











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
The phylum Echinodermata is the dominant megafauna taxon in hard and soft bottom communities of the Antarctic and sub-Antarctic regions in terms of abundance, biomass, and diversity (Moles et al., 2015). Likewise, their variety of feeding habits makes these invertebrates play an important role in the trophic energy transfer and therefore, in the communities structure in Antarctic benthic systems (McClintock, 1994; Moles et al., 2015). The Antarctic Ocean is currently the focus of many investigations in which it has been shown that climate change and anthropogenic influences have affected the environment in this region and therefore these alterations are relevant to understand the evolution and current biological structures of the community that inhabits there, and consequently, its connectivity with the biota outside the Polar Front (Vaughan et al., 2003; Barboza et al., 2011). Echinoderms have been a good component for these studies, given their evolutionary characteristics, great dispersion, and variety of habits, being studied not only in connectivity and endemism research, but also in ocean acidification and trophic networks (McClintock, 1994; Dahm, 1999; Barboza et al., 2011; Duquette et al., 2018). Regarding their diversity, it is currently known that around 10 % of the macrozoobenthic species known in Antarctica are echinoderms (Moles et al., 2015). There are inventories of the classes Asteroidea (299 species, Moreau et al., 2018), Ophiuroidea (126 species, Stöhr et al., 2012), Echinoidea (201 species, Fabri-Ruiz et al., 2017) and Holothuroidea (187 species, O’Loughlin et al., 2011). The Crinoidea class, although well represented (Eléaume et al., 2014) does not have a compiled inventory of species. Specifically, around the Antarctic peninsula, studies have focused primarily on the South Shetland Islands and the Weddell Sea (Gutt and Piepenburg, 1991; Massin 1992; Piepenburg et al., 1997; Moles et al., 2015; Martínez et al. al., 2015), and in many studies, the information is generally recorded for the peninsula without detailing localities. Few studies specifically include information on the Palmer Archipelago or Trinidad Island that form the Gerlache Strait (Greig, 1929; Manjón-Cabeza et al., 2001; Manjón-Cabeza and Ramos, 2003). Taking into account the diversity and ecological importance of echinoderms in Antarctic and sub-Antarctic communities, as well as the little specific information for the Gerlache Strait and adjacent islands, this work contributes to the inventory of echinoderms in this area of the Antarctic Peninsula, expanding the knowledge of the species, especially in terms of their geographical and bathymetric distribution. Incidentally collected samples contribute to the baseline that is being developed to generate new contributions to the inventories and biological information of the Gerlache Strait, in order to define possible conservation areas in this region. These results are part of the “Biodiversity and oceanographic conditions of the Gerlache Strait, Biogerlache-Antarctica” project, which is included in the Antarctic scientific agenda of the Marine and Coastal Research Institute- Invemar, which began in 2014 with three working approaches on climate change, conservation, and biodiversity.
MATERIALS AND METHODS
Study area
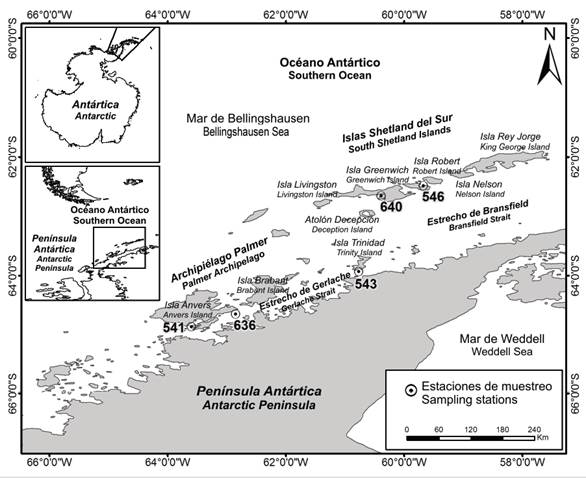
The Gerlache Strait is located to the west of the Antarctic Peninsula, separating it from the Palmer Archipelago, extending in a southwest to northeast direction including Trinidad Island, it has a depth between 270 and 1100 m, with slopes of 20 ° (Figure 1) (Anderson, 1999). Together with the Bransfield Strait (Figure 1), 220 km long and 120 km wide between the Antarctic Peninsula and the numerous islands that make up the South Shetland Islands archipelago, they constitute sites of interest to the scientific community; since they offer the opportunity to study the composition, structure, and dynamics of various marine communities, particularly benthic fauna (Secretariat of the Antarctic Treaty, 2009).
Sampling
During the Colombian scientific expeditions to Antarctica carried out between 2016 and 2019 (III - Admiral Padilla Expedition and V - Admiral Campos Expedition), 11 stations were sampled to characterize the soft bottom benthic community (Table 1). The samplings were carried out using sediment grab samplers: a Shipek dredge (0.04 m2) during the III Expedition and a Van Veen grab (0.066 m2) during the V Expedition. Once the grab samplers were deployed and recovered, before sieving and preserving the sediment, the mega-epifauna and the material of the dredging upper layer were manually separated (plant remains, small rocks, among others), to take advantage of these incidentally collecting. The mega-epifauna was anesthetized in a solution of filtered seawater with 10 % magnesium chloride in the dark, and later fixed and preserved in 96 % alcohol. The upper layer material was directly preserved in 96 % alcohol.
Table 1 Stations sampled during the Colombian scientific expeditions to Antarctica: III Admiral Padilla Expedition (2016-2017) and V Admiral Campos Expedition (2018-2019). Echinoderms were collected at the stations indicated in bold and with an asterisk.
The specimens and samples were stored in labeled bags or containers and transported to Invemar. In laboratory, the mega-epifaunal echinoderms were reviewed and separated by morphotypes. Likewise, the macro-epifauna present in the sediments upper layer was reviewed using stereo microscopes and the observed specimens were separated by morphotypes. All the morphotypes found were identified and the photographic record was made according to the morphological characteristics of interest of each one. For taxonomic identification, the following references were used for each class: Crinoidea: Messing (2019, 2020a, b, c, d), Antarctic Underwater Field Guide (2019); Asteroidea: Clark and Downey (1992), Janosik and Halanych (2010), Peck et al. (2018); Ophiuroidea: Bell (1902), Koehler (1912), Matsumoto (1915, 1917), Mortensen (1936), Fell (1960), A.M. Clark (1970), Mortensen (1927) and Holothuroidea: Miller and Pawson (1984), O’Loughlin and VandenSpiegel (2010), O’Loughlin et al. (2010), Thandar (2013), Martínez (2014), Moura et al. (2015), O’Loughlin et al. (2015). The collected specimens were deposited in the echinoderms collection of the Marine Natural History Museum of Colombia (MHNMC) of Invemar (INV EQU) enriching the biological collections and generating the possibility of participating in comparative taxonomic works.
RESULTS
Of the eleven stations sampled, only in five of them echinoderms were found (Figure 1, Table 1), collecting a total of 29 individuals, identified in 13 morphotypes belonged to the classes Ophiuroidea (5 morphotypes), Holothuroidea (4), Asteroidea (2) and Crinoidea (2) (Table 2). The largest number of morphotypes (9 in total) was found at Station 541, located at Yelcho Base in the Gerlache Strait, while at station 543 (Trinidad Island) two morphotypes were found, and one morphotype in the station 636 (Anvers Island) (Figure 1, Table 2). At the South Shetland Islands sampled stations, three morphotypes were collected, two at station 640 (Livingston Island) and one at 546 (Greenwich Island) (Figure 1, Table 2). Six of the identified morphotypes are mega-epifauna organisms that were separated directly in the field, the remaining seven morphotypes were found after reviewing the dredges upper layer material using stereo microscopes (Table 2). The main morphological characteristics, as well as the geographical and bathymetric distribution of the species collected and some comments, are described below.
Table 2 List of collected echinoderms. Expeditions: III - Admiral Padilla Expedition (2016-2017); V - Admiral Campos Expedition (2018-2019); Stations: 541, 543, and 636 Gerlache Strait; 546 and 640 South Shetland Islands (Figure 1, Table 1); G-Mega-epifauna; C-Macro-epifauna.
Class Crinoidea
Two individuals identified as Promachocrinus kerguelensis Carpenter, 1879 and Anthometrina adriani (Bell, 1908) were collected (Figure 2, Table 2). Promachocrinus kerguelensis was collected in the Gerlache Strait at 54 m depth. It is characterized by presenting 20 arms that arise from ten radial ossicles (Figure 2a, b). Its distribution is circumpolar between 65 and 1162 m depth and it is a common species in the Antarctic peninsula, registered in the South Shetland Islands (Hemery et al., 2012; Eléaume et al., 2014). It is considered the most abundant and morphologically variable crinoid species in the Southern Ocean (Hemery et al., 2013). It is currently the only accepted species in the genus, several previously described species have been synonymized (Messing, 2019; Messing, 2020a, b). However, based on molecular data (mitochondrial DNA), seven sympatric lineages have been found in Antarctica (Hemery et al., 2012; Eléaume et al., 2014).

Figure 2 Class Crinoidea. Promachocrinus kerguelensis a. Oral view and b. Lateral view; Anthometrina adriani c. General view and d. - e. Detail of the arms, centrodorsal, and cirri.
Anthometrina adriani was also collected in the Gerlache Strait at 523 m depth. It is characterized by presenting 10 arms that arise from 5 radial ossicles, distal segments of the oral pinnules with a comb-shaped structure, conical centrodorsal covered by numerous cirri, and well-developed “synarthrial” tubercles. (Figure 2d-e) (Messing, 2019; Messing, 2020d). Like P. kerguelensis, it is a circumpolar species, although it is found in the Antarctic peninsula, its populations in this area have few individuals compared to those in eastern Antarctica (Eléaume et al., 2014). Furthermore, it is a species confined to the high Antarctic shelf (approximately above 67 ° S), which has not been recorded from the South Shetland Islands, the Scottish arc, or the sub-Antarctic islands (Eléaume et al., 2014), so this record extends its geographic distribution to 64 ° 39 ‘S.
Class Asteroidea
Two individuals were collected, one was identified as Odontaster validus Koehler, 1906, and the other was identified to the taxonomic level of Family (Asteriidae) (Figure 3, Table 2). The specimen identified in the Family Asteriidae was collected in Puerto Mikelsen (Trinidad Island, Gerlache Strait) at 98 m depth. It could be a juvenile considering its size (R = 4 mm), however, the distribution of the abactinal spines, in two lateral rows and a central one (Figure 3a, b) is a characteristic on the basis of which identification could be made to a lower taxonomic level. Asteriidae is one of the most diverse families in Antarctica, in some works it has been recorded with the largest number of species, overcoming Astropectinidae and Goniasteridae (Majón-Cabeza et al., 2001).

Figure 3 Class Asteroidea. Asteriidae sp. a. Dorsal view and b. Ventral view; Odontaster validus c. Dorsal view, d. Ventral view, e. Detail of the paxilas, f. Detail of the interradial areas.
Odontaster validus was collected on Livingston Island (South Shetland Islands) at 57 m depth. This starfish is characterized by having the paxilas of the radial abactinal area with approximately 12 small, smooth, thin, and conical spines (Figure 3e), marginal plates covered by spines, without granules (Figure 3c), actinal plates covered by up to seven small thin spines, and two or three furrow spines (Figure 3d, f). Janosik and Halanych (2010) described two closely related species, O. roseus and O. pearsei which have been confirmed using DNA barcodes, however, Peck et al. (2018) have registered a high variability in some of the morphological characteristics that distinguish the three species. O. validus is the most common and abundant starfish species in the shallow environments of the Antarctic continent (Dearborn, 1977; Neill et al., 2016), its distribution is circumpolar, including South Georgia Island and Bouvet Island (Janosik and Halanych, 2010). Moles et al. (2015) record it in Deception Atoll (South Shetland Islands) and in the Weddell Sea.
Class Ophiuroidea
Twenty specimens of this class were collected belonged to five morphotypes, one of them was identified to family (Amphilepididae), another to genus (Amphiophiura), and the others were identified to species: Ophionotus victoriaeBell, 1902, Amphiura cf. joubiniKoehler, 1912 and Amphiura cf. microplaxMortensen, 1936, although two of them have not yet confirmed (Figure 4, Table 2). O. victoriae was collected on Greenwich Island (South Shetland Islands) at 29 m depth; the other four morphotypes were collected at Base Yelcho in the Gerlache Strait at 54 m depth. Among them, Amphiophiura sp. was the only morphotype found in more than one station, in Mikelsen Port-Trinidad Island, and also in the Strait at 98 m depth; furthermore it is the morphotype with the highest number of individuals in the samplings.
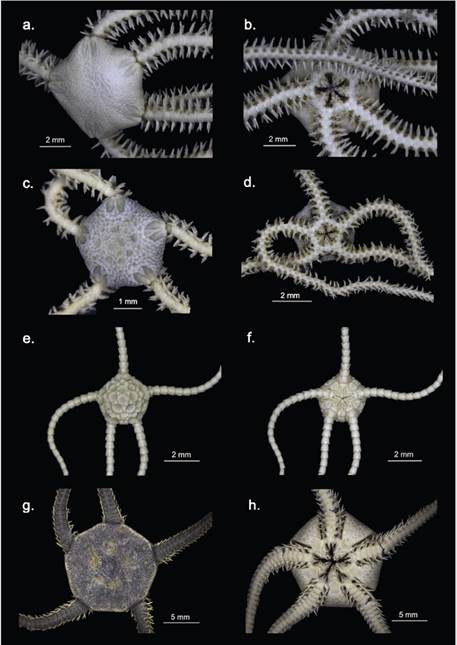
Figure 4 Class Ophiuroidea. Amphiura cf. joubini a. Dorsal view, b. Ventral view; Amphiura cf. microplax c. Dorsal view, d. Ventral view; Amphiophiura sp. e. Dorsal view, f. Ventral view; Ophionotus victoriae g. Dorsal view, h. Ventral view; Amphilepididae i. Dorsal view, j. Ventral view.
Amphiura joubini (Figure 4a, b) is characterized by presenting a disc covered with small and fine scales, being smaller on the ventral surface; elongated and narrow radial shields (four times as long as wide); small and triangular oral shields, as long as wide; block-shaped thick proximal oral papillae and spiniform distal oral papillae; large oral scale; large and pentagonal dorsal brachial plates, wider than long; six brachial spines and two small tentacular scales, of which the outer one is larger. The specimens collected agree with all mentioned characteristics, however, they have four brachial spines and the entire ventral surface of the disc is covered by small scales (Figure 4a, b), which differs with the diagnosis and with some descriptions of this species where it is mentioned that the area near the oral shield is “partially bare” (Koehler, 1912; Brogger and O’Hara, 2015). A. joubini is a common species in Antarctica, being recorded in gravel and sand from 15 to 2345 m depth in the Antarctic Peninsula, the Bellingshausen Sea, and the Weddell Sea and adjacent islands (Koehler, 1912; Moles et al., 2015). It is frequently confused with A. princeps Koehler, 1907 due to its great morphological similarity, however, the latest has rounded distal oral papillae (Brogger and O’Hara, 2015) and seems to have a shallower bathymetric distribution in the Southern hemisphere (Alitto et al., 2018). However, Brogger and O’Hara (2015) emphasized the need to review the records of these species to correctly establish their geographic and bathymetric distributions.
Amphiura microplax (Figure 4c, d) is characterized by presenting a pentagonal disc covered with medium scales, among which the six primary plates (rosette) stand out; short radial shields, separated in its proximal part; ventral surface of the disc covered with small scales; pentagonal oral shields; large triangular adoral shields, touching at their proximal end; square ventral brachial plates; rhomboidal dorsal brachial plates; five short and conical brachial spines in the basal segments and a small tentacular scale, the size of this scale differentiates A. microplax from the other species of the genus with a single tentacular scale that are recorded for Antarctic waters (Mortensen, 1936). The specimens collected agree with this diagnosis, however, as there are not enough photographs and bibliography to compare, their identification has not yet been confirmed (Figure 4c, d). This species is viviparous and parthenogenic (Mortensen, 1936), has been recorded in the Antarctic Peninsula, Weddell Sea, Enderby Plain, South Georgia Island, South Orkney Islands, South Shetland Islands, and the South Sandwich Islands, between 96 and 486 m depth (Dahm, 1999; Moles et al., 2015). Once this identification has been confirmed, this record would correspond to the shallower record of A. microplax (54 m).
The species of the genus Amphiophiura are characterized by presenting a swollen disc, covered with scales and plates, among which the primaries are very prominent, as are the radial shields; arm combs and genital papillae present; second tentacular pore outside the oral cleft and bordered by several tentacular scales; arms not very long and with blunt termination; dorsal and ventral brachial plates well developed; long lateral brachial plates, with short spines and large tentacular pores with numerous scales (Matsumoto, 1915). The collected specimens present the diagnostic characters of the genus and correspond to the same morphotype still undetermined up to a taxonomic level of species; whereby, it is presented as Amphiophiura sp. (Figure 4e, f). This genus contains scavengers and predators (Frensen et al., 2010), of which there are four recorded species for Antarctic waters at depths between 66 and 4773 m (Frensen et al., 2010; Moles et al., 2015).
Ophionotus victoriae (Figure 4g, h) is characterized by presenting a disc covered by several small and fine scales; five arms; second tentacular pore outside the oral cavity and bordered by several tentacular scales; small lateral brachial plates, dorsal brachial plates fragmented into a central plate with three to four small plates on the sides; and genital papillae along the genital clefts, which reach the disc edge (Bell, 1902; Koehler, 1912) ( Figure 4g, h). O. victoriae is circumpolar and possibly the most abundant and common ophiuroid in the Antarctic benthos, with a wide distribution in upper Antarctica, in the South Shetland Islands, South Georgia Island, South Orkney Islands, South Sandwich Islands, the Antarctic Peninsula, Gerlache Strait, Ross Sea, Wedell Sea, Amundsen Sea, Bouvet Island, and the Enderby Plain (Dahm, 1999; Moya et al., 2003; Moles et al., 2015). It inhabits a variety of substrates, has been found in soft bottoms (mud and gravel) and rocky bottoms; between 0 and 1704 m depth (Grange et al., 2004; Moles et al., 2015). Its diet is very varied, since, being an opportunist, it feeds on a great variety of invertebrates such as sponges, polychaetes, bivalves, copepods, diatoms, asteroids, among others; it is also a scavenger and cannibal of juveniles of the same species (Fratt and Dearborn, 1984; Dahm, 1996; Grange et al., 2004); it even consumes, if necessary, organic particulate material deposited on the bottom such as fecal matter, including its waste. This plasticity in the feeding strategy has provided certain advantages such as accelerated growth in the first years (Dahm, 1996).
The family Amphilepididae is characterized by presenting a disc covered with interspersed scales, among which the primaries are very evident; the radial shields, unlike its sister group the family Amphiuridae, are broad and well developed; it presents triangular oral papillae, the infradental papillae are not block-shaped and it presents an apical papilla, also, the second tentacular pore is outside the oral cavity (Matsumoto, 1915, 1917; Mortensen, 1927). The specimen collected presents the diagnostic characters of the family (Figure 4i, j), however, due to the taxonomic difficulty of the species within the superfamily Amphiuroidea, a lower taxonomic category has not yet been reached. Since Matsumoto (1915) erected the family Amphilepididae based on jaw characters to differentiate it from the Family Amphiuridae, it has presented numerous taxonomic changes throughout history, which has made it difficult to correctly identify the genera and species that compose it (Koehler, 1922; AM Clark, 1970). Even, due to its great similarity with specimens of the genera Histampica, Amphiura, and Amphioplus, some authors such as A.M. Clark (1970) have proposed treating Amphilepididae as a subfamily of Amphiuridae. However, Amphilepididae is currently a valid family with two accepted genera: Amphilepis Ljungman, 1867, and Ophiomonas Djakonov, 1952 (Stöhr et al., 2018), distributing mainly in the northern hemisphere, but with some records in the southern hemisphere, including South America, Madagascar, and Antarctica (Mills and O’Hara, 2010). The individual collected presents a pentagonal disc covered by scales; swollen and triangular radial shields; triangular oral shields; two oral papillae on each side of the mandible, the proximal one is triangular, and also has an apical papilla or ventral tooth; arms thin, inserted ventrally to disc; large tentacular pores, the second tentacular pore is outside the mouth and is provided with a large tentacular scale, the following tentacular pores without scales (Figure 4i, j).
Class Holothuroidea
Four specimens of this class were collected, identified as Taeniogyrus cf. prydziO`Loughlin and VandenSpiegel, 2010, Trachythyone cf. parva (Ludwig, 1875), Psolus sp. and Molpadia violacea Studer, 1876 (Figure 5, 6, Table 2). M. violacea was collected on Livingston Island (South Shetland Islands) at 29 m depth; the other three morphotypes were collected at the Yelcho Base in the Gerlache Strait at 54 m depth.
Moura et al. (2015) described the new species Taeniogyrus yvonnae Moura, Campos and Esteves, 2015; as part of their work they presented a review of the genus, which is distributed mainly in the Indo-Pacific, with some species in the Antarctic, the Mediterranean, and the Atlantic Ocean. The genus includes 24 species, of which two are distributed in Antarctica T. prydziO’Loughlin and VandenSpiegel, 2010 and T. antarcticus Heding, 1931. Two other species distributed in Antarctica according to O’Loughlin et al. (2010) are currently accepted as species of the genus Sigmodonta Studer, 1876 (WoRMS, 2020). Specimens collected in the Gerlache Strait at 54 m depth coincide with T. prydzi in body length (maximum 50 mm) (Figure 5a), sigmoid hooks size (232-272 µm) and wheels size (90 µm) (Figure 5c), however, they present tentacles with 3 to 4 pairs of digits, differing from T. prydzi which has 6 pairs (Figure 5b). T. prydzi is distributed in East Antarctica on the MacRobertson shelf between 109 and 830 m depth, so this would be the first record of the species for West Antarctica, specifically for the Antarctic Peninsula. For the genus, this record is the first in the peninsula considering that T. antarticus Heding, 1931 is distributed in the South Georgia Islands, the Scotia Sea, the South Orkney Islands and the Aurora Islands (Moura et al., 2015). Also, the depth at which the specimens were collected would widen the bathymetric distribution range of the species from 109 m depth, previously recorded by Moura et al. (2015), up to 54 m.
Molpadia violacea was considered synonym of M. musculus Risso, 1826, a species of cosmopolitan distribution, which included Antarctica, and with a wide bathymetric distribution range from 35 to 5205 m depth (Pawson, 1977; O’Loughlin et al., 2010). However, O’Loughlin et al. (2015) established once again M. violacea as a valid species to refer to Molpadia specimens from Antarctica previously referred to M. musculus. This decision was based on molecular evidence presented by O’Loughlin et al. (2010), and morphological characteristics that include the body color, with the oral and caudal ends whitish (Figure 5d), the absence of spires in the caudal fusiform rods (Figure 5f) and the presence of phosphatic bodies in the body wall (Figure 5e, f). Very few ossicles are observed in the middle of the body of the collected specimen, coinciding with O’Loughlin et al. (2015) on the phosphatizing and disappearance of ossicles in this species. M. violacea is distributed between 112 and 628 m depth in the Southern Ocean and in Antarctica where it has been recorded in Prydz Bay, the Ross Sea, near South Orkney Island and in the South Shetland Islands (O’Loughlin et al., 2015), where the specimen recorded in this work was collected. This is one of the few records of the species after having been separated from M. musculus, and also extends its bathymetric distribution from 112 m up to 57 m depth.
Trachythyone parva is characterized by presenting a U-shaped body, tube feet restricted to ambulacra, although they may be present in the inter-ambulacra at least in dorsal part (Figure 6a-c); body wall with basket-shaped ossicles developed as curved bowls, edge with short spiny projections (Figure 6d); crosses present, sometimes excluding baskets (Figure 6d); small plates (up to 1.5 mm), smooth, with numerous small holes in usually double series (Thandar, 2013; Martínez, 2014). Although most of these characteristics have been found in the collected specimen, differences in the plates are observed, in which the holes are irregularly distributed (Figure 6d). Also, the plates have projections at the ends, which are not mentioned in the diagnosis of T. parva. O’Loughlin et al. (2010) list nine species of the genus Trachythyone in Antarctica, two of them found in that work and not yet described, one of them “Trachythyone species 1” is distributed in the South Shetland Islands, near the Gerlache Strait, where the specimen described in present work was collected. However, if the species were confirmed as T. parva, this would be the first record for the Antarctic Peninsula, the previous ones are from Tierra del Fuego, the Falkland Islands up to 38ºS (Martinez, 2014), the Weddell Sea, and other locations of East Antarctica (GBIF, 2020).
The specimen identified as Psolus sp. has a length of 2 mm, so it could be a juvenile (Figure 6f, g), the review of morphological characteristics such as the shape of the spicules, necessary to identify the species, would involve its destruction. According to O’Loughlin et al. (2010) there are 21 species of the genus Psolus in Antarctica. More specimens are needed in order to review their morphology with more detail and make an accurate identification.
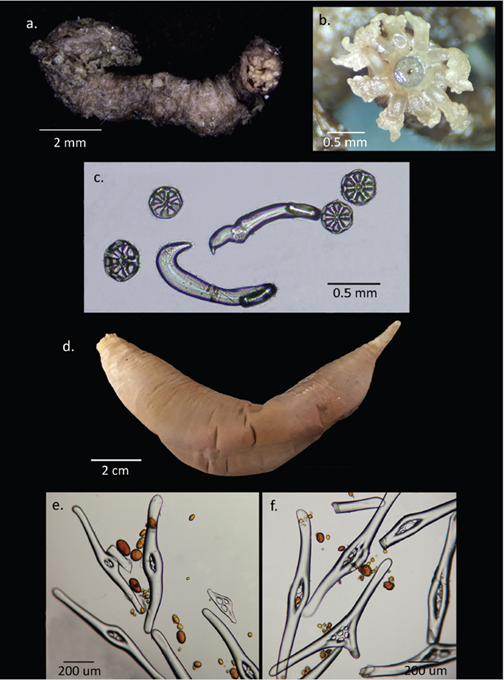
Figure 5 Class Holothuroidea. Taeniogyrus cf. prydzi. a. General view (Specimen 1), b. Detail of the mouth and tentacles (Specimen 2), c. Body wall ossicles shaped like sigmoid hooks and wheels; Molpadia violacea d. General view, e. Ossicles of the anterior body wall, f. Ossicles of the posterior body wall, in both images phosphatic bodies are observed.

DISCUSSION
Studies and inventories on Antarctic echinoderms have shown this group as one of the most diverse in the shallow and deep seabed communities in this region (O’Loughlin et al., 2010; Stöhr et al., 2012; Eléaume et al., 2014; Martín-Ledo and López-González, 2014; Fabri-Ruiz et al., 2017; Moreau et al., 2018). In the case of sea cucumbers, O’Loughlin et al. (2010) proposed the Antarctic region as a biodiversity “hotspot” for the class Holothuroidea, presenting numbers that are equivalent to 4 % of the marine diversity documented in Antarctica and 10 % of the diversity of Holothuroidea in the world. Likewise, the class Asteroidea in the Antarctic and Sub-Antarctic region is well represented, with 12 % of the total species richness of the benthic groups in this area (Moreau et al., 2018).
The number of identified morphotypes, 13 in total, based on 29 specimens collected in five stations confirms the high richness of this group in Antarctica, however, this is a low number of species compared to other works that include information on areas close to the Gerlache Strait, such as the South Shetland Islands, and even if it is taken into account that no echinoderms were collected in six of the eleven stations sampled and no representatives of the class Echinoidea were found. This result is due to the fact that the samplings carried out were aimed at characterizing the benthic macrofauna and not the mega and macro-epifauna. Martinez et al. (2015) based on 13 stations in the South Shetland Islands, four in the extreme south of the Antarctic peninsula, and two in the South Orkney Islands (Campaign CAV-III), recorded echinoderms in all the sampled stations, being Ophiuroidea the best represented class being found in 100 % of the stations, followed by Asteroidea with 90.5 %, Holothuroidea with 81 %, Crinoidea with 66.7 %, and lastly Echinoidea with the lowest percentage, being collected in 62.4 % of stations.
However, from another perspective, considering that the information presented in this work comes from five stations and from a sampling that was not addressed to these organisms, it is important to highlight the high number of morphotypes found and the relevance of these records for the knowledge of some species, as well as for the elaboration of the species inventory of the Gerlache Strait. The present record of the crinoid Anthometrina adriani extends its geographic distribution to 64 ° 39 ‘S. In the case of the sea cucumber Taeniogyrus cf. prydzi this is the first record of the genus for the Gerlache Strait and the Antarctic Peninsula, and if the species is confirmed, this would be its first record for West Antarctica. In some cases such as Amphiura cf. microplax this could be the first record for the Gerlache Strait, however, it is sometimes difficult to establish these first records because the Antarctic Peninsula is included in the distribution of many species but without specifying the locations. Furthermore, there are not as many specific inventories for the Gerlache Strait or the adjacent islands, comparing to other areas of Antarctica. Regarding the bathymetric distribution of Promachocrinus kerguelensis, Amphiura cf. microplax, Taeniogyrus cf. prydzi, and Molpadia violacea, the records presented in this work extend their depth range, being the shallower for these species (Station 541: 54 m depth and 640: 57 m depth). These results highlight the need and importance of carrying out samplings in future expeditions focused on the characterization of the mega and macro-epifauna of the Gerlache Strait and the adjacent islands, being one of the least studied areas in the Antarctic peninsula, especially compared to other locations such as the South Shetland Islands.
Some of the species that have been identified so far are also among the most common and abundant in shallow ecosystems of this region, in particular the crinoid Promachocrinus kerguelensis, the starfish Odontaster validus, and the brittle star Ophionotus victoriae of which ecological, reproductive, biogeographic, among others aspects, are known, as was mentioned in the comments presented in the results section. In the expeditions carried out, O. victoriae was collected only in the South Shetland Islands, where apparently the community structure of the class Ophiuroidea is heterogeneous and clearly determined by the variation in the abundance of O. victoriae, which, being a species of opportunistic habits, exhibits an evident dominance in the archipelago (Manjón-Cabeza and Ramos, 2003; Moya et al., 2003). However, in the other Antarctic and sub- Antarctic areas, this species presents a similar abundance to representatives of the genera Ophioplinthus and Ophiacantha (Manjón-Cabeza and Ramos, 2003). Although the sampling methodology does not allow to make inferences about the ophiuroids abundance, it is worth highlighting previous studies, where the community structure of the class Ophiuroidea in the Antarctic peninsula differs from the South Shetland Islands (Manjón-Cabeza and Ramos, 2003; Moya et al., 2003).
In several cases, taxonomic identification to species level has not been possible due to several reasons, including that some specimens are possibly juveniles or that some characteristics could not be confirmed due to lack of references to compare, or because the small size of some unique specimens. In the case of sea cucumbers, O’Loughlin et al. (2010), based on molecular data using the COI mitochondrial gene sequence, revealed the existence of 57 cryptic species of the 187 species they registered. Some of these species have already been studied clarifying their taxonomy, such as Molpadia violacea Studer, 1876 (O’Loughlin and VandenSpiegel, 2010), or describing new species, such as Taeniogyrus prydzi O’Loughlin & VandenSpiegel, 2010. However, in relation to the genus Trachythyone, the cryptic species found have not been described as new species, or the references were not located to include them in this work.
The results presented constitute a relevant contribution to the “Biogerlache - Antarctic” project and the continued work of the Colombian Expeditions to Antarctica, confirming a high richness of echinoderms in the Gerlache Strait and adjacent areas and generating updated information for the elaboration of the taxonomic inventories of this area and the establishment of a baseline of biological knowledge, which contribute to the definition of possible conservation areas.

