas de asentamiento (Birkeland, 1977; Sammarco, 1982, 1985; Richmond, 1987). Sin embargo, en este estudio, la primera causa es una explicación poco probable porque se utilizaron cinco materiales diferentes, incluyendo la cerámica, la cual ha producido los mejores resultados de reclutamiento tanto en el Indo-Pacífico (Harriott y Fisk, 1987) como en el Caribe (Tomascik, 1991). En el segundo caso, es bien sabido que los sustratos artificiales requieren un período de acondicionamiento durante el cual se desarrolla una comunidad incrustante antes de que ocurra el asentamiento de las larvas de coral (Segal et al., 2012). Aunque la duración del período de acondicionamiento no está bien establecida y puede variar mucho de un lugar a otro, en el sudoeste de México el reclutamiento de corales tuvo lugar de 5 a 10 meses después de sumergir las placas (López-Pérez et al., 2007). Dado que nuestras placas se mantuvieron sumergidas durante poco más de seis meses, es razonable pensar que la comunidad incrustante que induce o mejora el asentamiento de los corales aún no se hubiera desarrollado completamente. Sin embargo, en un intento previo de documentar el reclutamiento en placas de asentamiento en el mismo arrecife (La Azufrada), las placas fueron sumergidas durante un año completo y aun así no se encontraron reclutas (Lozano-Cortés y Zapata, 2014). En el tercer caso, se sabe que la competencia, la depredación, o la herbivoría en las placas de asentamiento, pueden evitar que los reclutas de coral sobrevivan y se produzca un verdadero reclutamiento sobre estos sustratos (Birkeland, 1977; Sammarco, 1982, 1985; Díaz-Pulido et al., 2010). Aunque se eliminaron periódicamente algas y otros invertebrados sésiles de la mitad de las placas para reducir la posible competencia con los potenciales reclutas coralinos, con frecuencia se observaron erizos de mar, ophiuroides, y gasterópodos sobre las placas, que pudieron haber interferido con el asentamiento o supervivencia del coral.
Además, existe un argumento metodológico razonable relacionado con el tamaño de la muestra, cuya solución puede ser imposible en la práctica: si la abundancia del reclutamiento de corales es naturalmente baja, entonces el área o el esfuerzo de muestreo debería ser considerablemente mayor que la utilizada para poder detectar con éxito los pocos reclutas de coral. Si bien existen otras diversas posibles explicaciones, ninguna de ellas es lo suficientemente general como para explicar el fracaso común para documentar el reclutamiento de corales en placas de asentamiento en el PTO, y, por lo tanto, la razón para estos resultados sigue sin estar clara.
En conclusión, los resultados presentados en este estudio indican que existe reclutamiento de corales producto de la reproducción sexual en Isla Gorgona. Esto, sin embargo, ha sido pasado por alto por la falta de observaciones de asentamiento coralino en sustratos artificiales, y la dificultad para diferenciar a los corales juveniles producto de la reproducción sexual de aquellos que son producto de la sobrevivencia de fragmentos de coral. Las futuras investigaciones requieren examinar la contribución relativa de la reproducción sexual y asexual al reclutamiento coralino, y su papel en la recuperación y el mantenimiento de las poblaciones coralinas y los arrecifes en el Pacífico colombiano y el PTO.

