










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.49 supl.1 Santa Marta Dec. 2020 Epub Sep 03, 2021
https://doi.org/10.25268/bimc.invemar.2020.49.suplesp.1090
Artículos de Investigación
Peces criptobentónicos en el Parque Nacional Natural Uramba Bahía Málaga (Colombia), Pacífico Oriental Tropical
1
 http://orcid.org/0000-0002-7849-5205
http://orcid.org/0000-0002-7849-5205
2
http://orcid.org/0000-0002-4156-0755
1Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle, Cali, Colombia. jaime.cantera@correounivalle.edu.co
2Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz, México. dmedinac1900@alumno.ipn.mx
3Departamento de Ciencias Naturales y Matemáticas, Facultad de Ingeniería y Ciencias, Pontificia Universidad Javeriana, Cali, Colombia
4Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle, Cali, Colombia. jaime.cantera@correounivalle.edu.co
Cryptobenthic fishes contribute significantly to the trophic dynamics of coastal ecosystems. This study aimed at determining the composition of this assemblage and testing if its structure differs temporally and spatially in the Uramba National Natural Park Bahía Málaga (Tropical Eastern Pacific). Fish assemblages were collected using artificial traps in three zones, between June 2008 and August 2009. Twenty-six fish species belonging 11 families were recorded. Gobiidae and Labrisomidae were the most representative families (species: 34.6 % and individuals: 66 %). Species richness and abundance increased in a gradient from the interior to the exterior of the park. Using multivariate statistic techniques, a spatial pattern in assemblage structure was identified with inner zone being significantly different from the mid and outer zones. Cerdale ionthans, Gobiosoma seminudum, Starksia fulva, and Gobulus hancocki were mainly responsible for this differentiation (> 59 % contribution). The use of artificial habitats to sample cryptobenthic fish proved to be an efficient way to identify an overlooked part ichthyofauna using traditional sampling techniques and was also useful to compare the spatial variability of these assemblages. Comparisons with cryptobenthic fish assemblages in other biogeographic regions indicate that, due to their low species richness in the Tropical Eastern Pacific, these assemblages in both Colombia and Mexico show low functional redundancy and considerable differences in their predominant trophic modes.
KEYWORDS: cryptic ichthyofauna; Gobiidae; Labrisomidae; estuarine ecology; rocky reefs.
Los peces criptobentónicos contribuyen de manera significativa a la dinámica trófica de los ecosistemas costeros. Este estudio tuvo como objetivo determinar la composición de esta comunidad y comprobar si su estructura difiere temporal y espacialmente en el Parque Nacional Natural Uramba Bahía Málaga (Pacífico Oriental Tropical). Los peces se recolectaron utilizando sustratos artificiales en tres zonas, entre junio de 2008 y agosto de 2009. Se registraron 26 especies, pertenecientes a 11 familias. Gobiidae y Labrisomidae fueron las familias más representativas (especies: 34,6 % e individuos: 66 %). La riqueza y abundancia aumentó en un gradiente del interior hacia el exterior del parque. Utilizando estadística multivariada, se identificó un patrón espacial en la estructura de la comunidad, con localidades internas significativamente diferentes de las medias y externas. Cerdale ionthas, Gobiosoma seminudum, Starksia fulva y Gobulus hancocki fueron responsables de esta diferenciación (contribución > 59 %). El uso de sustratos artificiales para tomar muestras de peces criptobentónicos demostró ser una forma eficiente de identificar una parte de la ictiofauna que no había sido identificada mediante técnicas de muestreo tradicionales y también fue útil para comparar la variabilidad espacial de esta comunidad. La comparación con comunidades de peces criptobentónicos en otras regiones biogeográficas indica que, debido a la baja riqueza de especies en el Pacífico Oriental Tropical, estas comunidades tanto en Colombia como en México muestran baja redundancia funcional y considerables diferencias en sus preferencias tróficas.
PALABRAS CLAVE: ictiofauna críptica; Gobiidae; Labrisomidae; ecología estuarina; arrecifes rocosos.
INTRODUCCIÓN
Los peces criptobentónicos pueden ser definidos como “peces adultos de usualmente < 5 cm que son visualmente y/o de comportamiento críptico, los cuales tienen una fuerte relación con el bentos” (Depczynski y Bellwood, 2003: 183). En fondos duros, como en los arrecifes de coral, estos pueden llegar a ser ~50 % de la abundancia de los peces, ~40 % del número de especies de peces y ~35 % del total de la biomasa íctica (Ackerman y Bellwood, 2000, 2002). La gran abundancia de larvas de peces criptobentónicos y el análisis del contenido estomacal de depredadores arrecifales sugieren que probablemente estos desempeñan un papel importante en la dinámica trófica de los arrecifes coralinos, al participar en el reciclaje de la biomasa derivada de la producción primaria a través de vías detríticas (Depczynski y Bellwood, 2003; Brandl et al., 2019). Por otra parte, análisis de isótopos estables (δ15N) en zonas estuarinas tropicales señalan un nivel trófico medio y alto para este tipo de organismos, lo cual indicaría que se comportan como depredadores de microfauna bentónica (Medina-Contreras et al., 2018).
Las técnicas de muestreo utilizadas comúnmente, como los censos visuales, no permiten obtener información ecológica sobre las comunidades de peces criptobentónicos (Willis, 2001) debido al sesgo que se presenta hacia individuos con longitud total de > 5 cm. Esta técnica suele subestimar la contribución de los peces criptobentónicos, por lo que se han realizado ajustes a los censos visuales para estudiar este grupo de organismos (Beldade y Gonçalves, 2007). En aguas turbias como en las zonas estuarinas cercanas a bosques de manglar, donde la visibilidad es reducida (< 1 m), los censos visuales son poco prácticos. Por lo tanto, para muestrear esta comunidad en estos ambientes se han usado sustancias anestésicas e ictiocidas (v. g. Barletta et al., 2002). Estas sustancias tienen restricciones en su uso debido a su impacto ambiental; sin embargo, pueden brindar información precisa sobre inventarios de biodiversidad a escalas pequeñas de muestreo (Robertson y Smith-Vaniz, 2008). En el caso de áreas marinas protegidas, el uso de estas sustancias es restringido debido a que podrían afectar otros organismos que no hacen parte del grupo de estudio, así como los ecosistemas muestreados. Por esta razón, es necesario contar con métodos de muestreo alternativos, los cuales incluyen el uso de unidades estandarizadas que imitan las condiciones del sustrato (Zimmerman y Martin, 2004; Ransome et al., 2017). Estas unidades de muestreo se ubican en hábitats específicos y se recuperan después de un cierto período, recolectando peces y especies de invertebrados crípticos de una manera efectiva. Estos dispositivos también se pueden utilizar para investigar eventos de asentamiento y reclutamiento (Robertson et al., 1988; Glynn, 2006; Valles et al., 2006; Carassou et al., 2009; Mellin y Pontón, 2009).
Hasta el momento no existen estudios que hayan documentado la abundancia relativa de la comunidad de peces crípticos en los sistemas estuarinos del Pacífico Oriental Tropical (POT) y particularmente en la costa Pacífica de Colombia. Para la mayoría de las especies que conforman esta comunidad, la única información disponible está restringida a su descripción original, registros únicos y listas de especies de áreas específicas. Los estudios que abordan la ecología de los peces crípticos en el POT se han realizado en charcos intermareales y arrecifes coralinos en México y Colombia (Castellanos-Galindo et al., 2005, 2014; Balart et al., 2006; González-Cabello y Bellwood, 2009; Alzate et al., 2014). Los peces criptobentónicos del POT poseen una baja riqueza de especies comparada con otras áreas, lo que produce baja redundancia funcional entre las especies que componen estas comunidades (González-Cabello y Bellwood, 2009). Lo anterior hace referencia a que pocas especies desempeñan una misma función en el ecosistema (v. g. hábitos alimentarios) (Loreau, 2004), por lo que esta función es vulnerable a perturbaciones. Dichas características hacen que esta comunidad de organismos sea vulnerable tanto a impactos antropogénicos como ambientales.
En el presente trabajo se documenta por primera vez la comunidad de peces criptobentónicos en el área marina protegida Parque Nacional Natural Uramba Bahía Málaga (PNN Uramba). También se evalúa si la estructura de esta comunidad difiere temporal y espacialmente en este lugar.
ÁREA DE ESTUDIO
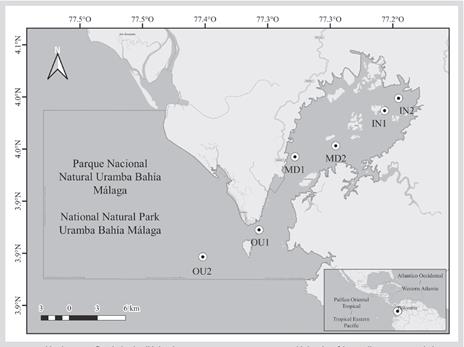
El PNN Uramba (3,953169 N-77,326456 W; Figura 1) es un área marina protegida (AMP) ubicada en la costa central del Pacífico colombiano. Esta bahía se formó durante eventos tectónicos en el Mioceno tardío-Holoceno (Martínez y López-Ramos, 2011). Tiene una extensión de 470,94 km2. Predominan condiciones estuarinas y se presentan profundidades promedio de 12-15 m. Además, alberga diversidad de hábitats que incluyen manglares, costas rocosas, planos lodosos, playas arenosas y fondos tanto blandos como duros (Cantera-Kintz et al., 2013). La precipitación anual es una de las mayores entre las áreas costeras del hemisferio occidental, con un promedio de 7236 ± 2033 mm entre 1969 y 2015 (estación n.° 5407003-IDEAM), con un periodo de menores precipitaciones entre enero y abril (x̃ = 422 mm/mes) y de mayores precipitaciones entre mayo y diciembre (x̃ = 746 mm/mes) (Castellanos-Galindo y Krumme, 2013). La temperatura superficial del mar es estable durante el año (27-30 °C), mientras que los valores de salinidad varían entre 0 y 30 dependiendo de las corrientes mareales y la precipitación (Cantera-Kintz et al., 1998). Esta área se caracteriza por presentar un régimen macromareal semidiurno (ca. 4 m de rango mareal).
MATERIALES Y MÉTODOS
Muestreo
Se utilizaron sustratos artificiales duros (SAD) como unidad de muestreo, los cuales contenían ladrillos y fragmentos de conchas recogidas en el área (concheros). Estos materiales fueron depositados en canastas plásticas (35 × 20 × 18 cm) y cubiertas con una malla con ojo de 2 mm (Figura 2). Se escogieron seis localidades a lo largo de la bahía de la siguiente manera: dos en la zona interna (La Plata: IN1; Coquero: IN2), dos en la media (Curichichi: MD1; Los Negros: MD2) y dos en la externa (Islote Morro Chiquito: OU1; Los Negritos: OU2) (Figura 1). El sustrato predominante de estas localidades es rocoso y está rodeado por fondos areno-fangosos. La profundidad a la cual estuvieron los SAD fue entre 3-7 m durante marea baja.

Figura 2 Sustratos artificiales duros (SAD) usados para la colecta de peces criptobentónicos en las seis localidades del PNN Uramba. A) SAD expuesto recientemente y B) SAD antes de ser recuperado, después de cinco meses.
El muestreo se llevó a cabo entre junio de 2008 y agosto de 2009, periodo en el cual se realizaron siete salidas de campo aproximadamente cada dos meses. En la primera salida, tres SAD fueron ubicados en cada una de las localidades (3 SAD × 6 localidades = 18 SAD). En las siguientes salidas un número similar de SAD fue ubicado en cada localidad. Los SAD fueron recuperados después de la tercera salida de campo, cuando fue extraído el primer juego de 18 SAD. En las siguientes salidas, la recuperación y ubicación de SAD continuó. En promedio, los SAD estuvieron sumergidos por cinco meses (x̃ ± d.e. = 145 ± 19,4 días). En total se expusieron y recuperaron 90 SAD durante todo el muestreo. Al ser recuperado, cada SAD fue levantado del fondo cuidadosamente y puesto al interior de canastas plásticas de mayor tamaño recubiertas con una malla con un ojo de menor tamaño mientras aún se encontraban en el agua. Después fueron llevadas a la superficie lentamente con ayuda de una bolsa recuperadora. Este procedimiento se realizó con el fin de evitar que algunos especímenes escaparan durante el proceso.
Después de ser extraídos del agua, los SAD se dispusieron en bandejas plásticas con agua de mar. Una vez en tierra, fueron desarmados cuidadosamente en busca de peces e invertebrados, raspando a mano el sustrato y tamizando los sedimentos que se acumularon en cada unidad. Todos los peces fueron identificados al menor nivel taxonómico posible (siguiendo a Robertson y Allen, 2008) y se midió la longitud total (precisión = 0,01 mm) y el peso (precisión = 0,001 g). Algunos peces fueron preservados en formalina al 10 % y transportados para ser depositados en la Colección de Referencia de Ictiología de la Universidad del Valle (CIRUV). Los datos de abundancia de los tres SAD para cada localidad fueron agrupados debido al bajo número de peces recolectados en cada una de las unidades experimentales. La densidad de peces se calculó al considerar el volumen de cada SAD (35 × 20 × 18 cm = 0,0126 m3 o 0,07 m2). Se obtuvo la información sobre los hábitos alimentarios (HA) para cada especie registrada en Robertson y Allen (2008) y Froese y Pauly (2009).
Análisis de datos
Se utilizó el escalamiento multidimensional no métrico (nMDS) para explorar si existían patrones de agrupación entre localidades. Un análisis multivariado permutacional (PERMANOVA) fue usado para probar estadísticamente si la comunidad de peces criptobentónicos difería espacial (entre zonas) y temporalmente (entre meses). Además, se utilizó un análisis de similitud de porcentajes (SIMPER) para identificar las especies responsables de las diferencias entre zonas y meses cuando se encontraron diferencias significativas en el análisis PERMANOVA. Todos los análisis fueron realizados después de la transformación de los datos de abundancia a la raíz cuadrada (se excluyó un mes de muestreo para la localidad IN1 debido a la ausencia de peces). En todos los casos se usó el índice de similaridad de Bray-Curtis. Los análisis fueron realizados con el paquete vegan (Oksanen et al., 2019) del programa estadístico R (R Core Team, 2019).
RESULTADOS
Un total de 197 individuos pertenecientes a 26 especies y 11 familias de peces fueron recolectados durante todo el periodo de muestreo. De las especies encontradas, 50 % corresponden a nuevos registros para el PNN Uramba (Figura 3, Tabla 1). Gobiidae, Labrisomidae y Gobiesocidae dominaron la comunidad en número de especies. Gobiidae (43,7 % del total de individuos) fue la familia más representativa en cuanto a abundancia, seguida por Labrisomidae (22,3 %) y Muraenidae (11,7 %). Las familias con menor representatividad en las muestras (menos de tres especies y pocos individuos) fueron Balistidae, Serranidae, Pomacentridae y Antennariidae. El trambollito amarillo Starksia fulva fue la especie más abundante al representar 16 % del total de individuos. El gobio gusano pecoso Cerdale ionthas y el gobio Gobulus hancocki representaron respectivamente 10 % y 15 % de la abundancia. Las especies restantes presentaron abundancias menores a 10 % cada una.
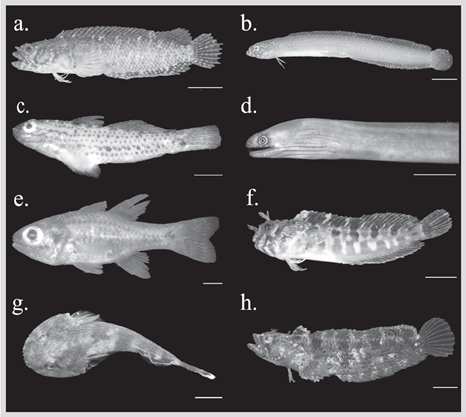
Figura 3 Algunos peces criptobentónicos del PNN Uramba. A) Starksia fulva. B) Cerdale ionthas. C) Coryphopterus urospilus. D) Gymnothorax flavimarginatus. E) Apogon dovii. F) Hypsoblennius caulopus. G) Gobiesox adustus. H) Paraclinus mexicanus. Barra de escala = 5 mm. Fotografías: Arthur Anker.
Tabla 1 Peces criptobentónicos recolectados en SAD (90) expuestos en las seis localidades del PNN Uramba en los cinco eventos de muestreo (2008-2009). Número total de individuos para cada especie (n) junto con el número de individuos medidos (en paréntesis). x̃ ± d.e. = Promedio ± desviación estándar. HA = Hábitos alimentarios. C = Carnívoro. P = Planctívoro. O = Omnívoro (Robertson y Allen, 2008). FO: Frecuencia de ocurrencia. *Nuevos registros para el PNN Uramba. El listado se presenta de acuerdo con la clasificación propuesta por Betancur et al. (2017).
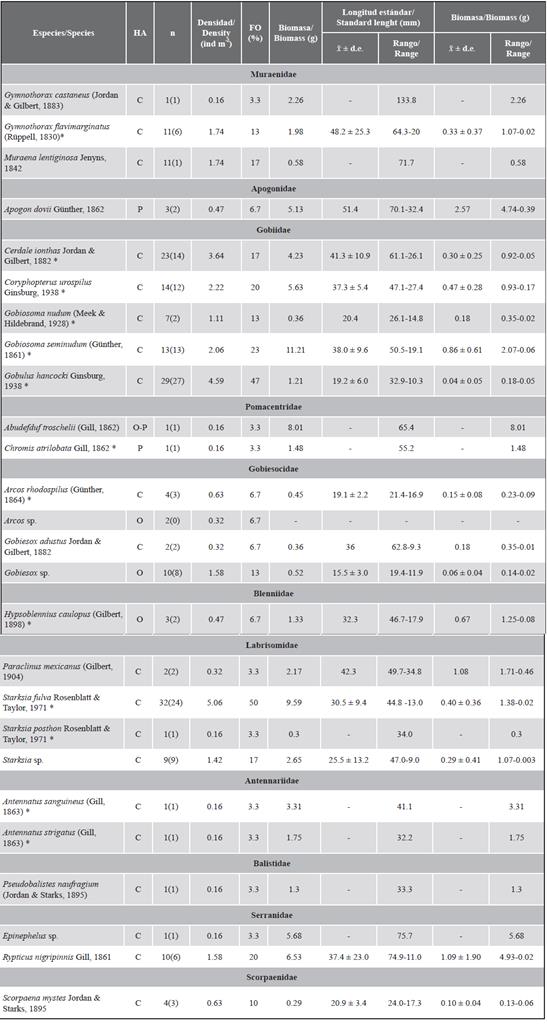
La densidad promedio de los peces fue de 31,2 ± 27,7 ind m-2. Para S. fulva y G. hanckoki fue de 5 ind m-2, mientras que especies con un solo individuo presentaron densidades de 0,16 ind m-2 (Tabla 1). La abundancia promedio fue mayor para OU2 (12,8 ± 10,5 ind m-2) y menor para OU1 (2,6 ± 2,3 ind m-2) y la riqueza promedio observada fue mayor para OU2 (5,4 ± 2,7 ind m-2) y menor para IN1 (1,6 ± 1,1 ind m-2).
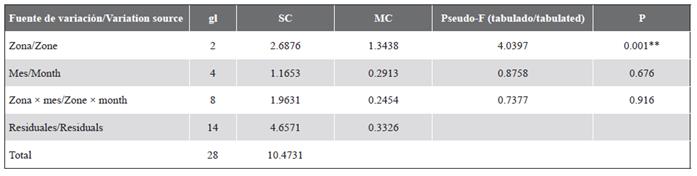
En el nMDS se observó la separación de IN1 y OU2 de las demás localidades (Figura 4) y de la zona interna de las otras zonas. El análisis PERMANOVA de dos vías (zonas y meses) mostró que solo las zonas presentaron diferencias significativas y no se presentó interacción entre los factores (Tabla 2).
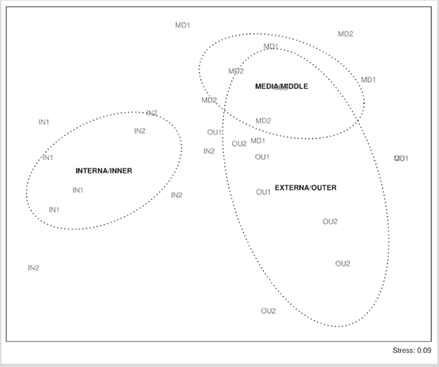
Figura 4. Escalamiento multidimensional no métrico (nMDS) de las localidades y zonas de muestreo de peces criptobentónicos en el PNN Uramba (Pacífico Oriental Tropical).
Tabla 2 Resultados del análisis multivariado permutacional (PERMANOVA) de dos vías evaluando los efectos de las zonas y los meses en la comunidad de peces criptobentónicos en el PNN Uramba (Pacífico Oriental Tropical). gl: Grados de libertad. SC: Suma de cuadrados. MC: Media de cuadrados. P: Nivel de significancia. **: Significativo (P < 0,05).
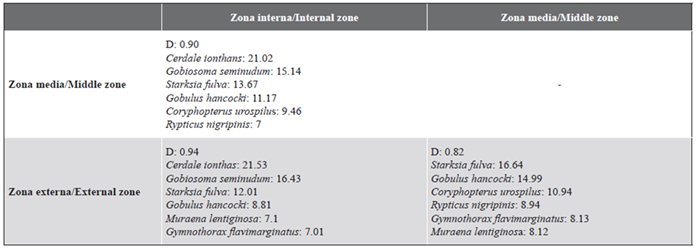
Para la localidad IN1, los gobios Gobiosoma seminudum y C. ionthas representaron 68,8 % y 31,2 % de las similitudes Bray-Curtis promedio. Para IN2 S. fulva fue la especie más representativa al contribuir con 57,4 % de la similitud Bray-Curtis promedio. Gobulus hancocki fue la especie que más contribuyó para MD1 con 65 % de la similitud promedio. Del mismo modo, Coryphopterus urospilus fue la especie más representativa para MD2 con una contribución de 41,6 % de la similitud de esta localidad. Las diferencias encontradas en la comunidad de peces criptobentónicos entre la zona interna y las otras dos zonas se deben a la presencia de C. ionthas, G. seminudum, S. fulva y G. hancocki. Finalmente, la especie más representativa para las localidades OU1 y OU2 de la zona externa fueron respectivamente S. fulva y la morena Gymnothorax flavimarginatus. Las especies mencionadas fueron responsables de las mayores contribuciones en las diferencias entre los pares de zonas al ser evaluadas por medio del análisis SIMPER (Tabla 3).
DISCUSIÓN
La comunidad de peces criptobentónicos del PNN Uramba presenta similitudes con la composición de familias que se han encontrado en los arrecifes coralinos del golfo de California (Balart et al., 2006; González-Cabello y Bellwood, 2009) y del Pacífico panameño (Enochs et al., 2011). En el golfo de California, Tripterygiidae, Gobiidae, Chaenopsidae y Labrisomidae son las familias con mayor número de especies, mientras que Chaenopsidae, Tripterygiidae y Gobiidae son aquellas con mayores abundancias. En el PNN Uramba, Gobiidae y Labrisomidae son las familias dominantes en la comunidad de peces criptobentónicos tanto en riqueza como en abundancia, mientras que Tripterygiidae y Chaenopsidae no se registraron. La ausencia de estas familias puede ser explicada debido a que muchas de las especies de peces de tres aletas (Tripterygiidae) y los trambollines tubícolas (Chaenopsidae) tienen nichos ecológicos especializados que no estuvieron presentes en los SAD. Por ejemplo, en ambientes someros de las zonas media e interna de la bahía se pueden observar especies de Chaenopsidae como el trambollín de los cirripedios Acanthemblemaria hancocki Myers and Reid, 1936, en testas de balanos vacías en sustratos rocosos cercanos a los SAD, así como individuos del pez de tres aletas Axoclinus lucillae Fowler, 1944 (Tripterygiidae) (información sin publicar por los autores) en charcos del intermareal rocoso de la estación más externa (OU2). Si bien en el PNN Uramba no existen arrecifes de coral consolidados como en el golfo de California o el Pacífico panameño, la presencia de comunidades de peces similares indica que la ocurrencia de algunos arrecifes rocosos en esta área protegida puede servir de hábitat para estas especies, ya que dichos arrecifes son un equivalente ecológico de los coralinos, con una estructura heterogénea donde los organismos pueden encontrar refugio, alimento e incluso cumplir el papel de guardería. Además, las especies encontradas, en su mayoría, son altamente tolerantes a las fluctuaciones en la salinidad propias de esta localidad del Pacífico colombiano.
En la costa Pacífica de Panamá se han simulado matrices arrecifales usando restos de esqueletos de corales pocillopóridos, en la cuales Glynn (2006) encontró a Gobulus hancocki entre las especies más abundantes. Sin embargo, el pez críptico más común en ese estudio fue el jaboncillo ocelado Pseudogramma thaumasia (Gilbert, 1900), el cual no fue encontrado en el PNN Uramba, probablemente debido a que esta especie prefiere sustratos duros en zonas de baja turbidez, situación que es poco común en esta AMP.
Desde la perspectiva de los grupos funcionales, la comunidad de peces criptobentónicos del PNN Uramba difiere del descrito para la Gran Barrera de Coral por Depczynski y Bellwood (2003). Estos autores documentaron la predominancia de carnívoros (ca. 70 % de las especies y 85 % de los individuos encontrados en ese estudio) y detritívoros en la comunidad de peces criptobentónicos en la Gran Barrera de Coral. En el POT, Glynn (2006) y González-Cabello y Bellwood (2009) encontraron que los carnívoros y planctívoros son dominantes, mientras que los detritívoros son un componente minoritario de esta comunidad de organismos. Las diferencias en estos grupos tróficos de los peces criptobentónicos que se encuentran en estas regiones (Indo-Pacífico Oriental y POT) puede deberse a la disparidad en las historias geológicas y ambientales de ambos lugares.
Los Negritos, la localidad más externa en este estudio (OU2) es la que presenta menores afectaciones por el drenaje de agua dulce y sedimentos terrígenos. El sustrato predominante allí es rocoso y usualmente presenta una menor turbidez en comparación con las demás zonas. Además, es considerada una de las zonas más biodiversas (Cantera-Kintz, 1991; Guevara-Fletcher et al., 2011) del PNN Uramba. La composición de familias en Los Negritos es similar a la que se ha observado en los charcos intermareales del Parque Nacional Natural Gorgona (Castellanos-Galindo et al., 2014), donde se pueden encontrar arrecifes coralinos adyacentes a la zona intermareal. Muraenidae es una familia dominante en los charcos intermareales en Gorgona, con seis especies que contribuyen significativamente a las densidades y la biomasa de estos ambientes costeros. Por su parte, G. flavimarginatus fue una de las especies que diferenció a OU2 de las demás localidades de esta área protegida. De forma similar, las especies de Gobiesocidae y Antennariidae estuvieron representadas, siendo exclusivas de esta estación de muestreo.
En contraste, IN1 fue dominada por C. ionthas y E. seminudus y se caracterizó por la menor riqueza de especies entre todas las localidades. Estas especies se observan comúnmente en planos lodosos en áreas de manglar cercanas a la localidad de muestreo, donde tienen un papel importante como depredadores de la red trófica de los ecosistemas de manglar. Esto se evidenció gracias al uso de isótopos estables (Medina-Contreras et al., 2018). La ubicación de IN1 en el canal principal del PNN Uramba se caracteriza por fuertes corrientes mareales y una mayor influencia de las condiciones estuarinas, donde la presencia de sedimentos terrígenos es alta y limita el asentamiento de los peces criptobentónicos. En relación con lo anterior, se observa que OU2 y IN1 tienen las mayores diferencias ambientales y en la composición de especies. Sin embargo, S. fulva y G. hancocki se encuentran en la mayoría de las localidades, lo que sugiere que son tolerantes a las condiciones dinámicas típicas de áreas estuarinas.
La proporción de nuevos registros para el área de estudio indica que los SAD son una herramienta adecuada para la colecta de organismos como peces que no son observados fácilmente en los censos visuales y también resaltan la importancia del PNN Uramba como un área biodiversa. En el último listado de especies de peces de esta área se registraron 237 (Castellanos-Galindo et al., 2006). Estos nuevos registros incrementan en 5 % el número total de la diversidad de especies para el parque (alcanzando 250). Es muy probable que en muestreos en otros hábitats del PNN Uramba (e.g. manglares), que utilicen métodos alternativos enfocados en las especies que componen estas comunidades, se incremente el número de especies de peces registradas en esta área protegida. Trabajos preliminares que se han centrado en la endofauna de los manglares en diferentes localidades de la costa central del Pacífico colombiano han revelado que la diversidad tanto de peces como de otros organismos como crustáceos es mayor de lo que tradicionalmente se ha creído (J.F. Lazarus, obs. pers.) y ha permitido encontrar nuevas especies para la ciencia (Anker y Lazarus, 2015a, 2015b, 2017). Lo anterior abre una nueva línea de investigación desde la cual se pueda conocer la diversidad de estos sistemas y evaluar cuál es el aporte de este componente de la fauna en las redes tróficas de los ecosistemas estuarinos.
Por otra parte, es probable que los peces demersales de mayor tamaño pudieran haber escapado de los SAD durante el proceso de recuperación, pero la mayoría de las especies criptobentónicas probablemente se encuentran representadas con este método de muestreo. Glynn (2006) encontró en Panamá que el número de especies de peces comunes entre censos visuales y sustratos artificiales es bajo. Algunos estudios en la Gran Barrera de Coral y en la costa de Nueva Zelanda también han demostrado que existe poca superposición de especies entre los censos visuales y los muestreos en los cuales se ha usado rotenona (Ackerman y Bellwood, 2000; Willis, 2001; Smith-Vaniz et al., 2006). Estructuras similares a los SAD han sido usadas frecuentemente para investigar los patrones de asentamiento de peces en una variedad de hábitats naturales y a diferentes escalas temporales (v. g.Lozano y Zapata, 2003; Mellin y Pontón, 2009). Zimmerman y Martin (2004) usaron un método similar que permitió recolectar de forma efectiva invertebrados arrecifales en el Caribe. Estos resultados permiten demostrar que el uso de los SAD es una alternativa económica, no invasiva y no destructiva que permite tener una muestra representativa de fauna criptobentónica, lo que facilita estudiar diferentes procesos ecológicos que se presentan en las comunidades bentónicas. No obstante, se debe tener cuidado al extrapolar las densidades de peces de los SAD a las densidades de peces observadas en la naturaleza, ya que este método puede constituirse en un sitio de agregación de individuos que en el medio natural se encontrarían dispersos, debido a la disponibilidad limitada de sustratos adecuados.
La densidad de peces criptobentónicos observada en el PNN Uramba (31,2 ind m-2) se encuentra en el rango superior en comparación con otras comunidades de peces estudiadas con técnicas diferentes de muestreo (v. g. rotenona, censos visuales) en otras regiones del mundo. Por ejemplo, Depczynski y Bellwood (2003) utilizaron rotenona para estudiar los peces criptobentónicos en la Gran Barrera de Coral y encontraron densidades de 11,43 ind m-2 (384 individuos en 33,6 m2). Una situación similar ocurrió con González-Cabello y Bellwood (2009), quienes encontraron que la densidad de peces criptobentónicos en el golfo de California era de 20,9 ind m-2. Por lo tanto, se considera que el uso de los SAD es un método sencillo y confiable para realizar estudios a este grupo de organismos.
El presente estudio es uno de los primeros intentos por describir la estructura de las comunidades de peces criptobentónicos de áreas estuarinas en el Pacífico Oriental Tropical. Observaciones preliminares sobre la abundancia de estas especies y los ciclos de vida cortos de muchas de estas permiten suponer que juegan un papel importante en la dinámica trófica de los ecosistemas donde se encuentran (Brandl et al., 2019). Por ello, las interacciones tróficas con otros componentes de los ecosistemas presentes en el PNN Uramba merecen ser investigadas a profundidad (v.g. metabarcoding de contenido estomacal; Casey et al., 2019). En particular, la presencia de un número elevado de carnívoros en los SAD indica que estos sustratos bentónicos heterogéneos en el PNN Uramba proveen también un hábitat a una gran variedad de presas (invertebrados) de las cuales los peces pueden alimentarse.
El bajo potencial de la redundancia funcional, el alto endemismo y el aumento en la frecuencia de eventos ENSO y sus impactos ecológicos son las principales causas de por qué algunos taxa marinos en el POT están amenazados de extinción (Polidoro et al., 2012; Chasqui et al., 2017). Por ello es necesario entender cómo esta comunidad de peces criptobentónicos reacciona espacial y temporalmente a los cambios de las condiciones ambientales que se presentan en los hábitats costeros de la región (v. g. contaminación y desarrollo de infraestructura). Comparar la respuesta de este grupo con otros con una mayor redundancia funcional (v. g. crustáceos, moluscos) permitiría tener una idea de cuáles son los mecanismos necesarios para mantener en óptimas condiciones los ecosistemas marinos. Los peces criptobentónicos podrían entonces ser usados como un modelo para probar cómo diferentes perturbaciones antropogénicas y naturales (v. g. ENSO) afectan el funcionamiento y la resiliencia de los ecosistemas en esta AMP. Este estudio muestra que el PNN Uramba es de vital importancia para la conservación de fauna marina poco conocida de Colombia.
AGRADECIMIENTOS
Este estudio se realizó como parte del proyecto “Vulnerabilidad de los ecosistemas marinos y costeros de Bahía Málaga (Pacífico colombiano): amenazas naturales y antrópicas”, cofinanciado por Minciencias, el Invemar y la Universidad del Valle (Contrato n.° 1106-52128786). Las muestras fueron obtenidas durante el proyecto “Biodiversidad de estadios de vida vulnerable de organismos marinos en Bahía Málaga (Pacifico colombiano) como criterio de conservación”, cofinanciado por Minciencias, Invemar y la Universidad del Valle (Contrato n.° 1106-405-20155). Durante el proceso de escritura, GACG fue financiado por el DAAD a través de una beca otorgada por CEMarin, al igual que JFL, quien contó con una beca del programa de formación doctoral de Minciencias. DMC fue financiada por el programa Jóvenes Investigadores e Innovadores de Minciencias (Contrato n.° 970-2011) y apoyada por una beca doctoral (n.° 734084) del Consejo Nacional de Ciencia y Tecnología (Conacyt) de México y por la Beca de Estímulo Institucional de Formación de Investigadores BEIFI del Instituto Politécnico Nacional de México. Los autores agradecen al grupo de investigación Ecomanglares por su colaboración en la fase de campo, a E. Rubio por su colaboración y consejos durante el proceso de identificación de las especies de peces, a F.A. Zapata quien facilitó información sobre los peces criptobentónicos del Parque Nacional Natural Gorgona, a A. Anker quien amablemente proporcionó las fotos de los peces y dio el permiso para usarlas en esta publicación. Al Programa de Liderazgo en Conservación (CLP), a M. Fischer, a G. Kattan (q. e. p. d.), a A. Giraldo y a H. Rosenthal por sus valiosos comentarios a este manuscrito.
REFERENCIAS
Ackerman, J.L. and D.R., Bellwood. 2000. Reef fish assemblages: a re-evaluation using enclosed rotenone stations. Mar. Ecol. Prog. Ser., 206: 227-237. https://doi.org/10.3354/meps206227 [ Links ]
Ackerman, J.L. and D.R., Bellwood. 2002. Comparative efficiency of clove oil and rotenone for sampling tropical reef fish assemblages. J. Fish Biol., 60: 893-901. https://doi.org/10.1111/j.1095-8649.2002.tb02416.x [ Links ]
Alzate, A., F.A., Zapata, and A., Giraldo. 2014. A comparison of visual and collection-based methods for assessing community structure of coral reef fishes in the Tropical Eastern Pacific. Rev. Biol. Trop., 62: 359-371. https://doi.org/10.15517/rbt.v62i0.16361 [ Links ]
Anker, A. and J.F., Lazarus. 2015a. Description of two new associated infaunal decapod crustaceans (Axianassidae and Alpheidae) from the tropical eastern Pacific. Pap. Avulsos Zool., 55(8):115-129. https://doi.org/10.1590/0031-1049.2015.55.08 [ Links ]
Anker, A. and J.F., Lazarus. 2015b. On two new species of the shrimp genus Salmoneus Holthuis, 1955 (Decapoda, Caridea, Alpheidae) from the tropical eastern Pacific. Zootaxa, 3957(5): 520-534. https://doi.org/10.11646/zootaxa.3957.5.2 [ Links ]
Anker, A. and J.F., Lazarus. 2017. First finding of the shrimp genus Harperalpheus Felder & Anker, 2007 in the eastern Pacific, with description of a new species from Bahía Málaga, Colombia (Malacostraca: Decapoda: Alpheidae). Zootaxa, 4329(2): 196-200. https://doi.org/10.11646/zootaxa.4329.2.7 [ Links ]
Balart, E.F., A., González-Cabello, R.C., Romero-Ponce, A., Zayas-Álvarez, M., Calderón-Parra, L., Campos-Dávila, and L.T., Findley. 2006. Length-weight relationships of cryptic reef fishes from southwestern Gulf of California, Mexico. J. Appl. Ichthyol., 22: 6-318. https://doi.org/10.1111/j.1439-0426.2006.00670.x [ Links ]
Barletta, M., U., Saint-Paul, A., Barletta-Bergan, W., Ekau, and D., Schories. 2002. Spatial and temporal distribution of Myrophis punctatus (Ophichthidae) and associated fish fauna in a northern Brazilian intertidal mangrove forest. Hydrobiologia, 426: 65-74. https://doi.org/10.1023/A:1003939000270 [ Links ]
Beldade, R. and E.J., Gonçalves. 2007. An interference visual census technique applied to cryptobenthic fish assemblages. Vie et Milieu, 57: 61-65. [ Links ]
Betancur, R., E.O., Wiley, G., Arriata, A., Acero, N., Bailly, M., Miya, G., Lecointre, and G., Ortí. 2017. Phylogenetic classification of bony fishes. BMC Evol. Biol., 17: 162. https://doi.org/10.1186/s12862-017-0958-3 [ Links ]
Brandl, S.J., L., Tornabene, C.H.R., Goatley, J.M., Casey, R.A., Morais, I.M., Côté, C.C., Baldwin, V., Parravicini, N.M.D. ,Schiettekatte, and D.R., Bellwood. 2019. Demographic dynamics of the smallest marine vertebrate fuel coral reef ecosystem functioning. Science, 364: 1189-92. https://doi.org/10.1126/science.aav3384 [ Links ]
Cantera-Kintz, J.R. 1991. Etude structurale des mangroves et des peuplements macrobenthiques littoraux de deux baies du Pacifique colombien (Malaga et Buenaventura), rapport avec les conditions du milieu et les perturbations anthropiques. Ph.D. Thesis, Univ Aix-Marseille II, Marseille. 429 p. [ Links ]
Cantera-Kintz, J. R., R., Neira y C., Ricaurte. 1998. Bioerosión en la costa pacífica colombiana: un estudio sobre la biodiversidad, la ecología y el impacto de los animales destructores de los acantilados rocosos. Fondo FEN Colombia, Bogotá. 135 p. [ Links ]
Cantera-Kintz, J.R., E., Londoño-Cruz, L.M., Mejía-Ladino, L. ,Herrera-Orozco, C.A., Satizabal, and N., Uribe-Castañeda. 2013. Environmental issues of a Marine Protected Area in a tectonic estuary in the Tropical Eastern Pacific: Uramba (Malaga Bay, Colombia). context, biodiversity, threats and challenges. J. Water Res. Prot., 5: 1037-1047. https://doi.org/10.4236/jwarp.2013.511109 [ Links ]
Carassou, L., C., Mellin, and D., Pontón. 2009. Assessing the diversity and abundances of larvae and juveniles of coral reef fish: a synthesis of six sampling techniques. Biodivers. Conserv., 18: 355-371. https://doi.org/10.1007/s10531-008-9492-3 [ Links ]
Casey ,J.M., C.P., Meyer, F. ,Morat, S.J., Brandl, S., Planes, and V. ,Parravicini. 2019. Reconstructing hyperdiverse food webs: gut content metabarcoding as a tool to disentangle trophic interactions on coral reefs. Methods Ecol. Evol., 10: 1157-1170. https://doi.org/10.1111/2041-210X.13206 [ Links ]
Castellanos-Galindo, G.A. and U., Krumme. 2013. Tidal, diel and seasonal effects on intertidal mangrove fish in a high-rainfall area of the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser., 494: 249-265. https://doi.org/10.3354/meps10512 [ Links ]
Castellanos-Galindo, G. A., A. ,Giraldo, and E.A., Rubio. 2005. Community structure of an assemblage of tidepool fishes on a Tropical Eastern Pacific rocky shore, Colombia. J. Fish Biol., 67: 392-408. https://doi.org/10.1111/j.0022-1112.2005.00735.x [ Links ]
Castellanos-Galindo ,G.A., J.A., Caicedo-Pantoja, L.M., Mejía-Ladino y E. ,Rubio. 2006. Peces marinos y estuarinos de Bahía Málaga, Valle del Cauca, Pacífico colombiano. Biota Colomb., 7(2): 263-282. [ Links ]
Castellanos-Galindo, G. A., A., Giraldo, and F.A., Zapata. 2014. Tidepool fish assemblages of Gorgona Island, Colombian Pacific coast: a local and regional comparison. Rev. Biol. Trop., 62: 373-390. [ Links ]
Chasqui, L., A., Polanco, A., Acero , P.A., Mejía-Falla, A., Navia, L.A. ,Zapata y J.P., Caldas (Eds). 2017. Libro rojo de peces marinos de Colombia. Instituto de Investigaciones Marinas y Costeras Invemar y Ministerio de Ambiente y Desarrollo Sostenible, Santa Marta. 552 p. [ Links ]
Depczynski, M. and D.R., Bellwood. 2003. The role of cryptobenthic reef fishes in coral reef trophodynamics. Mar. Ecol. Prog. Ser., 256: 183-191. https://doi.org/10.3354/meps256183 [ Links ]
Enochs, I.C., L.T., Toth, V.W., Brandtneris, J.C., Afflerbach, and D.P., Manzello. 2011. Environmental determinants of motile cryptofauna on an eastern Pacific coral reef. Mar. Ecol. Prog., 438: 105-118. https://doi.org/10.3354/meps09259 [ Links ]
Froese, R. and D., Pauly. 2019. FishBase. https://www.fishbase.se 15/11/2019. [ Links ]
Glynn, P.W. 2006. Fish utilization of simulated coral reef frameworks versus eroded rubble substrates off Panama, eastern Pacific. Proc. Coral Reef Symp., 1: 250-256. [ Links ]
González-Cabello, A. and D.R., Bellwood. 2009. Local ecological impacts of regional biodiversity on reef fish assemblages. J. Biogeogr., 36: 1129-1137. https://doi.org/10.1111/j.1365-2699.2008.02065.x [ Links ]
Guevara-Fletcher, C.E., J.R., Cantera-Kintz, L.M., Mejía-Ladino, and F.A., Cortes. 2011. Benthic macrofauna associated with submerged bottoms of a tectonic estuary in Tropical Eastern Pacific. J. Mar. Sci., 2011: 1-13. https://doi.org/10.1155/2011/193759 [ Links ]
Loreau, M. 2004. Does functional redundancy exist? Oikos, 104(3): 606-611. https://doi.org/10.1111/j.0030-1299.2004.12685.x [ Links ]
Lozano, S. and F.A., Zapata. 2003. Short-term temporal patterns of early recruitment of coral reef fishes in the Tropical Eastern Pacific. Mar. Biol., 142: 399-409. https://doi.org/10.1007/s00227-002-0948-9 [ Links ]
Martínez, J.O. and E., López-Ramos. 2011. High-resolution seismic stratigraphy of the late Neogene of the central sector of the Colombian Pacific continental shelf: a seismic expression of an active continental margin. J. S. Am. Earth. Sci., 31: 28-44. https://doi.org/10.1016/j.jsames.2010.09.003 [ Links ]
Medina-Contreras, D., J.R., Cantera-Kintz, A., Sánchez-González, and E., Mancera. 2018. Food web structure and trophic relations in a riverine mangrove system in the tropical eastern Pacific, central coast of Colombia. Estuaries Coast, 41(5): 1511-1521. https://doi.org/10.1007/s12237-017-0350-y [ Links ]
Mellin, C. and D., Pontón. 2009. Assemblages of reef fish settling on artificial substrates: effect of ambient habitat over two temporal scales. Mar. Freshwater. Res., 60: 1285-1297. https://doi.org/10.1071/MF08319 [ Links ]
Oksanen, J.F., G., Blanchet, M., Friendly, R., Kindt, P., Legendre, D., McGlinn, P.R., Minchin, R.B., O’Hara, G.L., Simpson, P., Solymos, M.H.H. ,Stevens, E., Szoecs, and H., Wagner. 2019. vegan: community ecology package. R package version 2.5-6. https://CRAN.R-project.org/package=vegan 20/02/2020. [ Links ]
Polidoro, B., T., Brooks, K.E., Carpenter, G.J., Edgar, S., Henderson, J., Sanciangco, and D.R., Robertson. 2012. Patterns of extinction risk and threat for marine vertebrates and habitat-forming species in the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser., 448: 93-104. https://doi.org/10.3354/meps09545 [ Links ]
R Core Team. 2019. R: a language and environment for statistical computing. https://www.R-project.org 05/12/2019. [ Links ]
Ransome, E., J.B., Geller, M., Timmers, M., Leray, A., Mahardini, A., Sembiring, A.G., Collins, and , Meyer, C.P. (2017). The importance of standardization for biodiversity comparisons: a case study using autonomous reef monitoring structures (ARMS) and metabarcoding to measure cryptic diversity on Mo’orea coral reefs, French Polynesia. PloS ONE, 12(4), e0175066. https://doi.org/10.1371/journal.pone.0175066 [ Links ]
Robertson, D.R. and G.R., Allen. 2008. Shorefishes of the Tropical Eastern Pacific online information system. Smithsonian Tropical Research Institute, Balboa. https://biogeodb.stri.si.edu/sftep 05/10/2009. [ Links ]
Robertson, D.R. and W.F., Smith-Vaniz. 2008. Rotenone: an essential but demonized tool for assessing marine fish biodiversity. Bioscience, 58(2): 165-170. https://doi.org/10.1641/B580211 [ Links ]
Robertson, D.R., G.D., Green, and C.B., Victor. 1988. Temporal coupling of production and recruitment of larvae of a Caribbean reef fish. Ecology, 69: 370-381. https://doi.org/10.2307/1940435 [ Links ]
Smith-Vaniz, W.F., H.L. ,Jelks, and L.A., Rocha. 2006. Relevance of cryptic fishes in biodiversity assessments: a case study at Buck Island Reef National Monument, St. Croix. Bull. Mar. Sci., 79: 17-48. [ Links ]
Valles, H., D.L., Kramer, and W., Hunte. 2006. A standard unit for monitoring recruitment of fishes to coral reef rubble. J. Exp. Mar. Biol. Ecol., 336: 171-183. https://doi.org/10.1016/j.jembe.2006.05.005 [ Links ]
Willis, T.J. 2001. Visual census methods underestimate density and diversity of cryptic reef fishes. J. Fish Biol., 59: 1408-1411. https://doi.org/10.1006/jfbi.2001.1721 [ Links ]
Zimmerman, T.L. and J.W., Martin. 2004. Artificial reef matrix structures (ARMS): an inexpensive and effective method for collecting coral reef-associated invertebrates. Gulf Caribb. Res., 16: 59-6. https://doi.org/10.18785/gcr.1601.08 [ Links ]
Recibido: 09 de Agosto de 2020; Aprobado: 30 de Octubre de 2020
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
