text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los cocolitóforos son microalgas marinas, unicelulares, pertenecientes a la división Haptophyta, que normalmente presentan una estructura externa compuesta de diminutas placas (<30 μm) de carbonato de calcio conocidas como cocolitos (Jordan et al., 1995). Una vez que el microorganismo muere, los cocolitos se hunden en la columna de agua, ya sea en forma de pellets fecales o fragmentos desarticulados incluidos en la "nieve marina", alcanzando el fondo oceánico donde pasan a constituir parte del sedimento (Young et al., 2003; Flores y Sierro, 2007). Una vez que se encuentran en las rocas, los cocolitóforos se incluyen en el grupo de nanofósiles calcáreos, incluyendo algunas formas similares que han sido observadas en sedimentos tan antiguos como el Triásico tardío (Bown, 1998). Los cocolitóforos han sido observados desde los océanos polares hasta los mares tropicales como parte del fitoplancton marino, habitando exclusivamente en la zona fótica (Flores y Sierro, 2007). Debido a su alta abundancia y capacidad de llevar a cabo procesos como la fotosíntesis y la calcificación, los cocolitóforos juegan un papel fundamental en la regulación del sistema océano-atmósfera a través de los ciclos biogeoquímicos del CO2 del planeta (Thierstein y Young, 2004). El estudio de estos microorganismos y su registro fósil ha ganado importancia en investigaciones paleoclimáticas y paleoceanográficas, ya que se ha demostrado su alta susceptibilidad ante los cambios de temperatura superficial marina, disponibilidad de nutrientes, luminosidad, y salinidad superficial; asimismo su uso como herramienta bioestratigráfica es habitual y de gran utilidad (Giraudeau, 1992; Flores et al., 1997, 1999, 2000; Balch, 2004; Antunes, 2007; Saavedra-Pellitero et al., 2010, 2011; Poulton et al., 2017). El océano Pacífico ecuatorial colombiano, lugar donde se desarrolla este trabajo, presenta características oceanográficas particulares, entre las que se destacan: 1) Sus altas tasas de precipitación (2000 a 12700 mm/año) que alimentan en alta medida los cauces de los ríos San Juan, Patía y Mira, entre otros. Estos afluentes de agua dulce arrastran una gran cantidad de sedimentos calculado en 35x109 m3/año, los cuales afectan considerablemente las propiedades físico-químicas de las aguas superficiales marinas tales como salinidad, turbidez, concentración de nutrientes y temperatura (Poveda y Mesa, 2000; Restrepo y Kjerfve, 2000; CCCP, 2002; Betancur y Martínez, 2003; Gómez y Martínez, 2005; López y Costeros, 2006; Restrepo, 2006; Restrepo y López, 2008; Patarroyo y Martínez 2013, 2016); 2) La dinámica estacional de surgencia oceánica, ocasionada por corrientes de bajo nivel que atraviesan el istmo panameño desde enero hasta marzo, producen un enfriamiento de las aguas superficiales y un incremento en la concentración de nutrientes debido al ascenso de aguas más profundas hacia la superficie (D'Croz y O'Dea, 2007, 2009).
El presente estudio tiene como objetivo determinar la composición y distribución de las asociaciones cuaternarias de cocolitos analizados en los 39 core tops extraídos del fondo marino de los departamentos de Chocó y Nariño (Figura 1). Esta información permitirá establecer un ámbito de tiempo y conocer el patrón de distribución de las comunidades de cocolitóforos que habitan en estas aguas marinas tropicales cercanas a la costa del Pacífico colombiano. Adicionalmente, esta información es un punto de partida para las bases de datos que pretendan analizar la composición del nanoplancton calcáreo en el territorio colombiano.
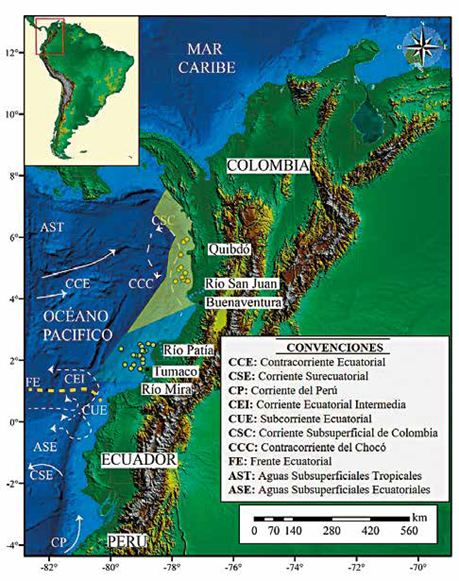
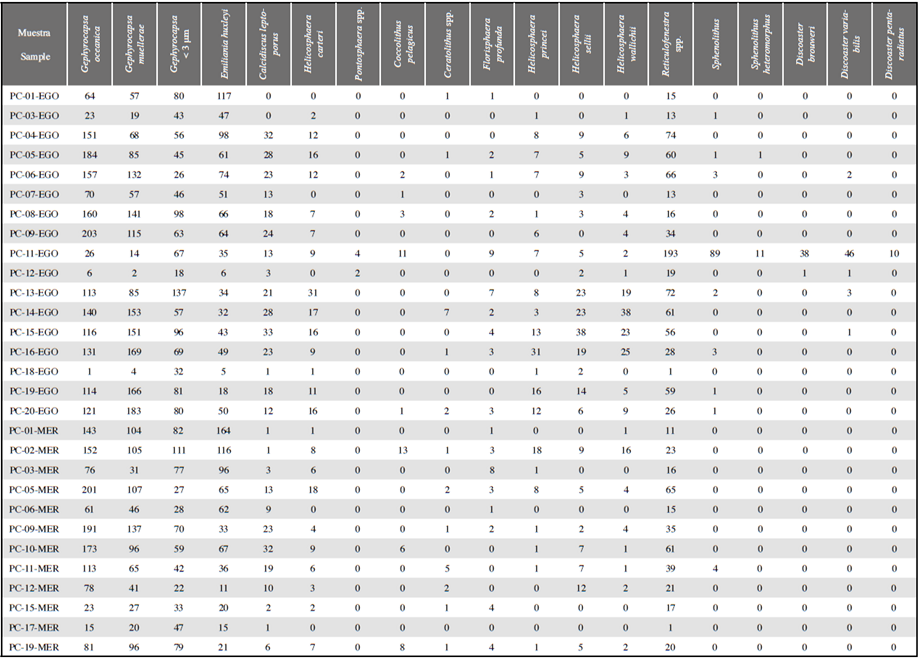
Figura 1 Algunas características de las corrientes oceánicas en el océano Pacífico colombiano. Las corrientes superficiales están representadas por las líneas continuas de color blanco; las líneas discontinuas en color blanco indican las corrientes subsuperficiales; el Frente Ecuatorial con líneas discontinuas color naranja. También se representan las aguas superficiales tropicales y ecuatoriales. Los polígonos amarillo y azul delimitan las áreas de las Cuencas Chocó y Tumaco respectivamente, y los puntos amarillos indican la localización de los pistón core. Mapa realizado integrando la información registrada en Wyrtki (1966), Restrepo y Correa (2002), Fiedler y Talley (2006), Restrepo y López (2008) y Rodríguez-Rubio et al. (2008).
ÁREA DE ESTUDIO
El Pacífico tropical oriental es una región caracterizada por la confluencia de corrientes marinas provenientes del norte (Corriente de California) y del sur (Corriente de Perú), dando origen a una zona fronteriza conocida como Frente Ecuatorial, la cual limita la circulación entre aguas superficiales templadas del norte y ecuatoriales frías de alta salinidad del sur (Wyrtki, 1965, 1966; Pak y Zaneveld, 1974; Okada, 1983; Fiedler y Talley, 2006). El océano Pacífico en Colombia se caracteriza por tener una importante dinámica océano- atmósfera en la que convergen los vientos alisios del norte y del sur, controlados por el desplazamiento de la Zona de Convergencia Intertropical (Wyrtki, 1965, 1966; Pak y Zaneveld, 1974; Fiedler y Talley, 2006). De acuerdo con Knauss (1960), Wyrtki (1966) y Fiedler y Talley (2006), las corrientes oceánicas que influyen en la dinámica superficial del Pacífico colombiano son: (a) Contracorriente Ecuatorial (CCE) de aguas tropicales-subtropicales con alta concentración de nutrientes (>25 °C y <34 UPS); (b) Corriente Surecuatorial (CSE) que transporta considerables volúmenes de agua cálida hacia el oeste, y su límite hacia el norte es con la CCE; y (c) Corriente del Perú (CP), también conocida como Corriente de Humboldt, caracterizada por sus aguas frías templadas y eutróficas (15 a 28 °C y >36 psu). Por otra parte, entre las corrientes subsuperficiales que circulan en el Pacífico colombiano se encuentran la Subcorriente Ecuatorial o de Cromwell y la Corriente Ecuatorial Intermedia (Restrepo y Correa, 2002). En adición a éstas, se registran dos corrientes descritas por Restrepo y López (2008) y Rodríguez-Rubio et al. (2008), en el margen continental del Pacífico colombiano, que son: la Corriente Subsuperficial de Colombia (CSC) y la Contracorriente del Chocó (CCC).
Las salinidades y temperaturas en el océano Pacífico colombiano se comportan de forma inversa, observándose que en las regiones donde existe una temperatura alta, las salinidades tienden a ser bajas (CCCP, 2002). La temperatura marina superficial ha oscilado entre 24,5 y 29,8 °C durante los últimos 100 años (Málikov y Villegas, 2005; NOAA, 2017), siendo mayor en las regiones proximales a la costa y menor hacia las más distales (Figura 2). Las aguas superficiales tienden a presentar una temperatura superficial más alta en la región de Chocó que en Tumaco, esto se debe principalmente a que la Contracorriente del Chocó transporta aguas cálidas provenientes del norte (CCCP, 2002). Este patrón se relaciona estrechamente con los cambios de salinidad en el Pacífico colombiano, con ámbitos en la costa entre 30 y 32,8 UPS y valores más altos alrededor de 35 psu a medida que se aleja hacia el sur y occidente (CCCP, 2002) (Figura 2).
MATERIALES Y MÉTODOS
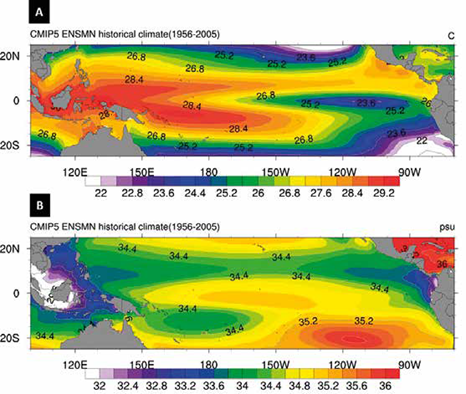
Entre el 25 de noviembre y el 11 de diciembre de 2009 la compañía GEMS-HRT y la DIMAR realizaron para Ecopetrol S.A. la toma de muestras del fondo marino en las áreas de evaluación técnica (TEA's) de Merayana (Cuenca Chocó) y Egoro (Cuenca Tumaco), en áreas offshore de los departamentos de Chocó y Nariño respectivamente. La adquisición de 39 pistón core se realizó desde la embarcación ARC Providencia a una distancia promedio de 50 km medida a partir de la línea de costa. Los puntos fueron seleccionados a partir de información sísmica, imágenes batimétricas y la señal de un penetrador de sedimentos de alta frecuencia de 3,5 KHz. Las muestras usadas en este estudio pertenecen a los primeros 11 cm (techo a base) de los 39 core tops, distribuidas de la siguiente manera: 18 piston core en la Cuenca Tumaco (TEA Egoro) y 21 piston core en la Cuenca Chocó (TEA Merayana) (Figura 3; Tabla 1). Cabe aclarar que los primeros centímetros del material sedimentario recuperado podrían haber experimentado mezcla de sedimentos, como consecuencia del mismo uso de la técnica de piston core.
Figura 3 Localización de las zonas de muestreo en las Cuencas Chocó y Tumaco. Nótese la especial localización de la muestra PC-11-EGO sobre un alto topográfico del fondo oceánico.
Tabla 1 Información de las muestras de piston core obtenidos en el Pacífico colombiano. Las muestras que no presentaron ningún contenido de cocolitóforos no se han incluido en esta tabla. *Los valores de las tasas de descarga de sedimentos se han tomado de Restrepo et al. (2000), Restrepo (2006) y Restrepo y López (2008).
Preparación de las muestras
El material colectado se preparó en el Laboratorio de Microfósiles Calcáreos del Instituto de Investigaciones en Estratigrafía (IIES) de la Universidad de Caldas, siguiendo la técnica de decantación propuesta por Flores y Sierro (1997). El uso de este método requiere pesar la cantidad de sedimento a decantar y permite obtener una distribución homogénea de los microfósiles en un área de dimensiones conocidas, facilitando así el cálculo de la abundancia de cocolitos por gramo de sedimento a través de la fórmula:
N= n x R2 x V x r-2 x g-1 x v-1.
En donde n es el número de nanofósiles contados en un área determinada, R el radio de la placa Petri donde se ha decantado la muestra, V el volumen de agua que se ha añadido al sedimento seco durante la preparación, r el radio del campo visual del microscopio utilizado en el recuento, g el peso del sedimento seco y v el volumen de solución añadido con la micropipeta.
Para el registro fotográfico se usó una cámara Nikon Digital Sight DS-Ri1 con un controlador DS-U2 acoplados a un microscopio petrográfico Nikon Eclipse 50i POL, usando el software Imaging NIS-Elements versión 3.2 de Nikon.
Con el fin de obtener fotografías de mayor resolución y detalle, las muestras más abundantes fueron preparadas para la toma de imágenes al Microscopio Electrónico de Barrido (SEM).
Análisis microscópicos cualitativos y cuantitativos
Después de la preparación de 39 placas para el estudio de cocolitóforos se hizo su análisis por medio de un microscopio petrográfico con un aumento de 1000x. Este análisis se dividió en dos partes, inicialmente se hizo una estimación cualitativa del estado de preservación de los cocolitos de acuerdo con los criterios establecidos por Roth y Thierstein (1972), Roth (1983) y Raffi y Flores (1995) en donde: (B) Buena: Los ejemplares presentan poca o ninguna disolución y/o recristalización. (B-M) Buena-Moderada: Muestra ligera a moderada disolución y/o recristalización y la identificación de las especies es limitada. (M) Moderada: Los ejemplares presentan disolución moderada y/o recristalización excesiva. (P) Pobre: Los microfósiles presentan una disolución extrema y/o recristalización excesiva.
Posteriormente se procedió a hacer el reconocimiento taxonómico de todos los grupos presentes, siguiendo la guía taxonómica de Young et al. (2003) para las asociaciones de cocolitos del Pleistoceno y las descripciones hechas por Perch-Nielsen (1985) para las formas retrabajadas del Mioceno-Plioceno. Por otra parte, se agruparon bajo el nombre de Gephyrocapsa <3 μm los cocolitos de Gephyrocapsa que se caracterizaron por tener tamaños inferiores a 3 μm y que por su diminuto tamaño al microscopio petrográfico no fueron identificados. Una lista de los taxones encontrados en las muestras es presentada en el Apéndice A.
Finalmente se realizó un análisis cuantitativo por medio de dos tipos de recuentos: el primero, un recuento de 500 especímenes por muestra, independientemente del número de campos visuales observados. En muestras con bajo número de cocolitos se contó un máximo de 400 campos visuales. Este método garantiza con un 95% de confianza que los especímenes con abundancias del 1% son incluidos dentro de la asociación (Dennison y Hay, 1967; Fatela y Taborda, 2001). El segundo recuento se hizo en 40 campos visuales, buscando exclusivamente aquellas especies que mostraron abundancia <2%; este recuento se realizó con la finalidad de cuantificar las especies subestimadas o sobreestimadas en el primer conteo (Apéndice B).
Análisis estadísticos
La matriz de abundancias usada para el análisis estadístico fue creada omitiendo aquellas muestras con una abundancia menor de 100 ejemplares y descartando aquellas especies con abundancias inferiores a 2% (Schneider et al, 2011). Esto garantiza que las interpretaciones basadas en los resultados estadísticos contienen los taxones más representativos de la asociación (Dennison y Hay, 1967; Fatela y Taborda, 2001; Schneider et al, 2011).
Adicionalmente, las formas de nanofósiles calcáreos retrabajadas fueron igualmente excluidas del tratamiento estadístico con la finalidad de evitar alteraciones durante el procesamiento.
Se realizó un análisis de redundancia (RDA) para resumir las relaciones entre las especies de cocolitóforos y los parámetros ambientales (Zar, 1996; Hammer et al., 2001; Whitlock y Schulter, 2009; Borcard et al, 2011). Los conteos se estandarizaron a partir de una transformación tipo Hellinger para permitir que los taxones de baja incidencia se ponderarán en el análisis estadístico en el software R core Team, (2015) con el paquete Vegan. La significancia de cada parámetro ambiental fue evaluada con base en la longitud del gradiente en el espacio de ordenación y su correlación con los dos ejes principales. La dirección y tamaño de los vectores son indicadores de la influencia de las variables en la distribución de los grupos taxonómicos.
RESULTADOS
Edad
La presencia de Emiliania huxleyi dentro de la asociación, cuya aparición es situada a partir del Ioniano (Pleistoceno Medio), marca el límite inferior de la biozona NN21 (Martini, 1971). Este bioevento está datado en 290.000 años (Thierstein et al, 1977; Backman et al, 2012). A partir de la asociación de cocolitos encontrada se establece que la edad de estos sedimentos no debe ser más antigua que la biozona NN21 del Ioniano (Pleistoceno Medio).
Asociación y abundancia de cocolitóforos
Los resultados del análisis microscópico mostraron que de las 39 muestras analizadas 9 resultaron estériles, 8 de la Cuenca Chocó y 1 de la Cuenca Tumaco. El resto de las muestras con contenido de microfósiles permitieron identificar 20 taxones de cocolitóforos en ambas localidades (Figura 4).
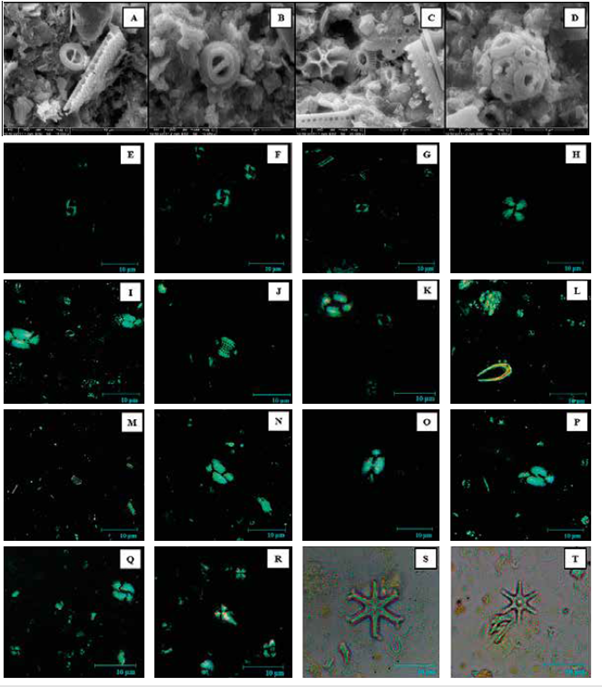
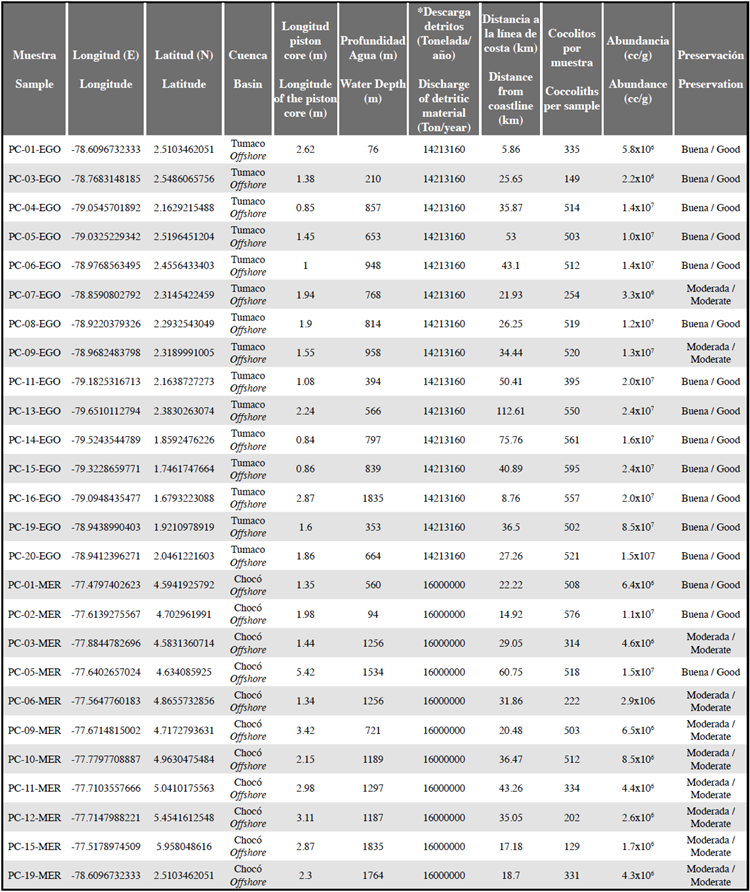
Figura 4 Especies de cocolitos encontrados en las muestras analizadas. Imágenes tomadas bajo el Microscopio Electrónico de Barrido (SEM). A. Gephyrocapsa oceanica (10 μm). B. Gephyrocapsa oceanica (5 μm). C. Emiliania huxleyi (5 μm). D. Coccosphere de Gephyrocapsa spp. (5 μm). Imágenes capturadas bajo microscopio petrográfico y ampliadas por 1000x. Nicoles cruzados (campo oscuro). Nicoles paralelos (campo claro). E. Gephyrocapsa oceanica. F. Gephyrocapsa muellerae. G. Emiliania huxleyi. H. Calcidiscus leptoporus. I. Helicosphaera carteri. J. Pontosphaera spp. K. Coccolithus pelagicus. L. Ceratolithus spp. M. Florisphaera profunda. N. Helicosphaera princei. O. Helicosphaera sellii. P. Helicosphaera wallichii. Q. Reticulofenestra pseudoumbilicus. R. Sphenolithus heteromorphus (inferior) y Sphenolithus abies (superior). S-T. Discoaster variabilis.
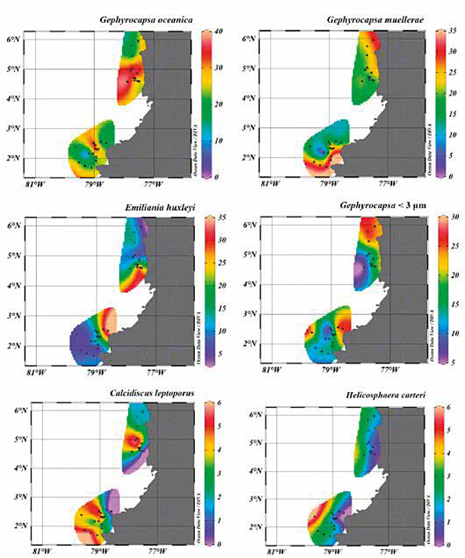
Entre los taxones registrados, G. oceanica, G. muellerae, Gephyrocapsa <3 |im, E. huxleyi, C. leptoporus y H. carteri presentaron abundancias relativas mayores a 2% (Figura 4 E-I; Tabla 2). Por el contrario, especies como Ceratolithus spp, C. pelagicus, F. profunda, H. princei, H. sellii, H. wallichii y Pontosphaera spp. ocuparon abundancias menores a 2% (Figura 4 J-P). Otro componente común de las asociaciones fueron las formas de nanofósiles retrabajados del Mioceno y el Plioceno, principalmente formas de Reticulofenestra spp. (Figura 4 Q). La muestra PC-11-EGO se caracterizó por presentar los valores más altos de retrabajo, observándose D. variabilis, S. abies, y S. heteromorphus. (Figura 4 R-T).
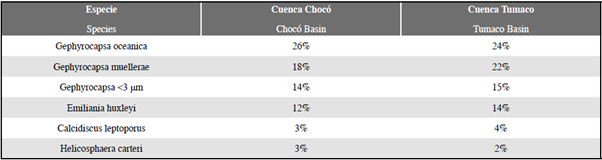
Aunque los especímenes más modernos de cocolitóforos fueron vistos tanto en la Cuenca Chocó como en la Cuenca Tumaco, se evidenció un contraste en cuanto al número de cocolitos por gramo (cc/g) calculados para ambas regiones (Tabla 1). Igualmente, los análisis cualitativos del grado de preservación de los cocolitos variaron entre las dos cuencas, mostrando que en la Cuenca Tumaco la preservación en general fue buena, sin disolución y/o recristalización excesiva; mientras que, los taxones de la Cuenca Chocó comúnmente presentaron una preservación moderada (Tabla 1). No obstante, el reconocimiento a nivel de especies se logró realizar en la mayoría de los ejemplares (Figura 5).
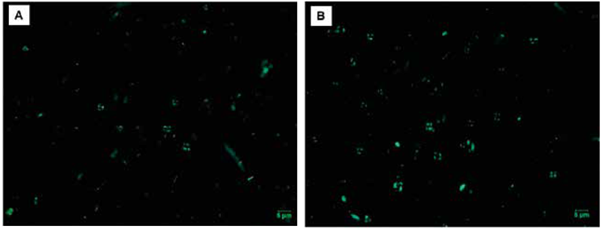
Figura 5 Ejemplo de la abundancia y preservación de los cocolitos. A. muestra PC-19-MER; B. muestra PC-15-EGO. Nótese la diferencia de abundancia y preservación en ambas muestras.
Los cálculos de abundancias de cocolitos por gramo de sedimento evidenciaron que la Cuenca Tumaco presenta los valores más altos, con un promedio de 1,2x107 cc/g. Las abundancias más altas se encuentran en las muestras PC-11-EGO, PC-13-EGO, PC-14-EGO, PC-15-EGO y PC-16-EGO, con valores alrededor de 2,0x107 cc/g, mientras que las muestras con menor abundancia corresponden a PC-12-EGO y PC-18- EGO con valores de 7,9x105 cc/g y 6,8x105, respectivamente. Los puntos muestreados procedentes de la Cuenca Chocó presentan valores inferiores, con un promedio de 5,7x106 cc/g, donde las muestras PC-02- MER y PC-05-MER presentan la mayor abundancia, con valores de 1,1x107 y 1,5x107 cc/g respectivamente. Una menor concentración se observa en las muestras PC-15-MER y PC-17-MER con valores entre 1,2x106 y 1,7x106 cc/g (Figura 6; Tabla 2).
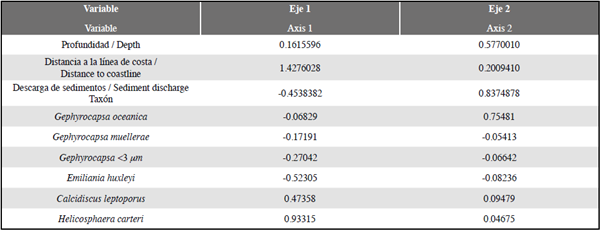
Análisis multivariados
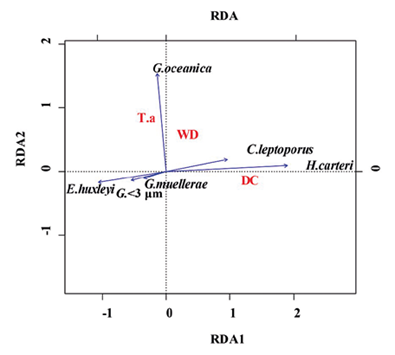
El análisis de redundancia RDA (Zar, 1996; Hammer et al, 2001; Whitlock y Schulter, 2009; Borcard et al, 2011) proporcionó información significativa sobre la relación entre las abundancias de los cocolitóforos, demostrando que el factor más explicativo y/o correlacionable respecto a la distribución y abundancia de los taxones es fundamentalmente la distancia a la línea de costa (eje 1; 1,4276028; DC) en comparación con la profundidad de la columna de agua (eje 2; 0,5770010; WD) y con la descarga de sedimentos (-0,4538382; T.a), (Tabla 3; Figura 7). Estos parámetros explican 74%, 14% y 12% respectivamente de la varianza total. Emiliania huxleyi, Gephyrocapsa <3 μm y G. muellerae revelan una tendencia hacia el eje 1 con valores negativos, mientras que el vector de mayor magnitud sobre este eje está dado por H. carteri y en menor medida por C. leptoporus (Figura 7). En cuanto a la especie G. oceanica, muestra una mayor correspondencia con el eje 2, siendo el vector de este taxón el más cercano a dicho eje, mostrando preferencias por condiciones de mayor profundidad y de mayor afinidad bajo condiciones de alta influencia de sedimentos (Figura 7).
DISCUSIÓN
Comparación con otras asociaciones de cocolitóforos del Pacífico Tropical Oriental
El conteo de cocolitos indica que la asociación principal (mayor a 2%) en las muestras de Chocó y Tumaco es G. oceanica, G. muellerae, Gephyrocapsa <3 μm, E. huxleyi, C. leptoporus y H. carteri (Figura 4; Tabla 2). En una menor proporción (<2%) se observaron Ceratolithus spp., Coccolithus pelagicus, F. profunda, H. princei, H. sellii, H. wallichii y Pontosphaera spp. junto con formas retrabajadas de R. pseudoumbilicus, D. variabilis, S. heteromorphus y S. abies (Figura 4). La asociación dominante registrada en este estudio difiere de los cocolitóforos registrados por Saavedra-Pellitero et al. (2010, 2011) cerca de la dorsal de Cocos y Carnegie (Figura 8). Una de las más marcadas diferencias en esta comparación es el número de ejemplares de F. profunda hacia el occidente en los sedimentos Cuaternarios de la Dorsal de Cocos y al sur en la Dorsal de Carnegie (Ahagon, 1993; Saavedra-Pellitero et al., 2010, 2011).
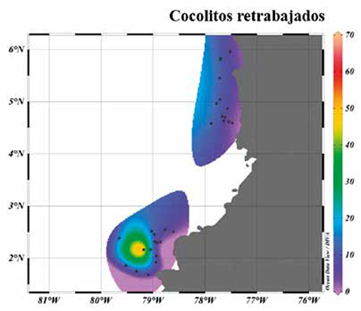
Figura 8 Mapa de distribución geográfica y abundancia relativa de los grupos considerados como retrabajados. Se observa que el área donde se presenta la mayor concentración de estos, corresponde a la muestra PC-11-EGO.
En la Dorsal de Cocos la especie más abundante registrada es F. profunda (34,89%), seguida de G. oceánica (21,98%), Gephyrocapsa <3 μm (18,74%) y G. muellerae (10,75%). En dichas asociaciones, C. leptoporus, E. huxleyi y H. carteri son componentes con abundancias inferiores a 4%. En la Dorsal de Carnegie los grupos más abundantes continúan siendo F. profunda (28,8%) y G. oceanica (26,2%), seguidas de Gephyrocapsa <3 μm (16,1%), E. huxleyi (11%), G. muellerae (10,75%) y C. leptoporus (4,3%) (Figura 9), mientras que en este trabajo se registran abundancias de G. oceanica (24-26%), G. muellerae (18-22%), Gephyrocapsa <3 μm (14-15%), E. huxleyi (1214%), C. leptoporus (3-4%), H. carteri (2-3%) y F. profunda con abundancias por debajo de 2% (Tabla 2). Estos patrones de distribución en los taxones podrían estar reflejando variaciones en las afinidades paleoecológicas de las especies y/o efectos de "sorting" asociados a la disolución. No obstante, discriminar entre los posibles factores que afectaron la distribución de las asociaciones requiere un estudio más exhaustivo que incluya otras herramientas químicas y paleontológicas junto con el uso de técnicas estadísticas.
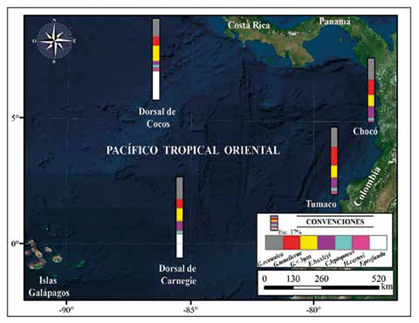
Figura 9 Comparación de las asociaciones y abundancias promedio de cocolitóforos reportadas en piston core tomados sobre la Dorsal de Cocos, la Dorsal de Carnegie (Saavedra-Pellitero 2010, 2011) y el Pacífico colombiano en este trabajo.
Abundancia y preservación de los cocolitos: Chocó vs Tumaco
El análisis descriptivo y cuantitativo de las asociaciones de cocolitos extraídos de las cuencas Chocó y Tumaco muestran una similitud en la composición de la nanoflora (Figura 4-6, Tabla 2). Sin embargo, se observó una diferencia en el grado de preservación y abundancia entre las localidades (Figura 5; Tabla 1). En el caso de la Cuenca Chocó, su preservación fue moderada y el número de cocolitos por gramo fue bajo (5,7x106 cc/g) en comparación con la Cuenca Tumaco. Este valor se debe a que la mayoría de muestras presentaron abundancias muy bajas, 8 de 21 muestras fueron catalogadas como estériles y solamente 11 tuvieron un recobro superior a 100 cocolitos (Tabla 1). En contraste, para la Cuenca Tumaco sólo se obtuvo una muestra estéril, estimándose una buena preservación y un promedio de abundancia de 1,2x107 cc/g (Tabla 1). En un intento por explicar esta diferencia en abundancia y distribución, se realizó un análisis estadístico multivariado RDA evaluando las variables disponibles y que posiblemente controlaron la producción de cocolitos. Dicho análisis reveló que las fluctuaciones de las abundancias de los cocolitos >2% pueden ser explicadas principalmente por la distancia a la línea de costa (Tabla 3; Figuras 6 y 7). Las mayores abundancias que se obtuvieron de E. huxleyi y Gephyrocapsa <3 μm se encontraron en las muestras cuya localización estuvo más cerca de la línea de costa y en zonas menos profundas (i.e. los puntos de muestreo PC-15 MER, PC-19 MER, PC-01-EGO y PC-03-EGO). Aunque H. carteri presentó bajas abundancias (Figura 6), la distribución y abundancia de esta especie parece estar muy controlada por la distancia a la línea de costa como lo sustenta el análisis RDA (Tabla 3), siendo más abundante en las muestras más alejadas del continente (Figura 7). Según el procesamiento estadístico, las variables con más bajos porcentajes de influencia son: la cantidad de detritos por año y profundidad de la columna de agua, estos factores no fueron determinantes en las abundancias de cocolitos calculadas para las muestras examinadas.
Tabla 3 Coeficiente de correlación entre las variables y los puntajes de las especies de cocolitos más representativas en el análisis de redundancia. Los valores en negrilla corresponden a los parámetros que presentaron mejor correlación con los ejes principales.
Modernos estudios en las aguas oceánicas de la Cuenca Chocó indican que la temperatura es mayor hacia la costa, y a su vez es mayor en comparación con la temperatura de la Cuenca Tumaco, en donde, por lo contrario, la salinidad es mayor hacia la zona distal y es más intensa que la presentada en las aguas de la Cuenca Chocó (Figura 2) (CCCP, 2002; NOAA, 2017). Estas características se encuentran estrechamente relacionadas a la influencia de corrientes oceánicas de mayor magnitud, como la Contracorriente del Chocó (con aguas menos frías) en la zona del Chocó, y de la Corriente de Perú (masas de agua más frías) en áreas de Tumaco (CCCP, 2002). Aunque de forma general estas características físico-químicas pueden estar siendo reflejadas por los microfósiles hallados en este estudio, la falta de datos cuantitativos no permite realizar una evaluación del impacto de estas variables sobre la abundancia y preservación de las asociaciones de cocolitóforos.
Presencia de retrabajados
Dentro de los resultados adquiridos a través de la identificación y conteo de los cocolitos de las 39 muestras, fueron identificadas y cuantificadas las formas retrabajadas (Figura 4 Q-T). Los retrabajados más comunes fueron las especies pertenecientes al género Reticulofenestra, las cuales fueron vistas en la mayoría de las muestras. La muestra PC-11-EGO, localizada al sur en la Cuenca Tumaco, mostró el mayor contenido y variedad de especies retrabajadas (Figuras 3 y 9).
Sphenolithus abies, Sphenolithus heteromorphus, Discoaster variabilis, y Reticulofenestra spp. fueron las más abundantes en dicha muestra (Figura 9). La presencia de esta asociación en sedimentos del Pleistoceno permite inferir el aporte de sedimentos marinos del Mioceno-Plioceno al área de estudio. Con base en la localización de la muestra PC-11-EGO en las vertientes de un relieve positivo (Figura 3), se propone que es factible que el origen de estos nanofósiles calcáreos pueda estar relacionado con procesos de remoción del fondo marino, el cual incorporaría formas del Neógeno en la sedimentación Cuaternaria. Esto parece estar apoyado al hecho que ésta fue la única muestra con recuperación de los especímenes más antiguos, notándose que los sitios aledaños no presentaron formas de Sphenolithus y Discoaster. Adicionalmente, si bien el curso de los ríos Mira y Patía sobre las rocas Neógenas marinas expuestas en la Bahía de Tumaco pudieron haber sido otro origen de los retrabajados (Borrero et al., 2012; Bedoya et al., 2013), la concentración exclusiva de nanofósiles del Mioceno-Plioceno en la muestra PC-11-EGO sugieren una vez más procesos más localizados que estarían aportando materiales más antiguos (Figura 3).
CONCLUSIONES
Los análisis micropaleontológicos de 39 muestras extraídas de las Cuencas offshore de Tumaco y Chocó muestran que la asociación principal (mayor al 2%) de cocolitóforos es G. oceanica, G. muellerae, Gephyrocapsa <3 μm, E. huxleyi, C. leptoporus y H. carteri. En una menor proporción (<2%) se observaron Ceratolithus spp., C. pelagicus, F.profunda, H. princei, H. sellii, H. wallichii y Pontosphaera spp. junto con formas retrabajadas de Reticulofenestra spp, Sphenolithus spp. y Discoaster spp. del Mioceno-Plioceno. La presencia de E. huxleyi dentro de la asociación indica que la edad de estos sedimentos no debe ser más antigua que la biozona NN21 del Ioniano (Pleistoceno Medio). Las estimaciones del número de cocolitos por gramo (cc/g) revelaron que la abundancia promedio de la Cuenca Chocó (5,7x106 cc/g) es menor en comparación con la Cuenca Tumaco (1,2x107 cc/g). De acuerdo con los análisis estadísticos multivariados RDA, esta diferencia puede ser explicada principalmente por la distancia a la costa, la cual parece haber controlado la distribución y abundancia de los cocolitos encontrados.