










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
versión impresa ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.49 supl.1 Santa Marta dic. 2020 Epub 04-Sep-2021
https://doi.org/10.25268/bimc.invemar.2020.49.suplesp.1046
Artículos de Investigación
Asociación de la temperatura superficial del mar y la abundancia del Piquero Café (Sula leucogaster) según el estado de desarrollo en el Parque Nacional Natural Gorgona

1Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle. alejandro.perlaza@correounivalle.edu.co
2Instituto de Investigaciones en Ciencias del Mar y Limnología (Incimar), Universidad del Valle, Cali, Colombia. alan.giraldo@correounivalle.edu.co
3Estación Científica Henry von Prahl, Parque Nacional Natural Gorgona, Parques Nacionales Naturales de Colombia, Cali, Colombia. estacioncientificagorgona@gmail.com
4Departamento de Ciencias Naturales y Matemáticas, Pontificia Universidad Javeriana, Cali, Colombia. felipe.estela@javerianacali.edu.co
Variation in the physicochemical conditions of the sea can influence the distribution and abundance of seabirds by affecting the trophic structure of the pelagic environment. For this reason, we aimed to evaluate the association between the abundance of three developmental stages (chicks, juveniles, and adults) of the Brown Booby (Sula leucogaster etesiaca) and the sea surface temperature (SST) and its thermal anomaly (ANOM) in Gorgona Island, Colombian Pacific. These trends were assessed by performing cross-correlations and generalized linear models. Each developmental stage showed a different abundance tendence linked to SST and ANOM variation. It is suggested that these trends are due to the fact that increased sea temperature may be associated with reduced abundance and availability of prey resources. Under these conditions, adults tend to decreased the effort invested in parental care and even reduce the food supply to dependent juveniles and chicks to increase their own survival. These trends may have been associated with a breeding regime that allows fledglings to achieve independence from parents during the period of greatest availability of prey.
KEYWORDS: time delay; prey; Brown Bobby; survival; sea surface temperature
La variación de las condiciones fisicoquímicas del mar puede influir en la distribución y la abundancia de las aves marinas al afectar la estructura trófica del ambiente pelágico. Por ello se pretendió evaluar la asociación entre la abundancia de tres estados de desarrollo (pollos, juveniles y adultos) del Piquero Café (Sula leucogaster etesiaca) y la variación de la temperatura superficial del mar (TSM) y su anomalía térmica (ANOM) en isla Gorgona, Pacífico colombiano. Estas tendencias fueron evaluadas por medio de correlaciones cruzadas y modelos lineales generalizados. En cada estado de desarrollo la abundancia presentó una tendencia diferente ante la variación de la TSM o ANOM. Se sugiere que dichas tendencias se deben a que el incremento de la temperatura del mar podría estar asociado con una menor abundancia y disponibilidad de recursos presa. Bajo estas condiciones, los adultos tienden a disminuir el esfuerzo invertido en el cuidado parental e, incluso, reducen el suministro de alimento a juveniles dependientes y pollos para aumentar su propia supervivencia. Es posible que estas tendencias hayan estado asociadas con un régimen reproductivo que les permita a los volantones alcanzar la independencia de los padres durante el periodo con mayor disponibilidad de alimento.
PALABRAS CLAVE: desfase temporal; presas; piquero café; supervivencia; temperatura superficial del mar
INTRODUCCIÓN
Los esfuerzos continuos de monitoreo de aves marinas permiten, entre otras cosas, relacionar tendencias poblacionales con la variación en las condiciones oceanográficas (Gaston et al., 2009; Grémillet y Boulinier, 2009; Humphries, 2015). Por ejemplo, ha sido establecido que los cambios en las condiciones de la temperatura superficial del mar (TSM) pueden afectar la reproducción y supervivencia de las aves marinas, al cambiar la estructura trófica del ambiente pelágico por la alteración en la disponibilidad y calidad alimenticia de sus presas (Schreiber, 2002; Frederiksen et al., 2006; Tompkins et al., 2017; Champagnon et al., 2018). Sin embargo, la respuesta a los cambios climáticos parece estar relacionada con la localidad, especie, sexo y estado de desarrollo (Anderson, 1989; Maness y Anderson, 2013; Champagnon et al., 2018). Por ejemplo, la supervivencia de los juveniles del Piquero de Patas Azules (Sula nebouxii) se reduce durante la fase cálida del ENSO (El Niño-Oscilación del Sur) (Oro et al., 2010), mientras que el número de individuos inmaduros del Albatros de Ceja Negra (Thalassarche melanophris) se incrementa al aumentar la TSM (Pardo et al., 2017).
En el Parque Nacional Natural Gorgona (PNNG) se ha estado desarrollando el programa de monitoreo de aves marinas más longevo de Colombia, iniciado en 2002 (Estela et al., 2010). Durante este monitoreo se ha registrado continuamente información de presencia, abundancia y estado reproductivo sobre cuatro especies de aves marinas recurrentemente avistadas en el PNNG (suliformes y pelecaniformes) (Cadena-López y Naranjo, 2010; Perlaza-Gamboa et al., 2020). Entre ellas se encuentra el Piquero Café (Sula leucogaster etesiaca), subespecie endémica del Pacífico tropical húmedo americano (Ospina-Álvarez, 2004), residente en la isla Gorgona, donde se ubica la mayor población con una colonia de aproximadamente 300 individuos y 100 parejas reproductivas (Cadena-López y Naranjo, 2010; Estela et al., 2016). Esta subespecie es considerada como en amenaza en el país (Renjifo et al., 2016).
Considerando que la variabilidad de las condiciones oceanográficas asociadas al PNNG son influenciadas por el ENSO (Blanco, 2009) y que este ha sido identificado como un factor relevante para la supervivencia y los aspectos reproductivos de otras especies de piqueros en el Pacífico Oriental Tropical (Anderson, 1989; Ribic et al., 1992; Mauck y Grubb, 1995; Ancona et al., 2012) y en isla Gorgona (Perlaza-Gamboa et al., 2020), en esta investigación se evalúa la dinámica temporal de la colonia del Piquero Café en esta localidad con el propósito de responder la siguiente pregunta de investigación: ¿afecta la temperatura superficial del mar la variación temporal de la abundancia de los diferentes estados de desarrollo del Piquero Café (Sula leucogaster etesiaca) en isla Gorgona? Para ello, se parte del análisis de los registros de la abundancia de adultos, juveniles y pollos de esta especie realizados durante los monitoreos de aves marinas en el PNNG desde el 2002 hasta el 2018, así como de las condiciones de temperatura superficial del mar (TSM) y anomalía térmica (ANOM) en esta localidad.
ÁREA DE ESTUDIO
Dentro del área de conservación delimitada para el Parque Nacional Natural Gorgona (PNNG), se encuentran las islas Gorgona y Gorgonilla (2˚ 55′ 45″-3˚ 00′ 55″ N y 78˚ 09′ 00″-78˚ 14′ 30″ W), ubicadas aproximadamente a 30 km del continente (Diaz et al., 2001; Giraldo et al., 2014a). En el extremo norte de Gorgona y alrededor de Gorgonilla se encuentran una serie de islotes y promontorios rocosos emergidos, en los cuales el Piquero Café construye sus nidos (Ospina-Álvarez, 2004; Cadena-López y Naranjo, 2010). Estos islotes presentan formas e inclinaciones variables. Los piqueros son capaces de anidar en terrenos con pendientes menores a 10° y hasta de 90° (Ospina-Álvarez, 2004). Los islotes pueden presentar alguna cobertura vegetal, principalmente de herbáceas, aunque los sitios de anidación pueden establecerse en partes del islote desprovistas de vegetación, a una altura promedio de 8,9 m.s.n.m. (Ospina-Álvarez, 2004).
La marea es de tipo semidiurno, con un rango de altura entre 4 y 5 m (Díaz et al., 2001; IDEAM, 2019). Se registran niveles de precipitación mayores a 6000 mm/año, con mayores registros entre mayo y noviembre y menores desde diciembre hasta abril (Blanco, 2009). Con relación a las características de la columna de agua del ambiente pelágico, se ha reportado un periodo frío de alta salinidad entre enero y abril, durante el cual la termoclina se ubica a 7 m de profundidad, y un periodo cálido de baja salinidad entre mayo y diciembre, durante el cual la termoclina se ubica a más de 40 m de profundidad (Giraldo et al., 2008).
MATERIALES Y MÉTODOS
La información utilizada para esta investigación correspondió a los registros mensuales de abundancia de Piquero Café realizados por funcionarios entrenados del PNNG, entre 2002 y 2018, mediante censos visuales a lo largo de un trayecto predeterminado en el marco del programa de monitoreo de aves marinas de esta área protegida (Figura 1). Estos registros se realizan durante dos días consecutivos de la primera semana de cada mes. Información detallada sobre la metodología asociada a este programa de monitoreo puede ser obtenida en Payán (2016) y Perlaza-Gamboa et al. (2020).
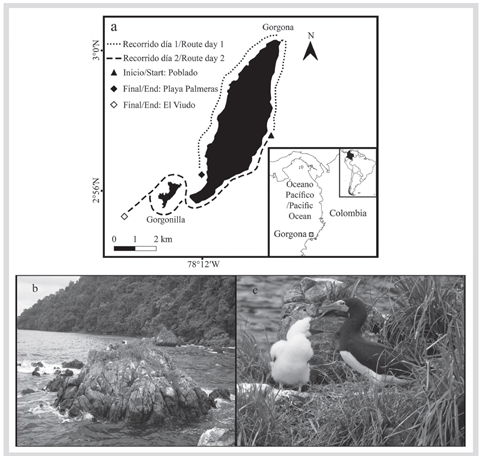
Figura 1 Generalidades del monitoreo de aves marinas del PNN Gorgona, Pacifico colombiano. A) Recorrido realizado durante el monitoreo. B) Ejemplo de islote en el sector norte de la Isla. C) Hembra y polluelo de Piquero Café (Sula leucogaster etesiaca). Fotografías: Felipe A. Estela.
Cada individuo observado fue asignado a uno de tres estados de desarrollo definidos de acuerdo con las características de su plumaje: los pollos se identificaron por su plumón blanco característico, los juveniles por presentar una coloración general café grisácea con motas café oscuro y los adultos por el contraste entre la coloración café oscura de la cabeza, cuello y dorso con el blanco del pecho bajo y vientre (Hilty y Brown, 2001; Ospina-Álvarez, 2004). Se estandarizaron las abundancias dividiendo el número de individuos observados por la distancia total recorrida en cada uno de los muestreos. Para dar uniformidad a los datos de abundancia, solo se consideraron para el análisis numérico los eventos de muestreo en los que se llevaron a cabo las dos jornadas de observación. Se utilizaron las abundancias de pollos registradas desde 2002, ya que su identificación fue siempre posible. Sin embargo, durante los primeros años de muestreo no se discernía entre individuos adultos y juveniles, por lo que para analizar estos dos estados de desarrollo se emplearon los datos obtenidos a partir de 2011, año desde el cual se empezó a diferenciar entre estas dos categorías. Los datos de individuos con estado de desarrollo no identificado no fueron utilizados. Para el análisis descriptivo de estos datos se utilizó la mediana como parámetro de tendencia central y la máxima y mínima abundancia como parámetros de dispersión para disminuir la influencia de datos atípicos y por tratarse de datos discretos.
Los datos de la TSM de isla Gorgona fueron obtenidos de las imágenes mensuales satelitales registradas por el sensor MODIS-Aqua del Ocean Biology Processing Group (OBPG), en la estación CPC16 de la malla de muestreo ERFEN (3° 0′ 0″ N-78° 0′ 0″ W) (OBPG, 2015). La anomalía térmica (ANOM) fue calculada al substraer el promedio histórico mensual del periodo 2002-2018 de la serie temporal mensual de la TSM.
Para establecer el desfase temporal de la posible asociación entre la temperatura y abundancia de pollos, adultos y juveniles, se implementó una correlación cruzada entre los registros de la abundancia mensual con la TSM y la ANOM de isla Gorgona. El mayor desfase (Lag) permitido para los pollos fue de cuatro meses, puesto que es el lapso que incluye el periodo de incubación más el tiempo que tardan los pollos en crecer hasta ser visibles en el nido (Nelson, 1978; Ospina-Álvarez, 2004). Para los juveniles el mayor desfase permitido fue de siete meses, que corresponde, en promedio para la especie, al tiempo comprendido desde la incubación hasta la alimentación de volantones por parte de los padres (Nelson, 1978; Ospina-Álvarez, 2004). Finalmente, para los adultos el máximo desfase permitido fue de 12 meses, ya que este lapso abarca completamente la variación de su abundancia. La fuerza de la correlación fue medida con la función de autocorrelación (ACF) que, para este estudio, adquiere un valor de -1 en la mayor correlación negativa y de 1 para la mayor correlación positiva.
Se desarrolló un modelo lineal generalizado (MLG) para cada estado de desarrollo de manera independiente con la finalidad de explicar la posible relación entre la abundancia y la TSM y la ANOM, que presentaron correlaciones significativas teniendo en cuenta el desfase temporal encontrado en la correlación cruzada. Se utilizó la distribución quasipoisson porque se evaluaron datos de conteo y no se presentó sobredispersión. Los análisis y las figuras fueron hechos en el programa R (R Core Team, 2018). Se utilizó el paquete ggplot2 para las figuras (Wickham, 2016), astsa para la correlación cruzada (Stoffer, 2017) y MASS para los modelos (Venables y Ripley, 2002), además de otras funciones para organizar los datos (Wickham, 2007; Fox y Weisberg, 2011; Trapletti y Hornik, 2018).
RESULTADOS
Durante el periodo de estudio fueron hechos 184 registros mensuales de pollos de Piquero Café y 92 registros mensuales de adultos y juveniles. La abundancia de adultos presentó la mayor variación, con una mediana mensual de 5,70 ind/km y un máximo y mínimo de 11,02 y 2,00 ind/km, respectivamente. Los pollos presentaron una mediana mensual de 0,22 ind/km (mínimo = 0 ind/km y máximo = 2,62 ind/km) y los juveniles de 0,42 ind/km (mínimo = 0 ind/km y máximo = 1,54 ind/km), con mayores registros, en ambos casos, durante el último semestre (Figura 2).
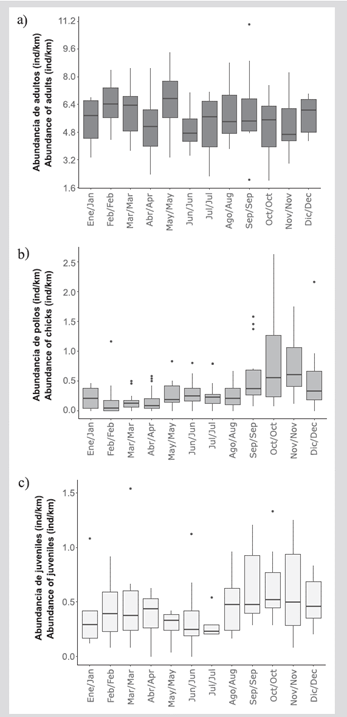
Figura 2 Variación mensual de la abundancia de A) adultos, B) pollos y C) juveniles de Piquero Café (Sula leucogaster etesiaca) en isla Gorgona, Pacífico colombiano. La caja representa la mediana y los percentiles de 25% y 75%. Las barras representan el valor máximo y mínimo no atípico y los puntos corresponden a datos atípicos.
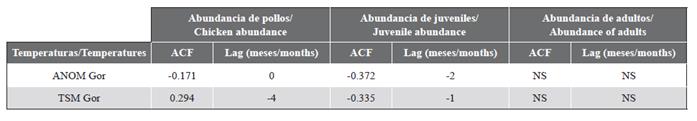
La temperatura superficial del mar de isla Gorgona presentó una mediana igual a 27,26 °C, con registros entre 25,47 °C y 28,05 °C. Los adultos no presentaron ninguna correlación significativa con la TSM ni con la ANOM. En contraste, la abundancia de juveniles presentó una correlación significativa con la ANOM y la TSM, con un desfase temporal de dos y un mes, respectivamente. Esto sugiere que el número de juveniles está asociado con la ANOM registrada dos meses antes y con la TSM del mes anterior. Finalmente, la abundancia de pollos estuvo correlacionada con la ANOM sin ningún desfase temporal y con la TSM registrada cuatro meses antes (Tabla 1).
Tabla 1 Correlación cruzada entre la abundancia mensual de pollos, juveniles y adultos de Piquero Café (Sula leucogaster etesiaca) con la temperatura superficial del mar (TSM) de isla Gorgona (Gor) y su anomalía térmica (ANOM). ACF = fuerza de correlación. ANOM = anomalía térmica. Lag = desfase temporal de la correlación medido en meses. NS = correlación no significativa.
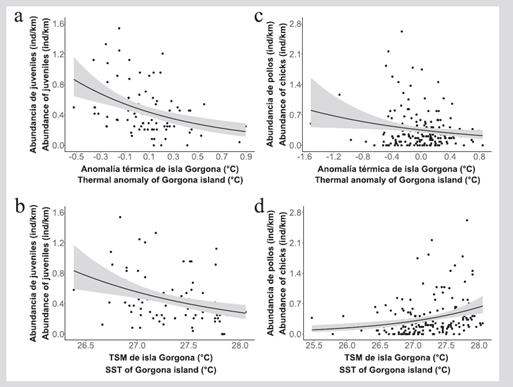
De acuerdo con el modelo lineal generalizado (MLG) desarrollado entre la abundancia de juveniles con la ANOM (coeficiente = 0,33; P < 0,001) y la TSM (coeficiente = 0,52; P < 0,001), considerando el respectivo desfase (Lag), el número de juveniles tiende a disminuir con el incremento de ambas variables térmicas (Figura 3A y 3B). Por otro lado, la abundancia de pollos tiende a disminuir con el aumento de la ANOM (coeficiente = 0,58; P = 0,017) (Figura 3C), pero incrementa al aumentar la TSM (coeficiente = 2,08; P < 0,001) (Figura 3D).
DISCUSIÓN
La abundancia de pollos y juveniles de Piquero Café asociadas a isla Gorgona fue mayor en los últimos meses del año, probablemente como consecuencia de la concentración del esfuerzo reproductivo entre julio y septiembre, condición que fue reportada por Cadena-López y Naranjo (2010). Sin embargo, esta especie exhibe actividad reproductiva durante todo el año, con una baja variación intraanual del tamaño de la población (Ospina-Álvarez, 2004; Cadena-López y Naranjo, 2010; Perlaza-Gamboa et al., 2020). En este régimen reproductivo la mayoría de las posturas ocurre entre junio y agosto, con un periodo de incubación entre 40 y 44 días, seguido de un lapso entre uno y tres meses en que los pollos crecen hasta ser visibles y adquieren el plumón blanco característico, que al final de este periodo empieza a mudar por el plumaje café (Nelson, 1978). Una vez estos individuos realizan el primer vuelo, aproximadamente a los 96 días de edad (Nelson, 1978; Ospina-Álvarez, 2004), son alimentados por los padres durante las siguientes tres a ocho semanas, aunque este periodo puede extenderse dependiendo de la disponibilidad de presas (Schreiber y Norton, 2002). Este sistema favorece que los individuos adquieran la independencia de los padres durante los primeros meses del año, tiempo en el cual se ha registrado mayor productividad en el ambiente pelágico aledaño a isla Gorgona (Giraldo et al., 2014b).
Esta estrategia reproductiva ha sido registrada para otras especies de aves marinas que comienzan a reproducirse cuando hay menor productividad en el ecosistema y sus presas están menos disponibles, pero permite que las crías alcancen la independencia de los padres cuando se presenta la mayor disponibilidad de presas (Passuni et al., 2016, 2018). Por lo general, esta estrategia es llevada a cabo, principalmente, por las aves cuyo alimento está ampliamente distribuido, como ocurre con las aves marinas (Nelson, 1978; Furness y Monaghan, 1987; Vilchis et al., 2006). Además, cabe mencionar que el desfase de cuatro meses en la correlación positiva entre la abundancia de pollos y la TSM sugiere que la mayor abundancia observada en octubre y noviembre se ve influenciada por las temperaturas registradas a mitad de año, tiempo en el que ocurre la transición entre el periodo de mayor a menor productividad en el Pacífico Oriental Tropical (Pennington et al., 2006) y probablemente en los alrededores de isla Gorgona (Giraldo et al., 2014b). Este patrón es similar al observado en el Pacífico tropical mexicano, donde el periodo reproductivo estuvo asociado principalmente con la época más cálida y menos productiva del año, pero coincide con el incremento de la disponibilidad de presas migratorias de aguas cálidas (Hernández-Vázquez et al., 2017). Por estas razones, para comprender el régimen reproductivo del Piquero Café en isla Gorgona, resulta necesario profundizar en el conocimiento sobre la variación poblacional intra anual de sus recursos presa, así como determinar el periodo en el que la mayoría de las crías adquiere la independencia de los padres.
Es probable que la menor productividad asociada con aguas más cálidas y mayor precipitación entre mayo y diciembre (Diaz et al., 2001; Giraldo et al., 2014b) provoque mayor pérdida de pollos, como ha sido registrado para otras aves marinas en el Pacífico Oriental (Jaksic y Fariña, 2010). Esto podría desencadenar con más frecuencia segundas nidadas por parte de las parejas reproductivas (Nelson, 1978) y resultar así en un mayor número de pollos observados en los últimos meses. Por otro lado, la correlación negativa encontrada entre la abundancia de pollos y la anomalía térmica sugiere que estos individuos son afectados por los cambios de la temperatura del mar, posiblemente por la disminución en el alimento brindado por sus padres. Por ejemplo, para algunas aves marinas se ha asociado el aumento de la temperatura del mar y la disminución en el éxito reproductivo con una alteración en la productividad del mar y la disponibilidad de presas (Ancona et al., 2011; Furness, 2016). La menor disponibilidad de recursos presa relacionada con aguas más cálidas puede explicar la disminución de la abundancia de juveniles del Piquero Café cuando aumenta la TSM y ANOM en el ambiente pelágico de isla Gorgona.
Durante condiciones adversas, como la baja disponibilidad de presas, algunas especies longevas como el Piquero Café (Hennicke et al., 2012) generalmente tienden a minimizar los costos individuales de la reproducción (Mauck y Grubb, 1995). Por ejemplo, los adultos disminuyen el esfuerzo invertido en el cuidado parental y en el suministro de alimento a los juveniles dependientes o pollos con la finalidad de incrementar su propia supervivencia, pero disminuyendo el éxito reproductivo (Maness y Anderson, 2013). Esta estrategia puede explicar por qué no se observó ninguna correlación significativa entre los adultos y la TSM o ANOM, pero sí en los pollos y juveniles. Además, la abundancia de este piquero descrita por Perlaza-Gamboa et al. (2020) podría deberse sobre todo a la disminución en el número de juveniles, posiblemente debido a que el suministro de alimento haya sido menor cuando aumentaba la TSM. No obstante, es probable que este efecto sea menor en las especies que presentan cuidado parental antes y después de que las crías se conviertan en volantones (Stienen y Brenninkmeijer, 2002; Maness y Anderson, 2013), como es el caso del Piquero Café (Ospina-Álvarez, 2004). Así y todo, estas hipótesis deben ser confirmadas por un estudio que evalúe las estrategias llevadas a cabo por los parentales durante condiciones de estrés y sus consecuencias en el desarrollo y la supervivencia de su descendencia antes y después de haber alcanzado la independencia.
CONCLUSIONES
La TSM y ANOM de isla Gorgona están asociados con las tendencias observadas en la variación poblacional de los pollos y juveniles de Piquero Café, posiblemente a través de un efecto sobre el suministro de peces pelágicos brindados por los padres, debido a que es probable que estos individuos no puedan satisfacer las necesidades de las crías, pero sí logren mantenerse vivos durante la época de escasez de alimento. Esto se evidencia en un menor número de juveniles y pollos durante los meses con mayores anomalías, lo que sugiere que la variación de la TSM y ANOM puede repercutir en la reproducción de esta población. Además, posiblemente, el régimen reproductivo garantiza que las crías alcancen la independencia de los padres cuando haya mayor disponibilidad de recursos presa.
AGRADECIMIENTOS
A Ximena Zorrilla y Héctor Chirimía del Parque Nacional Natural Gorgona (PNNG). A Julio César Herrera Carmona por su ayuda en la construcción de la serie temporal de la TSM y a Wilmar Alexander Torres por sus recomendaciones para el desarrollo de los análisis estadísticos. A WWF y a la Asociación Calidris por el apoyo en el programa de monitoreo de aves marinas desde sus inicios hasta ahora. Este trabajo de investigación fue cofinanciado por el PNNG, la Universidad del Valle y WWF-Colombia, y se realizó en el marco de un acuerdo de entendimiento entre el PNNG y el grupo de investigación en Ecología Animal de la Universidad del Valle. Publicación 008 del Instituto de Ciencias del Mar y Limnología (Incimar) de la Universidad del Valle.
REFERENCIAS
Ancona, S., S., Sánchez-Colón, C., Rodríguez, and H. ,Drummond. 2011. El Niño in the warm tropics: local sea temperature predicts breeding parameters and growth of blue-footed boobies. J. An. Ecol., 80: 799-808. https://doi.org/10.1111/j.1365-2656.2011.01821.x [ Links ]
Ancona, S., I., Calixto-Albarrán, and H., Drummond. 2012. Effect of El Niño on the diet of a specialist seabird, Sula nebouxii, in the warm eastern tropical Pacific. Mar. Ecol. Progr. Ser., 462: 261-271. https://doi.org/10.3354/meps09851 [ Links ]
Anderson, D.J. 1989. Differential responses of boobies and other seabirds in the Galapagos to the 1986-87 El Nino-Southern Oscillation event. Mar. Ecol. Progr. Ser., 52: 209-2016. [ Links ]
Blanco, J.F. 2009. The hydroclimatology of Gorgona Island: seasonal and Enso-Rel. patterns. Actual. Biol., 1(91): 111-121. [ Links ]
Cadena-López, G. y L.G., Naranjo. 2010. Distribución, abundancia y reproducción de las aves marinas residentes en el Parque Nacional Natural Gorgona, Colombia. Bol. SAO, 20: 22-32. [ Links ]
Champagnon, J., J. D. ,Lebreton, H., Drummond, and D. J. ,Anderson. 2018. Pacific decadal and El Niño oscillations shape survival of a seabird. Ecology, 99(5): 1063-1072. https://doi.org/10.1002/ecy.2179 [ Links ]
Díaz, J.M., J.H. ,Pinzón, A.M., Perdomo, L.M., Barrios y M., López-Victoria. 2001. Generalidades. 17-26. En Barrios, L. M. y M., Lopez-Victoria (Eds.), Gorgona marina: contribución al conocimiento de una isla única. Ser. Publ. Esp. Invemar, (7). [ Links ]
Estela, F.A., M., López Victoria, L.F., Castillo y L.G., Naranjo. 2010. Estado del conocimiento sobre aves marinas en Colombia después de 110 años de investigación. Bol. SAO, 20: 2-21. [ Links ]
Estela, F.A., G., Cadena y J., Lasso-Zapata. 2016. Sula leucogaster. 139-143. En Renjifo, L.M., Á. M., Amaya-Villarreal, J., Burbano-Girón y J., Velásquez-Tibatá (Eds.). Libro rojo de aves de Colombia. Volumen II: ecosistemas abiertos, secos, insulares, acuáticos continentales, marinos, tierras altas del Darién y Sierra Nevada de Santa Marta y bosques húmedos del centro, norte y oriente del país. Pont. Univ. Javeriana e Inst. Alexander von Humboldt, Bogotá. 563 p. [ Links ]
Fox, J. and S., Weisberg. 2011. An R Companion to Applied Regression [software]. Thousand Oaks: Sage. http://socserv.socsci.mcmaster.ca/jfox/Books/Companion.04/04/2019. [ Links ]
Frederiksen, M., M., Edwardst, A.J., Richardson, N.C. ,Halliday, and S., Wanless. 2006. From plankton to top predators: bottom-up control of a marine food web across four trophic levels. J. An. Ecol., 75(6): 1259-1268. https://doi.org/10.111 1l/j.1365-2656.2006.01 [ Links ]
Furness, R.W. 2016. Impacts and effects of ocean warming on seabirds. 271-288. En Laffoley, D. y J. M., Baxter (Eds.). Explaining ocean warming: causes, scale, effects and consequences. IUCN, Gland, Switzerland. [ Links ]
Furness, R.W. and P., Monaghan. 1987. Seabird ecology. Chapman & Hall, New York. [ Links ]
Gaston, A.J., D.F., Bertram, A. W., Boyne, J.W. ,Chardine, G. ,Davoren, A.W., Diamond, A., Hedd, W.A., Montevecchi, J.M., Hipfner, M.J.F. ,Lemon, M.L., Mallory, J.F., Rail, and G.J., Robertson. 2009. Changes in Canadian seabird populations and ecology since 1970 in relation to changes in oceanography and food webs. Environ. Rev., 17: 267-286. https://doi.org/10.1139/A09-013 [ Links ]
Giraldo, A., E., Rodríguez-Rubio y F. ,Zapata. 2008. Condiciones oceanográficas en isla Gorgona, Pacífico oriental tropical de Colombia. Latin Am. J. Aq. Res., 36(1): 121-128. https://doi.org/10.3856/vol36-issue1-fulltext-12 [ Links ]
Giraldo, A., M.C., Díaz-Granados y C.F., Gutiérez-Landázuri. 2014a. Isla Gorgona, enclave estratégico para los esfuerzos de conservación en el Pacífico Oriental Tropical. Rev. Biol. Trop., 62: 1-12. [ Links ]
Giraldo, A., B., Valencia, J.D., Acevedo y M., Rivera. 2014b. Fitoplancton y zooplancton en el área marina protegida de Isla Gorgona, Colombia, y su relación con variables oceanográficas en estaciones lluviosa y seca. Rev. Biol. Trop., 62: 117-132. [ Links ]
Grémillet, D. and T., Boulinier. 2009. Spatial ecology and conservation of seabirds facing global climate change: a review. Mar. Ecol. Progr. Ser., 391(2): 121-137. https://doi.org/10.3354/meps08212 [ Links ]
Hennicke, J.C., B., King, D., Drynan, L.J., Hardy, A., Stokes, and S. ,Taylor. 2012. New life-span records of the Brown Booby Sula leucogaster. Mar. Ornithol., 40(2), 125-126. [ Links ]
Hernández-Vázquez, S., E., Mellink, J.A. ,Castillo-Guerrero, R., Rodríguez-Estrella, J.Á., Hinojosa-Larios y V.H., Galván-Piña. 2017. Ecología reproductiva del bobo café (Sula leucogaster) en tres islas del Pacífico Tropical mexicano. Ornitol. Neotrop., 28: 57-66. [ Links ]
Hilty, S.L. y W.L., Brown. 2001. Guía de las aves de Colombia. Princeton, New Jersey. 1040 p. [ Links ]
Humphries, G.R.W. 2015. Estimating regions of oceanographic importance for seabirds using a-spatial data. PLoS ONE, 10(9): e0137241. https://doi.org/10.1371/journal.pone.0137241 [ Links ]
IDEAM. 2019. Pronóstico de pleamares y bajamares Costa Caribe colombiana 2020. Inst. Hidrol. Meteorol. Est. Amb., Bogotá. 135 p. [ Links ]
Jaksic, F.M. and J.M., Fariña. 2010. El Niño and the birds: a resource-based interpretation of climatic forcing in the southeastern Pacific. An. Inst. Patagonia, 38(1): 121-140. https://doi.org/10.4067/s0718-686x2010000100009 [ Links ]
Maness, T.J. and D.J., Anderson. 2013. Predictors of juvenile survival in birds. Ornithol. Monogr., 78: 1-55. https://doi.org/10.1525/om.2013.78.1.1. [ Links ]
Mauck, R.A. and T.C., Grubb. 1995. Petrel parents shunt all experimentally increased reproductive costs to their offspring. An. Behav., 49: 999-1008. https://doi.org/10.1006/anbe.1995.0129 [ Links ]
Nelson, J.B. 1978. The Sulidae, gannets and boobies. Oxford University Press, Oxford. 1012 p. [ Links ]
OBPG. 2015. MODIS aqua level 3 SST thermal IR monthly 4km daytime v2014.0. https://podaac.jpl.nasa.gov/dataset/MODIS_AQUA_L3_SST_THERMAL_MONTHLY_4KM_DAYTIME_V2014.0. 21/01/2019. [ Links ]
Oro, D., R. ,Torres, C., Rodríguez, and H., Drummond. 2010. Climatic influence on demographic parameters of a tropical seabird varies with age and sex. Ecology, 91(4): 1205-1214. [ Links ]
Ospina-Álvarez, A. 2004. Ecología reproductiva y colonialidad del piquero café Sula leucogaster (Aves: Sulidae), en el PNN Gorgona, Pacífico colombiano. Tesis Biol., Univ. Valle, Cali. 116 p. [ Links ]
Pardo, D., S. ,Jenouvrier, H., Weimerskirch, and C., Barbraud. 2017. Effect of extreme sea surface temperature events on the demography of an age-structured albatross population. Phil. Trans. Royal Soc. B. Biol. Sci., 372: 20160. https://doi.org/10.1098/rstb.2016.0143 [ Links ]
Passuni, G., C., Barbraud, A., Chaigneau, H., Demarcq, J., Ledesma, A., Bertrand, R. ,Castillo, A., Perea, J. ,Mori, V. A., Viblanc, J., Torres-Maita, and S., Bertrand. 2016. Seasonality in marine ecosystems: Peruvian seabirds, anchovy, and oceanographic conditions. Ecology, 97(1), 182-193. https://doi.org/10.1890/14-1134.1 [ Links ]
Passuni, G., C., Barbraud, A., Chaigneau, A., Bertrand, R., Oliveros-Ramos, J., Ledesma, R., Castillo, M., Bouchon, and S., Bertrand. 2018. Long-term changes in the breeding seasonality of Peruvian seabirds and regime shifts in the Northern Humboldt Current System. Mar. Ecol. Progr. Ser., 597: 231-242. https://doi.org/10.3354/meps12590 [ Links ]
Payan, L.F. 2016. Informe de monitoreo de aves marinas, Parque Nacional Natural Gorgona, Febrero - Diciembre 2016. Cali. [ Links ]
Pennington, J.T., K.L. ,Mahoney, V.S., Kuwahara, D.D. ,Kolber, R., Calienes, and F.P. ,Chavez. 2006. Primary production in the eastern tropical Pacific: a review. Progr. Oceanogr., 69(2-4): 285-317. https://doi.org/10.1016/j.pocean.2006.03.012 [ Links ]
Perlaza-Gamboa, A., A., Giraldo, L.F. ,Payán y F.A., Estela. 2020. Variación poblacional de tres especies de piqueros (Suliformes: Sulidae) en isla Gorgona, Pacífico colombiano, según la temperatura del mar. Rev. Biol. Trop., 68(2): 704-713. [ Links ]
R Core Team. 2018. R: a language and environment for statistical computing (Version 3.5.2). R Foundation for Statistical Computing, Vienna. https://www.r-project.org.04/04/2019. [ Links ]
Renjifo, L.M., Á.M., Amaya-Villarreal, J., Burbano-Girón y J., Velásquez-Tibatá (Eds). 2016. Libro rojo de aves de Colombia. Volumen II: Ecosistemas abiertos, secos, insulares, acuáticos continentales, marinos, tierras altas del Darién y Sierra Nevada de Santa Marta y bosques húmedos del centro, norte y oriente del país. Bogotá, D.C.: Editorial Pontificia Universidad Javeriana e Instituto Alexander von Humboldt. [ Links ]
Ribic, C.A., D.G., Ainley, and L.B., Spear. 1992. Effects of El Nino and La Nina on seabird assemblages in the equatorial Pacific. Mar. Ecol. Progr. Ser., 80: 109-124. https://doi.org/10.3354/meps080109 [ Links ]
Schreiber, E.A. 2002. Climate and weather effects on seabirds: 179-216. En Schreiber, E.A. y J., Burger (Eds.). Biology of marine birds. CRC Press, Boca Ratón, USA. [ Links ]
Schreiber, E.A. and R.L., Norton. 2002. Brown Booby (Sula leucogaster), version 2.0. En Poole, A. y F.B., Gill (Eds.). The birds of North America online. Cornell Lab of Ornithology, Ithaca, USA. https://doi.org/10.2173/bna.649 [ Links ]
Stienen, E.W.M. and A., Brenninkmeijer. 2002. Variation in growth in Sandwich Tern chicks Sterna sandvicensis and the consequences for pre- and post-fledging mortality. Ibis, 144: 567-576. https://doi.org/10.1046/j.1474-919X.2002.00086.x [ Links ]
Stoffer, D. 2017. astsa: Applied Statistical Time Series Analysis [software]. https://cran.r-project.org/package=astsa. 04/04/2019. [ Links ]
Tompkins, E.M., H.M., Townsend, and D.J., Anderson. 2017. Decadal-scale variation in diet forecasts persistently poor breeding under ocean warming in a tropical seabird. PLoS ONE, 12(8): 1-24. https://doi.org/10.1371/journal.pone.0182545 [ Links ]
Trapletti, A. and K., Hornik. 2018. tseries: Time Series Analysis and Computational Finance [software]. https://cran.r-project.org/package=tseries. 04/04/2019. [ Links ]
Venables, W.N. and B.D., Ripley. 2002. Modern applied statistics with S [software]. Springer, New York, USA. http://www.stats.ox.ac.uk/pub/MASS4.04/04/2019. [ Links ]
Vilchis, L.I., L.T., Balance, and P.C., Fiedler. 2006. Pelagic habitat of seabirds in the Eastern Tropical Pacific: effects of foraging ecology on habitat selection. Mar. Ecol. Progr. Ser., 315: 279-292. https://doi.org/10.3354/meps315279 [ Links ]
Wickham, H. 2007. Reshaping data with the reshape package [software]. J. Statist. Softw., 21(12): 1-20. http://www.jstatsoft.org/v21/i12/. 04/04/2019. [ Links ]
Wickham, H. 2016. ggplot2: Elegant graphics for data analysis [software]. Springer-Verlag, New York. http://ggplot2.org. 04/04/2019. [ Links ]
Recibido: 23 de Julio de 2020; Aprobado: 14 de Octubre de 2020
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
