










Services on Demand
Journal
Article
 text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.48 no.1 Santa Marta Jan./June 2019 Epub Aug 30, 2019
https://doi.org/10.25268/bimc.invemar.2019.48.1.755
Research Articles
Growth parameters of Cardisoma guanhumi in the department of Magdalena, Colombia

1 Universidad Nacional de Colombia, Sede Caribe, CECIMAR, El Rodadero, Santa Marta, Colombia. lfcardonaac@unal.edu.co, nhcamposc@unal.edu.co.
2 Universidad Nacional de Colombia, Sede Caribe, CECIMAR, El Rodadero, Santa Marta, Colombia. nhcamposc@unal.edu.co.
3 Parques Nacionales Naturales de Colombia, Bogotá, Colombia. elkinhernandezrolon@gmail.com.
In Colombia, the blue land crab Cardisoma guanhumi faces two main threats: extraction and habitat loss. This project aims to show an approximation of the growth parameters of C. guanhumi in protected areas within the Magdalena department. Individuals were captured monthly from May to October 2016 in five localities in the Tayrona National Natural Park (PNNT) and the Salamanca Island Road Park (VIPIS). A total of 816 crabs were collected, each of which was measured, weighed, sexed and released. Both sexes showed a negative allometric length-to-weight ratio. Using the FiSAT II program's ELEFAN I routine, the size frequencies obtained were analyzed in order to determine the growth parameters for 1) the department, considering gender, and 2) the PNNT, where the data from VIPIS was excluded. The asymptotic carapace width was higher for males (109.16 mm) than for females (96.74 mm), but the growth coefficient (K) was higher for females (0.19 year-1) than for males (0.15 year-1). Growth performance indexes were similar: 3.252 for males and 3.250 for females. The FISAT II length converted catch curve routine was used to calculate the total (Z), natural (M), and fishing (F) mortality rates, as 0.41, 0.36, and 0.05 year-1 respectively. The grow coefficient K was higher for the PNNT (0.29 year-1) than for the Magdalena department (0.15 year-1), thus, longevity was lower for the PNNT (tmax = 10.42 years) compared to the department (tmax = 20.06 years). In general terms, the C. guanhumi population in the study area is longevous, slow growing, with low extraction morality. The data obtained in this work are important in the management of this species, as its slow growth makes it vulnerable to overfishing.
KEYWORDS: Cardisoma guanhumi; von Bertalanffy; Growth parameters; FISAT; Magdalena Colombia
En Colombia, el cangrejo Cardisoma guanhumi enfrenta dos amenazas: la extracción y la pérdida de hábitat. El presente trabajo tuvo como objetivo estimar los parámetros de crecimiento de C. guanhumi en el departamento del Magdalena. Se realizaron capturas mensuales de mayo a octubre de 2016 en cuatro localidades del Parque Nacional Natural Tayrona (PNNT) y una localidad de la Vía Parque Isla Salamanca (VIPIS). Se capturaron 816 individuos, los cuales fueron medidos, pesados, sexados y liberados. Con la rutina ELEFAN I del programa FiSAT II se analizaron las frecuencias de tallas obtenidas para conocer: 1) los parámetros de crecimiento a nivel departamental, teniendo en cuenta el sexo, para lo cual se usaron todos los datos y 2) para el PNNT, para lo cual se excluyeron los datos de VIPIS. El tamaño asintótico del ancho del caparazón fue mayor en machos (109.16 mm) que en hembras (96.74 mm), mientras que el parámetro de crecimiento K fue mayor para las hembras (0.19 año-1) que para los machos (0.15 año-1). Se obtuvieron índices de desempeño de crecimiento similares: 3.252 en machos y 3.250 en hembras. Con la rutina de curva de captura linealizada de FISAT II se calcularon las tasas de mortalidad total (Z), natural (M), y por pesca (F), las cuales fueron 0.41, 0.36 y 0.05 año-1 respectivamente. Para el PNNT se obtuvo un K mayor (0.29 año-1) que en el departamento (0.15 año-1) y por tanto una menor longevidad (tmax = 10.42 años) que en el departamento (tmax = 20.06 años). En términos generales la población de C. guanhumi es longeva, de crecimiento lento y con baja mortalidad por extracción en la zona de estudio. Los datos obtenidos en el presente trabajo son importantes para el manejo de esta especie, ya que al tener un crecimiento lento la hace vulnerable a la sobrepesca.
PALABRAS CLAVE: Cardisoma guanhumi; von Bertalanffy; Parámetros de crecimiento; FISAT; Magdalena Colombia
INTRODUCTION
The blue land crab Cardisoma guanhumi Latreille in Latreille, Le Peletier, Serville and Guérin, 1828, is a widely distributed species spanning from southeastern Florida, United States, to Santa Catarina, Brazil, including Bermuda and the West Indies in the Caribbean Sea (Taissoun, 1974; Branco, 1990). In Colombia, it is distributed in all the departments located on the Colombian continental Caribbean and on the island of San Andrés.
C. guanhumi populations currently face two main threats. Given that the species constitutes an important economic resource in places such as Venezuela (Taissoun, 1974; Carmona-Suárez, 2011), Brazil (Oshiro et al., 1999; Botelho et al., 2001), Mexico (Rivera, 2005), Cuba (Álvarez et al., 1993) and other Caribbean islands, in which it has even been overexploited, one of these threats is extraction. The other is the disruption or loss of their habitat (Rodríguez-Fourquet and Sabat, 2009). In Colombia, the species is classified as Vulnerable (VU) due to the pressures it faces through habitat deterioration and indiscriminate capture (Bermúdez et al., 2002).
Knowing the growth parameters and mortality rates of the populations of a species is of great importance to generating adequate management and conservation strategies, as these allow us to evaluate how extraction or other types of anthropogenic activities can affect the populations. Unfortunately there are few studies on these aspects for C. guanhumi populations. Under laboratory conditions, Henning (1975) estimated that an individual with 98 mm carapace width can be 13 years old and only two studies, both conducted in Brazil, have been carried out to determine the population growth parameters of this species. The first was carried out in an area where this crab is exploited, where a K= 0.594 years-1 was estimated (Botelho et al., 2001). The second study was carried out in an area where commercial catches do not take place, and the estimated K was 0.145 years-1 (Costa, 2016), which, in both cases, provides evidence of the species'' slow growth rates. Bearing this in mind, the purpose of this study is to estimate the growth parameters of C. guanhumi populations in protected areas in the department of Magdalena, in the Colombian Caribbean, based on the size frequencies in order to obtain relevant information to help manage the species.
MATERIALS AND METHODS
Study areas
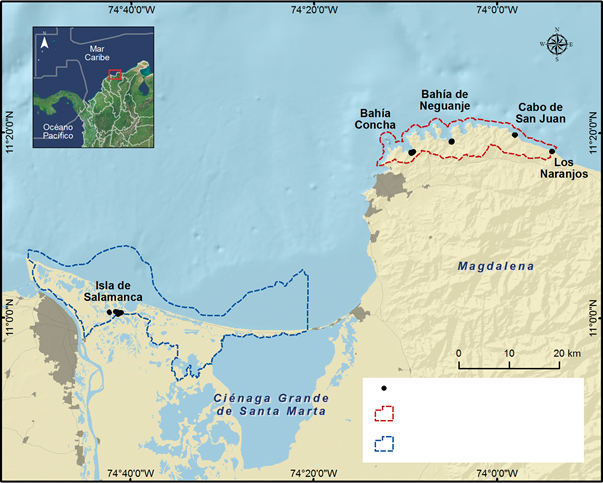
Five locations in the Colombian Caribbean belonging to two natural parks in the department of Magdalena were chosen: The Centro de Visitantes in the Salamanca Island Road Park (SIRP), and Bahía Concha, Bahía Nenguange, Cabo San Juan del Guía and Los Naranjos in the Tayrona National Natural Park (TNNP) (Figure 1). Despite being protected areas, these places are tourist attractions in which environmental modifications have been made, related to housing, transport or tourism, and where, despite the efforts of the relevant entities, illegal extraction of this crab may take place. However, there is no information on the intensity with which this occurs.
Population structure
Individuals were collected monthly, from May to October 2016, in each of the localities. To capture the crabs, kitchen tongs and landing nets were used. The latter were thrown on the individuals in order to trap them. This was carried out at night, given that in the exploratory outings, it became apparent that the crabs' activity increased at night. Traps were also made with plastic bottles with an inlet diameter of 7.62 cm and 10.16 cm, packed with raw chicken or mango as bait, and left overnight. These were checked the following day. For each individual captured, the following biometric measurements were taken using calipers and scales: carapace width (AC) and length (LC), and weight. Each crab was sexed taking into account the shape of its abdomen: triangular or inverted V in males, and rounded in females. For the latter, note was taken on whether they were ovigerous or not.
Size frequency histograms for both males and females were derived from the structural data. A Chi-square test with a 95% confidence level (Sokal and Rohlf, 1979) was performed to find out whether the ratio of males to females differed significantly from 1:1.
Growth parameters
The width-weight relationship for males and females was estimated by regression analysis using potential equation:
P = a*ACb (Ricker, 1975)
Where P corresponds to the total weight (g), AC is the carapace width (mm), parameter a is the intercept and b is the curve gradient. These data were transformed with natural logarithm and the null hypothesis (H 0 : b = 3) was tested with the student t test (α = 0.05). Parameter b represents the allometry coefficient, so that when b = 3, the growth is isometric, and when b ≠ 3, the growth is allometric. Because the AC was used in millimeters and not centimeters, the intercept was corrected using a= a'*10b (Froese, 2006). For this analysis, we excluded those individuals who were missing one of their limbs.
The von Bertalanffy growth equation (1938) was used to describe the crabs' growth parameters:
AC t = AC∞ (1-e-(K(t-t0)),
Where ACt is the carapace width at age t, AC∞ is (Pauly, 1984), where ACmax is the maximum carapace width recorded during the sampling. Subsequently, to estimate the curvature parameter K, the monthly size (carapace width) frequency distribution of males and females separately was used, by grouping them into 2 mm classes. These data, together with the calculated AC∞, were analyzed using the subroutine K scan, within the ELEFAN I routine (Electronic Length Frequency Analysis) (Pauly, 1987; Pauly and David, 1981), incorporated in the FiSAT II program (FAO-ICLARM Stock Assessment Tools II) (Gayanilo et al, 2005). Subsequently t0 was calculated using the empirical equation of (Pauly, 1979):
The ELEFAN routine has been widely used because it is a low-cost method that depends only on regular measurements of individual sizes, and because it does not require knowledge of age, which can be particularly difficult in crustaceans because permanent growth structures are scarce in this group.
Subsequently, the ages corresponding to the different sizes were calculated using the inverted von Bertalanffy equation:
t = t0 - (1/k) * ln (1-ACt / AC∞ )
Finally, longevity was estimated using the following equation:
t max = t0+ 3/k (Taylor, 1958).
An analysis of growth parameters at departmental level was conducted, using data from all sampled localities. Two estimates were made: one total and one sex-sensitive. Aditionally, the parameters for each one of the parks were estimated in order to observe if there were differences among them. This way, the parameters for the PNNT were determined, for which data from the VIPIS location were excluded. Unfortunately the analysis for VIPIS was not carried out due to the sample size of this location. So a comparison was made only between the parameters at departmental level and those of the PNNT.
Subsequently, the growth performance index phi prime (Ø’) was estimated, which tends to be similar in stocks of the same species and even in species of the same family (Munro and Pauly, 1983). This was calculated using the growth parameters K and AC∞:
Ø’ = logK+2logAC ∞. (Pauly y Munro, 1984)
We also calculated (Ø’) for other populations of C. guanhumi, using data available in publications and compared with the estimate for the department of Magdalena. Size frequency data were also used to calculate the total mortality rate (Z), using a length converted catch curve routine, in FISAT, based on the equation:
Ln(Ni/Δti) = a+b*ti (Pauly, 1983)
Where Ni is the number of individuals captured in a class i interval, Δti is the time required by the crab to grow across the length of class i, ti is the age corresponding to the class i interval; parameters a and b are calculated by means of a regression analysis, in which slope b, with the sign changed, corresponds to the Z estimated. The program considered those individuals viable for capture, that is, those from the point at which the capture curve begins to fall at a constant rate (Pauly, 1984); and therefore the regression to calculate Z was carried out using this group of individuals. The calculation of the natural mortality rate (M) was based on the Pauly equation (1980): , where T is the annual mean temperature of the place where the individuals were collected (28°C); and the fishing mortality (F) was calculated using the formula F=Z-M. The mortality rate was estimated for the general population, given that C. guanhumi extraction is not sex-selective
RESULTS
A total of 816 individuals were collected during the six-month sampling period. Of these, 181 in Bahía Concha, 154 in Cabo San Juan del the Salamanca Island Road Park, 186 in Nenguange, 183 in Los Naranjos and 112 in the Salamanca Island Road Park (SIRP). Except for the VIPIS location, in all other locations, the individuals were most active during the night, making SIRP the smallest sample size.
A Spearman correlation (Sokal and Rohlf, 1979) of the carapace width-length ratio was calculated, in which it was evidenced that the measurements are strongly associated (correlation coefficient: 0.99, p < 0.001), thus, only the carapace width (AC) was used for the other analyses.
The size frequency distribution (Figure 2) shows that, in general, the males present a wider distribution, between 20.2 and 103.7 mm (61.35 ± 0.87 mm) (average ± standard error), while in females it was between 25.9 and 91.9 mm (57.66 ± 0.50 mm). In females, a modal distribution of 62-64 mm was observed, while in males it was not possible to observe a clear modal distribution. Male weights were between 4 and 516 g (133.49 ± 5.21 g), and female weights were between 9 and 266 g (95.90 ± 2.12 g). Sexual ratio differed significantly from 1:1 (x2= 20.08, p < 0.001), with a predominance of males.
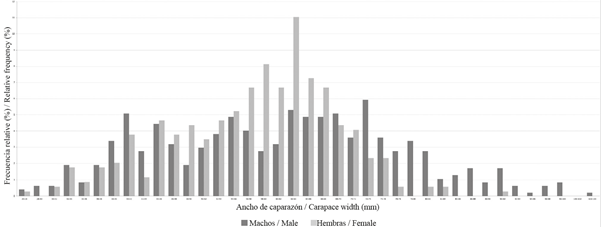
Figure 2 Histogram of Cardisoma guanhumi size frequency for males and females collected between May and October 2016 in protected areas within the department of Magdalena.
The width-weight relationship for both males and females showed negative allometric growth (b = 2.93, IC = 2.89 - 2.97, t-student = 3.186, p = 0.002 for males and b = 2.80, IC = 2.74 - 2.86, t-student = 6.397, p < 0.001 for females) (Figure 3).
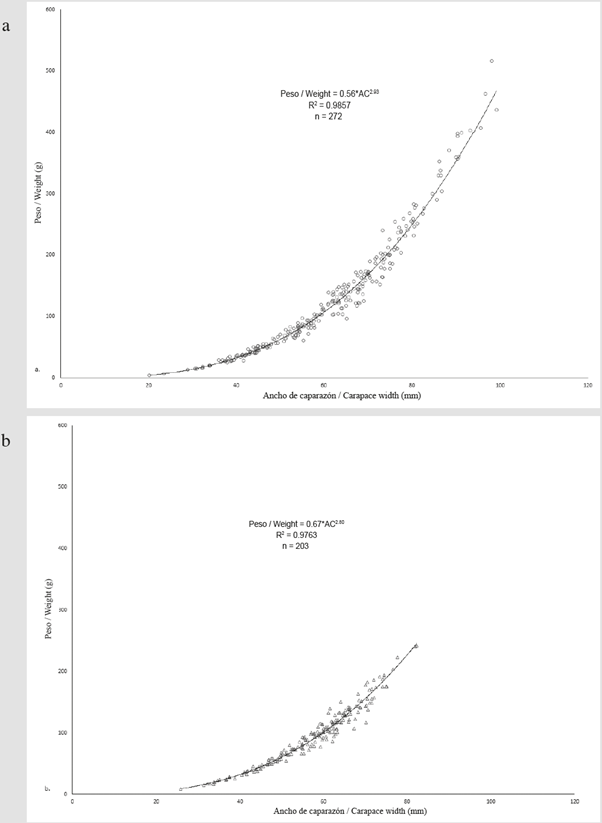
Figure 3 Carapace width-weight relationship for C. guanhumi males (a) and females (b) collected between May and October 2016 in protected areas within the department of Magdalena.
For the growth parameters of C. guanhumi in the department of Magdalena, a higher ACmax was obtained in males than in females, making the estimated AC∞ higher for males. When entering the respective values of AC°° in the ELEFAN K scan subroutine, the males had a lower K value than the females. This indicates that males are slower growing and have a longer lifespan than females (Table 1 and Figure 4). With regard to the parameters estimated for the PNNT, a higher K value and a lower longevity were found than those obtained for the department (taking into account both PNNT and VIPIS individuals) (Table 2).
Table 1 Growth parameters for Cardisoma guanhumi males and females in the department of Magdalena. n= number of individuals sampled, ACmax = maximum carapace width obtained in the study, AC∞=asymptotic carapace width, K= curvature parameter, t0= the theoretical age at which the individual is size 0, tmax= maximum age, Ø′=growth performance index.
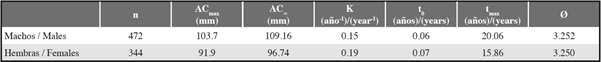
Table 2 Growth parameters for Cardisoma guanhumi males and females in the department of Magdalena. n= number of individuals sampled, ACmax = maximum carapace width obtained in the study, AC∞ = asymptotic carapace width, K= curvature parameter, t0= the theoretical age at which the individual is size 0, t = maximum age, Ø’=growth performance index.
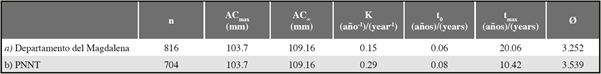
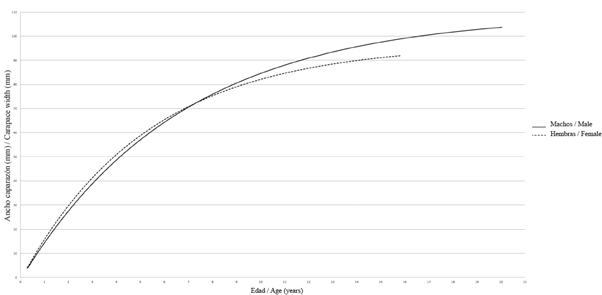
Figure 4 von Bertalanffy growth curve for Cardisoma guanhumi males and females captured in different localities in the Magdalena department. The growth curves were plotted up to the ACmax age obtained in the study.
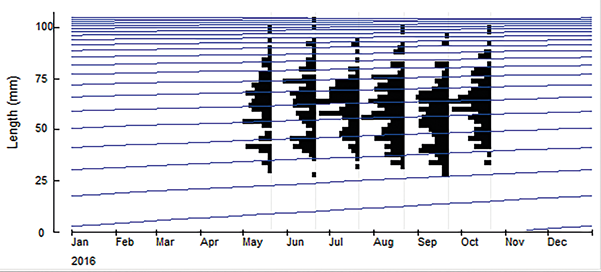
Using this same routine, the monthly frequency distributions of carapace width for the Magdalena department population were plotted and it was found that there is no modal progression, as it is not possible to visualize cohorts (Figure 5). The growth performance index (Ø’) was similar in males and females. The estimated total mortality rate for the department's population was Z=0.41 year-1, the natural mortality rate was M=0.36 year-1 and the fishing mortality rate was F=0.05 year-1.
DISCUSSION
The distribution of the monthly frequencies was multimodal, since it was not possible to observe the presence of cohorts throughout the months. This has been previously observed for this species and has been attributed to the fact that the recruitment of juveniles is continuous, and to the species' characteristic slow growth (Silva et al., 2014; Costa, 2016).
It was possible to show that the males reached larger sizes, grew more slowly and had greater longevity than the females. Costa (2016), using a mark-recapture method, obtained K = 0.159 year-1 for males, which is very similar to that obtained in this study (K = 0.150 year-1). The author found, however, that females grow slower than males (Table 3). This may be related to the AC∞ estimated in that study, which was higher for females than for males (Table 3), which is unusual since it has been widely reported that Gecarcinidae family males reach larger sizes and higher weights than females, as the latter invest more energy in reproductive processes (Hartnoll, 1982, 1988).
Table 3 Comparison of the estimated growth parameters for Cardisoma guanhumi with respect to the literature. M= males, F= females, T= total. AC∞= asymptotic carapace width, K= growth coefficient, t = theoretical lifetime, phi prima index. *Data calculated from the information reported by the authors. To estimate tmax we used: tmax= t0 + (3/K) (Taylor, 1958), if t0 was not reported, 0 was used in the equation. For phi prima the following was used: Pauly y Munro (1984). **Carapace length measurements (LC). For Botelho et al. (2001) tmax was not estimated because the authors calculated K from LC values. *** Data not reported by the author.
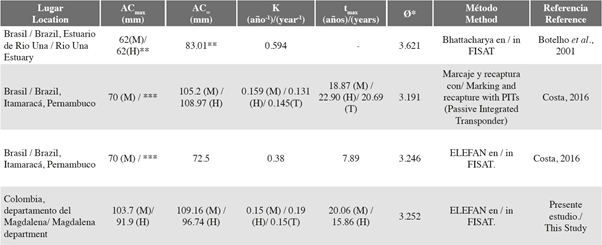
As previously mentioned, there are three approaches to growing this species: one in laboratory conditions (Henning, 1975), and the other two in natural environments. In the first of these, Botelho et al. (2001) used the Bhattacharya method (which is also based on size frequencies), and in the second, Costa (2016) compared two methodologies: ELEFAN and mark-recapture (Table 3). In both studies, the K estimates were higher than those in this paper, possibly because of the maximum reported sizes and the length measurements used, which are important factors in the estimation of growth parameters in the size frequency analyses. For example, Botelho et al. (2001), whose samples were taken in a locality in which C. guanhumi was exploited, estimated the growth parameters based on LC measurements and not AC ones. They reported an LCmax=62 mm, which could lead to a high K being calculated.
70 mm, estimated two values for K, the first result from an ELEFAN analysis (n=1078) and the second using marking and recapture data (n=291). Thus, Costa (2016) obtained a higher K with ELEFAN than the K estimated from tagging, catch and recapture data. This can be explained because to estimate the K parameter, the ELEFAN routine is based on AC∞ (which is calculated from the ACmax acquired in the sample). It may be that during sampling, Costa (2016) was not able to capture an individual with an ACmax representative of the population (it is likely that there were larger sizes considering that the sampling was carried out in a surveillance area where no commercial catches of C. guanhumi occur). Thus, when calculating a smaller AC∞ the value of K is overestimated.
While with the tagging marking and recapture data, Costa (2016) was able to estimate the increase in length from direct changes in crab growth, and therefore the K parameter was not affected by the ACmax obtained in the sampling.
Based on the growth parameters for the PNNT, it was observed that. C. guanhumi showed faster growth (K= 0.29 year-1) and shorter longevity (tmax = 10.42 years) than at departmental level (K= 0.15 year-1 and tmax = 20.06 years). Even though both PNNT and VIPIS are part of the Magdalena department, it is possible that the differences obtained when analyzing the PNNT data separately are related to the proximity between the four localities of the PNNT, since, by being closer, these populations are more closely related and therefore have more common characteristics, such as growth. Additionally, it was evident that C. guanhumi individuals from VIPIS showed a different behavior from other localities, as during most months, the individuals showed little activity outside the burrows, restricting the sample size in this locality.
The growth performance indices (Ø’) for the department were similar to those calculated for the Costa tagging and recapture data (2016): 3,246 and 3,191 for males and females respectively (Table 3). On the other hand, the Ø’ calculated for Botelho et al. data (2001) was 3,621, slightly higher than the others (Table 3). However, as previously mentioned, these differences may be related to the fact that the study used the LC and not AC for size frequency analysis.
The total mortality rate (Z = 0.41 year-1), natural mortality (M = 0.36 year-1) and fishing mortality (F = 0.05 year-1) in the department were low, which is consistent with the fact that the surveys were conducted in protected areas. It should be noted that for Bahía Concha there have been reported events of extraction of C. guanhumi, so a study focused on this place is required to evaluate whether the mortality caused by extraction is high here. Costa (2016) estimated a total mortality Z = 2.31 year-1, based on monthly catch data and values of AC∞ and K obtained with tagging and recapture data (108.03 mm and 0.145 year-1). This high mortality in Costa (2016) could be related to the difference between AC∞ (108.03 mm) and ACmax (70 mm), which would be interpreted as high mortality in adult individuals. This author did not determine fishing mortality, as she worked in an area where they are not caught for commercial purposes and therefore, assumed Z=M.
From the growth parameters estimated in the present study, and those obtained by Costa (2016) considering that the marking and recapture data provide more accurate information on growth, it is possible to infer that C. guanhumi is a slow growing and longevous species, with a life expectancy of more than 10 years. These characteristics have also been reported in other large terrestrial crustaceans such as Ucides cordatus, which has been estimated to live up to 17 years (Diele and Koch, 2010).
It should also be noted that differences in growth parameters might not be related only to the methodology used, but also to other factors that may influence growth rate and longevity. These include food availability, habitat quality, and extraction, among others, which may generate differences between populations of the same species. This may explain the differences obtained in the value of K obtained at departmental level with respect to the K obtained for the PNNT. It is also possible that if the localities within the PNNT were evaluated separately, differences in crab growth could be observed between these.
On the other hand, the implementation of capture and recapture methods should be considered in order to obtain more robust data in growth analyses in species such as C. guanhumi characterized by slow growth, longevity and constant recruitment of juveniles (Silva et al, 2014; Costa, 2016), which may present difficulties when using methods based on size frequencies.
CONCLUSIONS
In general, the C. guanhumi population studied showed negative allometric growth, is longevous, and slow growing, which coincides with what has been previously reported for this species. This is of great importance because when a slow-growing species is subjected to intensive extraction, it requires more time to recover its stocks (Silva et al, 2014). This should be taken into account when devising management plans for the responsible use of this crab. In addition, low fishing mortality (extraction) was obtained, indicating that, from a general perspective for the sampled area, the species is not being overexploited.
ACKNOWLEDGEMENTS
The authors would like to thank Universidad Nacional de Colombia, Caribbean Campus, for its funding through CECIMAR. The National Parks administrative unit, for authorizing the study, and for providing the available personnel and infrastructure. Likewise we would like to thank the officials at Dirección Territorial Caribe for all the support in coordinating field trips. We would like to thank the evaluators and editors for their suggestions and improvements to the manuscript. Contribution No. 479 from Instituto de Estudios en Ciencias del Mar, CECIMAR, at the Caribbean Campus of Universidad Nacional de Colombia.
REFERENCES
Álvarez, I., E. Gimenez, P. Machado y R. Flores. 1993. Explotación y aprovechamiento del cangrejo de tierra (Cardisoma guanhumi ) en la ciénaga de Zapata, Cuba. Rev. Sian Ka'an, Ser. Doc., 1: 48-56. [ Links ]
Bermúdez, A., N.H. Campos y G.R Navas. Cardisoma guanhumi. 120-122. En: Ardila, N., G.R. Navas y J. Reyes (Eds.). 2002. Libro rojo de los invertebrados marinos de Colombia. Invemar, Min. Medio Amb. La serie Libros rojos de especies amenazadas de Colombia, Bogotá. 175 p. [ Links ]
Bertalanffy, L.V. 1938. A quantitative theory of organic growth. Human Biol., 10(2): 181-213. [ Links ]
Botelho, E.R., M. do C. do Santos y J.R. Souza. 2001. Aspectos populacionais do Guaiamum, Cardisoma guanhumi Latreille 1825, do estuário do Rio Uma (Pernambuco-Brasil), Bol. Técn. Cient. CEPENE. Tamandaré, 9(1): 123-146. [ Links ]
Branco, J. 1990. Aspectos ecológicos dos brachyura (Crustácea: Decapoda) no manguezal do Itacorubi, SC-Brasil., Rev. Bras. Zool., 7(1-2): 165-179. [ Links ]
Carmona-Suárez, C. 2011. Present status of Cardisoma guanhumi Latreille, 1828 (Crustacea: Brachyura: Gecarcinidae) populations in Venezuela. Interciencia, 36(12): 908-913. [ Links ]
Costa, D.F. de M. 2016. Aspectos de crescimento e mortalidade do guanhumi (Cardisoma guanhumi) em um manguezal de acesso restrito na Ilha de Itamaracá-PE. 70p. [ Links ]
Diele, K. and V. Koch. 2010. Growth and mortality of the exploited mangrove crab Ucides cordatus (Ucididae) in N-Brazil, J. Exp. Mar. Biol. Ecol., 395(1-2): 171-180. [ Links ]
Froese, R. 2006. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations. J. App. Icht., 22(4): 241-253. [ Links ]
Gayanilo, F.C.J., P. Sparre and D. Pauly. 2005. The FAO-ICLARM stock assessment tools II (FISAT II). Revised version. User's guide, FAO Computerized Information Series - Fisheries, No. 8, FAO, Rome. 168 p. [ Links ]
Hartnoll, R. 1982. Growth, The biology of Crustacea: embryology, morphology and genetics. Academic Press, New York. 196 p. [ Links ]
Hartnoll, R. 1988. Growth and moulting, Biology of the land crabs. Cambridge, Cambridge Univ. Press, 186-210 p. [ Links ]
Henning, H. 1975. Kampf-, fortplanzungs- und hautungsverhalten-wachstum und geschlechtsreife von Cardisoma guanhumi Latreille (Crustacea, Brachyura). Forma et Functio, 8: 463-510. [ Links ]
Munro, J. and D. Pauly . 1983. A simple method for comparing the growth of fishes and invertebrates. Fishbyte, 1(1): 5-6. [ Links ]
Oshiro, L.M.Y., R.. Silva y C.. Silveira. 1999. Rendimento de carne nos caranguejos guaiá, Menippe nodifrons Stimpson, 1859 e guaiamum, Cardisoma guanhumi Latreille, 1825. Acta Biol. Leopoldensia, 21(1): 83-88. [ Links ]
Pauly, D. 1979. Gill size and temperature as governing factors in fish growth: a generalization of von Bertalanffy's growth formula. Berich. Inst. Meeresk. Univ. Kiel. 156 p. [ Links ]
Pauly, D. 1980. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci., 39(2): 175-192. [ Links ]
Pauly, D. 1983. Lengt-converted catch curves. A powerful tool for fisheries research in the tropics. Part I. Fishbyte, 1: 9-13. [ Links ]
Pauly, D. 1984. Fish population dynamics in tropical waters: a manual for use with programmable calculators, ICLARM Stud. Rev. 8, 325 p. [ Links ]
Pauly, D. 1987. A review of the ELEFAN system for the analysis of length-frequency data in fish and aquatic invertebrates: 7-34. In Pauly, D. and G Morgan (Eds.) Length-based methods in fisheries research. ICLARM and Kuwait Institute for Scientific Research. [ Links ]
Pauly, D. and N. David. 1981. ELEFAN I, a basic program for the objective extraction of growth parameters from the lengh-frecuency data. Meeresforsch. Rep. Mar. Res., 28(4): 205-211. [ Links ]
Pauly, D . and J. L. Munro. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte, The WorldFish Center, 2(1): 1-21. [ Links ]
Ricker, W. 1975. Computation and interpretation of biological statistic of fish populations. Bull. Fish. Res. Board Canada, 191(1): 382. [ Links ]
Rivera, J.J. 2005. El cangrejo terrestre Cardisoma guanhumi ¿un recurso pesquero?, ECOfronteras, 25: 15-17. [ Links ]
Rodríguez-Fourquet, C. and A.M. Sabat. 2009. Effect of harvesting, vegetation structure and composition on the abundance and demography of the land crab Cardisoma guanhumi in Puerto Rico. Wetl. Ecol. Manag., 17(6): 627-640. [ Links ]
Silva, C., R. Schwamborn and J.L. Oliveira. 2014. Population biology and color patterns of the blue land crab, Cardisoma guanhumi Latreille 1828) (Crustacea: Gecarcinidae) in the Northeastern Brazil, Braz. J. Biol., 74(4): 949-958. [ Links ]
Sokal, R. y F. Rohlf. 1979. Biometría: principios y métodos estadísticos en la investigación biológica. Blume, Madrid. 832 p. [ Links ]
Taissoun, E. 1974. El cangrejo de tierra Cardisoma guanhumi (Latreille) en Venezuela. Bol. Centro Invest. Biol., 10: 1-36. [ Links ]
Taylor, C.C. 1958. Cod growth and temperature. ICES J. Mar. Sci ., 23: 366-370. [ Links ]
Received: May 25, 2018; Accepted: March 13, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
