











 text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El cangrejo azul de tierra Cardisoma guanhumi Latreille in Latreille, Le Peletier, Serville y Guérin, 1828 es una especie con una distribución amplia que va desde el sureste de Florida, Estados Unidos, hasta Santa Catarina, Brasil, incluyendo Bermudas y las Antillas en el Mar Caribe (Taissoun, 1974; Branco, 1990). En Colombia se encuentra distribuida en todos los departamentos ubicados en el Caribe continental colombiano y en la isla de San Andrés.
Actualmente, las poblaciones de C. guanhumi enfrentan dos amenazas principalmente. Una de estas es la extracción, ya que esta especie constituye un recurso económico importante en algunos lugares como Venezuela (Taissoun, 1974; Carmona-Suárez, 2011), Brasil (Oshiro et al., 1999; Botelho et al., 2001), Mexico (Rivera, 2005), Cuba (Álvarez et al., 1993) y otras islas del Caribe, e incluso se ha llegado a sobreexplotar en algunos de estos sitios. Otra amenaza que enfrentan las poblaciones de C. guanhumi es la alteración y/o pérdida del hábitat (Rodríguez-Fourquet y Sabat, 2009). En Colombia se encuentra clasificada como Vulnerable (VU) debido a las presiones que enfrenta por el deterioro de su hábitat y la captura indiscriminada (Bermúdez et al., 2002).
Conocer los parámetros de crecimiento y tasas de mortalidad de las poblaciones de una especie es de gran importancia para generar estrategias de manejo y conservación adecuadas, ya que estos permiten evaluar cómo la extracción u otro tipo de actividades antrópicas pueden afectar las poblaciones. Lastimosamente son pocos los estudios acerca de estos aspectos para las poblaciones de C. guanhumi. En condiciones de laboratorio Henning (1975) estimó que un individuo con 98 mm de ancho del caparazón puede tener 13 años de edad y solo se han realizado dos estudios sobre la determinación de los parámetros de crecimiento poblacionales de esta especie, ambos en Brasil. El primero, se llevó a cabo en un lugar en el que se explota este cangrejo, allí se estimó un K= 0.594 años-1 (Botelho et al., 2001); por otro lado, el segundo estudio se llevó a cabo en un área en donde no ocurren capturas comerciales, y el K estimado fue de 0.145 años-1 (Costa, 2016), lo que en ambos casos permite evidenciar un crecimiento lento para esta especie. Teniendo en cuenta esto, el objetivo de este estudio es estimar los parámetros de crecimiento de las poblaciones de C. guanhumi en áreas protegidas dentro del departamento del Magdalena, en el Caribe Colombiano, a partir de las frecuencias de tallas con el fin de obtener información relevante para el manejo de esta especie.
MATERIALES Y MÉTODOS
Área de estudio
Se eligieron cinco localidades en el Caribe colombiano pertenecientes a dos parques naturales en el departamento del Magdalena: El Centro de Visitantes en la Vía Parque Isla Salamanca (VIPIS), y Bahía Concha, Bahía Nenguange, Cabo San Juan del Guía y Los Naranjos en el Parque Nacional Natural Tayrona (PNNT) (Figura 1). Estos lugares a pesar de tratarse de áreas protegidas, constituyen un atractivo turístico en los que se han llevado a cabo modificaciones ambientales, relacionadas con la vivienda, el transporte o el turismo, y en donde, a pesar de los esfuerzos de las entidades pertinentes, puede llegar a darse la extracción ilegal de este cangrejo, sin embargo, se desconoce la intensidad con la que esta se da.
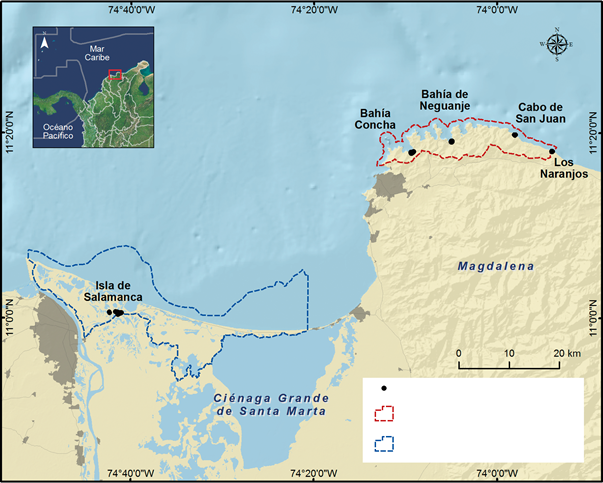
Estructura de la población
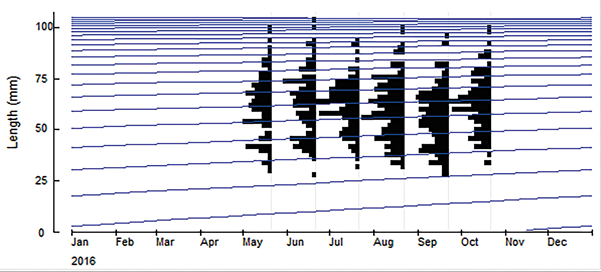
Mensualmente, entre mayo y octubre de 2016, se recolectaron individuos en cada una de las localidades. Para capturar los cangrejos se usaron pinzas de cocina y salabardos (redes) los cuales se lanzaban sobre el individuo de tal forma que éste quedara atrapado en la red; esta actividad se llevó a cabo en horas de la noche, ya que en las salidas de reconocimiento fue posible observar que la actividad de los cangrejos se incrementaba durante este periodo de tiempo. Adicionalmente se elaboraron trampas con botellas plásticas con un diámetro de entrada de 7.62 cm y 10.16 cm, las cuales se dejaron durante la noche con pollo crudo o mango como carnada, y se revisaban al otro día. A cada individuo capturado se le tomaron las siguientes medidas biométricas usando calibrador y balanza: ancho (AC) y largo (LC) del caparazón, y peso. Cada cangrejo se sexó teniendo en cuenta la forma del abdomen: triangular o V invertida en los machos, y redondeado en las hembras. Para estas últimas se anotó si estaban ovadas o no.
A partir de los datos de estructura se elaboraron histogramas de frecuencia de tallas para machos y hembras. Se realizó una prueba Chi-cuadrado, con un nivel de confianza del 95% (Sokal y Rohlf, 1979) para conocer si la proporción de machos y hembras difería significativamente de 1:1.
Parámetros de crecimiento
Se estimó la relación entre la talla y el peso para machos y hembras a través de un análisis de regresión, empleando la ecuación potencial:
P = a*ACb (Ricker, 1975)
Dónde P corresponde al peso total (g), AC es el ancho del caparazón (mm), el parámetro a es el intercepto y b es la pendiente de la curva; estos datos se transformaron con logaritmo natural y se probó la hipótesis nula (H0: b = 3) con la prueba t de "student" (α = 0.05). El parámetro b representa el coeficiente de alometría, de tal forma que cuando b = 3, el crecimiento es isómetrico, y cuando b ≠ 3 el crecimiento es alométrico. Debido a que se usó el AC en milímetros y no en centímetros, el intercepto fue corregido usando a= a'*10b (Froese, 2006). Para este análisis se excluyeron aquellos individuos a los que les faltaba alguna extremidad.
Para describir el crecimiento de los cangrejos se usó la ecuación de crecimiento de von Bertalanffy, (1938):
AC t = AC∞ (1-e-(K(t-t0)),
donde ACt es el ancho de caparazón a la edad t, AC∞ es el ancho de caparazón asintótico, es decir la talla máxima que la especie puede alcanzar, K es el parámetro de curvatura (ó coeficiente de crecimiento), con el que se puede determinar qué tan rápido puede ser alcanzado el ancho de caparazón asintótico y t0 representa la edad teórica a la cual el individuo tiene la talla cero (AC0). Para calcular el ancho de caparazón asintótico (AC∞) se usó la fórmula AC∞ = ACmax /0.95 (Pauly, 1984), en donde ACmax es el ancho de caparazón máximo registrado durante los muestreos. Posteriormente, para estimar el parámetro de curvatura K, se usó la distribución de frecuencias de tallas (ancho de caparazón) mensuales de machos y hembras por separado, los cuales se agruparon en clases de 2 mm. Estos datos, junto al AC∞ calculado, se analizaron con la subrutina K scan, dentro de la rutina ELEFAN I (Electronic Length Frequency Analysis) (Pauly, 1987; Pauly y David, 1981), incorporada en el programa FiSAT II (FAO-ICLARM Stock Assessment Tools II) (Gayanilo et al, 2005). Posteriormente se calculó t0 mediante la ecuación empírica de (Pauly, 1979): log (-t0)= -0.3922-0.2752*(log ACo ∞-1.038*logK).
La rutina ELEFAN, ha sido usada ampliamente debido a que es un método de bajo costo que solo depende de las mediciones regulares de las tallas de los individuos, y a que no es necesario conocer las edades, lo cual puede llegar a ser particularmente difícil en los crustáceos debido a que las estructuras de crecimiento permanentes son escasas en dicho grupo.
Posteriormente, se calcularon las edades correspondientes a las distintas tallas, a través de la ecuación de von Bertalanffy invertida:
t = t0 - (1/k) * 1n (1-ACt / AC ∞ )
Finalmente se estimó la longevidad usando la siguiente ecuación:
t max = t0+ 3/k (Taylor, 1958).
Se hizo un análisis de los parámetros de crecimiento a nivel departamental, para lo cual se usaron los datos de todas las localidades muestreadas y se realizó una estimación total y otra teniendo en cuenta el sexo. Adicionalmente, se estimaron los parámetros de crecimiento para cada uno de los parques por separado con el fin de observar si existían diferencias entre estos. De esta forma para el análisis del PNNT se excluyeron los datos de la localidad de VIPIS. Desafortunadamente no fue posible llevar a cabo un análisis para VIPIS debido al tamaño muestral de esta localidad. Por lo cual solo se comparan los parámetros a nivel departamental con los obtenidos para el PNNT.
Posteriormente, se estimó el índice de desempeño de crecimiento phi prima (Ø'), el cual tiende a ser similar en stocks de una misma especie e incluso en especies de una misma familia (Munro y Pauly, 1983). Este se calculó a partir de los parámetros de crecimiento K y AC∞:
Ø' = logK+2logAC∞. (Pauly y Munro, 1984)
También se calculó (Ø’) para otras poblaciones de C. guanhumi, usando los datos disponibles en publicaciones y se comparó con el estimado para el departamento del Magdalena. Los datos de las frecuencias de tallas también se usaron para calcular la tasa de mortalidad total (Z), a través de la rutina de la curva de captura linealizada, en FISAT, la cual se basa en la ecuación:
Ln(Ni/Δti) = a+b*ti (Pauly, 1983)
donde Ni es el número de individuos capturados en un intervalo de clase i, Δti es el tiempo requerido por el cangrejo para crecer a través de la longitud de clase i, ti es la edad correspondiente al intervalo de clase i; los parámetros a y b son calculados por medio de un análisis de regresión, en los que la pendiente b, con el signo cambiado, corresponde a la Z estimada. En el programa se consideraron aquellos individuos viables para captura, es decir aquellos a partir del punto en el que la curva de captura comienza a bajar a un ritmo constante (Pauly, 1984); y por ende la regresión para calcular Z se llevó a cabo usando este grupo de individuos. El cálculo de la tasa de mortalidad natural (M), se basó en la ecuación de Pauly (1980): , donde T es la temperatura media anual del lugar donde se recolectaron los individuos (28°C ); y el cálculo de la mortalidad por pesca (F) se realizó a través de la fórmula F=Z-M. La estimación de la tasa de mortalidad se hizo para la población general, ya que la extracción de C. guanhumi no es selectiva de sexo.
RESULTADOS
En total se recolectaron 816 individuos durante los seis meses de muestreo. De estos, 181 en Bahía Concha, 154 en Cabo San Juan del Guía, 186 en Nenguange, 183 en los Naranjos y 112 en el centro de visitantes del Vía Parque Isla Salamanca (VIPIS). Exceptuando la localidad de VIPIS, en todas las localidades los individuos mostraron una alta actividad durante la noche, esto hizo que VIPIS tuviera el tamaño muestral más pequeño.
Se llevó a cabo una correlación de Spearman (Sokal y Rohlf, 1979) entre el ancho y el largo del caparazón, en el cual se evidenció que estas medidas se encuentran fuertemente asociadas (coeficiente de correlación: 0.99, p < 0.001), por lo cual solo se usó el ancho de caparazón (AC) para los demás análisis.
La distribución de las frecuencias de tallas (Figura 2) muestra que en general los machos presentan una distribución más amplia, entre 20.2 y 103.7 mm (61.35 ± 0.87 mm) (promedio ± error estándar), mientras que en las hembras estuvo entre 25.9 y 91.9 mm (57.66 ± 0.50 mm); en las hembras se observó una distribución modal de 62-64 mm, mientras que en los machos no fue posible observar una distribución modal clara. Por otro lado, los pesos de los machos estuvieron entre 4 y 516 g (133.49 ± 5.21 g), y los de las hembras entre 9 y 266 g (95.90 ± 2.12 g). La proporción sexual difirió significativamente de 1:1 (x2= 20.08, p < 0.001), con un predominio de los machos.
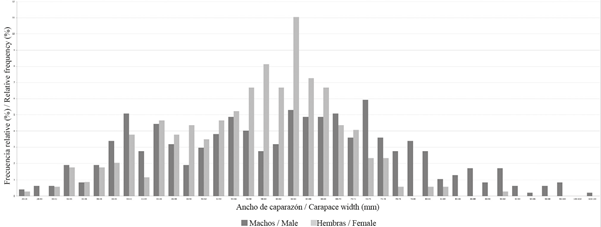
Figura 2 Histograma de la frecuencia de tallas para machos y hembras de Cardisoma guanhumi recolectados entre mayo y octubre de 2016 en áreas protegidas dentro del departamento del Magdalena.
La relación talla peso tanto para machos como para hembras presentó un crecimiento alométrico negativo (b = 2.93, IC = 2.89 - 2.97, t-student = 3.186, p = 0.002 para machos y b = 2.80, IC = 2.74 - 2.86, t-student = 6.397, p < 0.001 para hembras) (Figura 3).
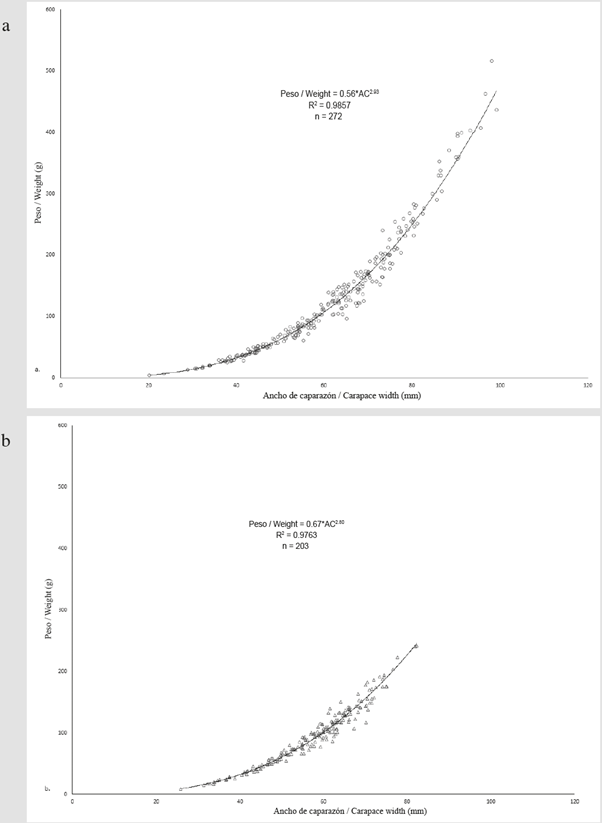
Figura 3 Relación entre el ancho del caparazón y el peso para machos (a) y hembras (b) de C. guanhumi recolectados entre mayo y octubre de 2016 en áreas protegidas dentro del departamento del Magdalena.
Para los parámetros de crecimiento de C. guanhumi en el departamento del Magdalena, se obtuvo un ACmax superior en los machos qué en las hembras, por lo cual el AC∞ estimado fue mayor para los machos. Al ingresar los valores respectivos de AC∞ en la subrutina K scan de ELEFAN, los machos tuvieron un valor de K menor al de las hembras. Esto indica que los machos tienen un crecimiento más lento y un tiempo de vida más largo que el de las hembras (Tabla 1 y Figura 4). En cuanto a los parámetros estimados para el PNNT se obtuvo un valor de K superior y una longevidad menor que los obtenidos para el departamento (teniendo en cuenta tanto los individuos del PNNT como los de la VIPIS) (Tabla 2)
Tabla 1 Parámetros de crecimiento para machos y hembras de Cardisoma guanhumi en el departamento del Magdalena. n= número de individuos muestreados, ACmax= ancho de caparazón máximo obtenido en el estudio, AC∞= ancho de caparazón asintótico, K= parámetro de curvatura, t0= la edad teórica en la que el individuo tiene talla 0, tmax= edad máxima, Ø′=índice de desempeño de crecimiento.
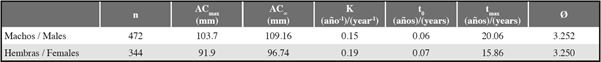
Tabla 2 Parámetros de crecimiento de Cardisoma guanhumi estimados para: a) el departamento del Magdalena (teniendo en cuenta todas las localidades) y b) el Parque Nacional Natural Tayrona (PNNT), para lo cual se excluyeron los datos de la localidad de VIPIS. n= número de individuos muestreados, ACmax= ancho de caparazón máximo obtenido en el estudio, AC∞= ancho de caparazón asintótico, K= parámetro de curvatura, t0= la edad teórica en la que el individuo tiene talla 0, tmax= edad máxima, Ø′=índice de desempeño de crecimiento.
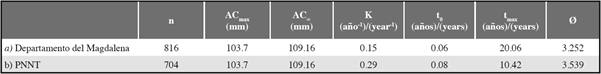

Figura 4 Curva de crecimiento de von Bertalanffy para machos y hembras de Cardisoma guanhumi capturados en las distintas localidades del departamento del Magdalena. Las curvas de crecimiento se graficaron hasta la edad del ACmax obtenido en el estudio.
Con esta misma rutina, se graficaron las distribuciones de las frecuencias mensuales del ancho del caparazón para la población del departamento del Magdalena y se observó que no hay una progresión modal, ya que no es posible visualizar cohortes (Figura 5). El índice de desempeño de crecimiento (Ø’) fue similar en machos y hembras. La tasa de mortalidad total estimada para la población del departamento fue Z= 0.41 año-1, la de mortalidad natural M= 0.36 año-1 y la de mortalidad por pesca F= 0.05 año-1.
DISCUSIÓN
La distribución de las frecuencias mensuales fue multimodal, ya que no fue posible observar la presencia de cohortes a lo largo de los meses; esto ha sido previamente observado para esta especie y ha sido atribuido al hecho de que el reclutamiento de juveniles es continuo y al crecimiento lento característico de la especie (Silva et al., 2014; Costa, 2016).
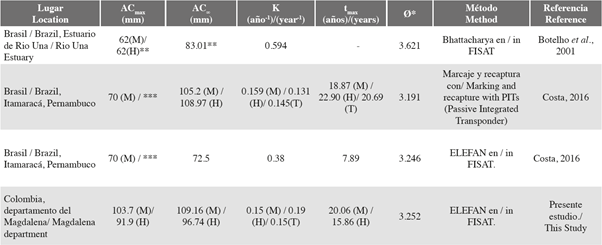
Fue posible evidenciar que los machos alcanzaron tallas más grandes, tuvieron un crecimiento más lento y una longevidad mayor que la de las hembras. Costa (2016) mediante el uso de un método de marcaje y recaptura obtuvo K = 0.159 año-1 para los machos, muy similar al obtenido para este estudio (K = 0.150 año-1). Sin embargo, esta autora obtiene que las hembras crecen más lento que los machos (Tabla 3). Es posible que esto esté relacionado con el AC∞ estimado en dicho estudio, el cual fue mayor para las hembras que para los machos (Tabla 3), lo cual es inusual ya que ha sido reportado ampliamente que los machos de las especies de la familia Gecarcinidae alcanzan tallas y pesos mayores que las hembras, las cuales invierten más energía en los procesos reproductivos (Hartnoll, 1982, 1988).
Como se mencionó previamente hay tres aproximaciones al crecimiento de esta especie: una en condiciones de laboratorio (Henning, 1975), y otras dos en ambientes naturales. En la primera de éstas Botelho et al. (2001) usó el método de Bhattacharya (el cual también se basa en las frecuencias de tallas) y en la segunda Costa (2016) realizó una comparación entre dos metodologías: ELEFAN y marcaje y recaptura (Tabla 3). En ambos estudios, los K estimados fueron más altos que los del presente trabajo, esto posiblemente está relacionado con las tallas máximas reportadas y a las medidas de longitud usadas, factores importantes en la estimación de los parámetros de crecimiento en los análisis de frecuencias de tallas. Por ejemplo Botelho et al. (2001), cuyos muestreos se realizaron en una localidad en la cual C. guanhumi era explotada, estimaron los parámetros de crecimiento a partir de las medidas del LC y no del AC, allí reportaron un LCmax = 62 mm, lo que pudo llevar a que se calculara un K alto.
Tabla 3 Comparación de los parámetros de crecimiento estimados para Cardisoma guanhumi respecto a la literatura. M= machos, H= hembras, T= total. AC∞= ancho de caparazón asintótico, K= coeficiente de crecimiento, tmax= tiempo de vida teórico, índice phi prima. *Datos calculados a partir de la información reportada por los autores. Para estimar tmax se utilizó: tmax= t0 + (3/K) (Taylor, 1958), si t0 no estaba reportado, se usaba 0 en la ecuación. Para phi prima se usó: Pauly y Munro (1984). **Medidas de largo del caparazón (LC). Para los datos de Botelho et al. (2001) no se estimó tmax debido a que los autores calcularon K a partir de los valores de LC. *** Dato no reportado por el autor.
Por otro lado Costa (2016), cuyo ACmax fue de 70 mm, estimó dos valores para K, el primero resultado de un análisis de ELEFAN (n=1078) y el segundo usando datos de marcaje y recaptura (n=291). De esta forma, Costa (2016) obtuvo un K mayor con ELEFAN, en comparación con el K estimado a partir de los datos de marcaje y recaptura. Esto puede explicarse porque para estimar el parámetro K, la rutina ELEFAN se basa en el AC∞ (el cual se calcula a partir del ACmax adquirido en el muestreo). Es posible que durante el muestreo, Costa (2016) no haya podido capturar un individuo con un ACmax representativo de la población (es probable que existieran tallas más grandes considerando que los muestreos se llevaron a cabo en un área con vigilancia donde no ocurren capturas comerciales de C. guanhumi). De esta forma, al calcular un AC∞ más pequeño se sobreestima el valor de K. Mientras que con los datos de marcaje y recaptura, Costa (2016) pudo estimar el aumento de la talla a partir de los cambios directos en el crecimiento de los cangrejos, y por lo tanto el parámetro K no se vio afectado por el ACmax obtenido en el muestreo.
A partir de los parámetros de crecimiento del PNNT, C guanhumi presentó un crecimiento más rápido (K= 0.29 año-1) y una menor longevidad (tmax = 10.42 años) que a nivel departamental (K= 0.15 año-1 y tmax= 20.06 años). A pesar de que tanto el PNNT como VIPIS se encuentran dentro del departamento del Magdalena, es posible que las diferencias obtenidas al analizar por separado los datos del PNNT se encuentren relacionadas a la cercanía entre las cuatro localidades de este parque, ya que, al estar más cerca, es probable que estas poblaciones se encuentren más emparentadas y por tanto presenten más características en común, como lo que respecta al crecimiento. Adicionalmente durante los muestreos se evidenció que los individuos de C. guanhumi de la VIPIS mostraron un comportamiento distinto al de las demás localidades, ya que durante la mayoría de los meses los individuos presentaron poca actividad por fuera de las madrigueras, lo que restringió el tamaño muestral en esta localidad.
Los índices de desempeño de crecimiento (Ø’) para el departamento fueron similares a los que se calcularon para los datos de marcaje y recaptura de Costa (2016), los cuales fueron de 3.246 y 3.191 para machos y hembras respectivamente (Tabla 3). Por otro lado, el Ø’ calculado para los datos de Botelho et al. (2001) fue de 3.621, valor ligeramente más alto que los demás (Tabla 3). Sin embargo, como se mencionó previamente, estas diferencias pueden estar relacionadas al hecho de que en dicho estudio se usara el LC y no el AC en los análisis de frecuencias de tallas.
La tasa de mortalidad total (Z = 0.41 año-1), la mortalidad natural (M = 0.36 año-1) y la mortalidad por pesca (F = 0.05 año-1) fueron bajas en el departamento, lo cual es consistente con el hecho de que los muestreos se llevaron a cabo en áreas protegidas. Cabe anotar que para la localidad de Bahía Concha se han reportado eventos de extracción de C. guanhumi, por lo que es necesario llevar a cabo un estudio enfocado en dicha localidad para evaluar si la mortalidad ocasionada por la extracción es alta en este lugar. Costa (2016) estimó una mortalidad total Z = 2.31 año-1, a partir de los datos de capturas mensuales y los valores de AC∞ y K que obtuvo con los datos de marcaje y recaptura (108.03 mm y 0.145 año-1). Esta alta mortalidad en Costa (2016) pudo estar relacionada con la diferencia entre el AC∞ (108.03 mm) y el ACmax (70 mm) lo que se interpretaría como una mortandad alta en los individuos adultos. Esta autora no determinó la mortalidad por pesca, ya que trabajó en un área en donde no son capturados comercialmente y por ende asumió Z=M.
A partir de los parámetros de crecimiento estimados en el presente estudio, y de los obtenidos por Costa (2016) teniendo en cuenta que los datos de marcaje y recaptura brindan información más exacta sobre el crecimiento, es posible inferir que C. guanhumi es una especie de crecimiento lento y longeva, con una esperanza de vida de más de 10 años. Estas características también han sido reportados en otros crustáceos terrestres de gran tamaño como Ucides cordatus, el cual se ha estimado que puede vivir hasta 17 años (Diele y Koch, 2010).
También debe tenerse en cuenta que las diferencias en los parámetros de crecimiento entre las distintas áreas de estudio pueden no solo estar relacionadas a la metodología usada, sino que pueden deberse a otros factores que pueden influenciar sobre la rapidez del crecimiento y la longevidad, como la disponibilidad de comida, la calidad del hábitat, y la extracción, entre otros, los cuales pueden generar diferencias entre poblaciones de una misma especie. Esto también puede explicar las diferencias obtenidas en el valor de K obtenido a nivel departamental con respecto al K obtenido para el PNNT. De igual forma es posible que si se evaluaran por separado las localidades al interior del PNNT, se observasen diferencias en el crecimiento de los cangrejos entre estas.
Por otro lado debe considerarse la implementación de métodos de captura y recaptura para obtener datos más robustos en los análisis de crecimiento en especies como C. guanhumi caracterizadas por tener un crecimiento lento, ser longevas y con un reclutamiento constante de juveniles (Silva et al., 2014; Costa, 2016), lo que puede presentar dificultades al usar métodos basados en las frecuencias de tallas.
CONCLUSIONES
En general la población de C. guanhumi estudiada mostró un crecimiento alométrico negativo, es longeva, y de crecimiento lento, lo que coincide con lo previamente reportado para esta especie. Esto es de gran importancia porque cuando una especie de crecimiento lento es sometida a una extracción intensiva, requiere más tiempo para recuperar sus stocks (Silva et al, 2014). Esto debe tenerse en cuenta a la hora de realizar planes de manejo para el aprovechamiento responsable de este cangrejo. Adicionalmente se obtuvo una baja mortalidad por pesca (extracción), lo que indica que, desde una perspectiva general para la zona muestreada, la especie no está siendo sobreexplotada.

