texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink
Echinoderm research in the Colombian Caribbean Sea is very recent. In the 70s they were mainly focused on shallow waters and since the 90s a new era began with research in deep waters (> 200 m depth) with some expeditions carried out entirely by the Marine and Coastal Research Institute ‘‘José Benito Vives de Andréis’’ (Invemar) with the support from several institutions in Colombia and other countries (Borrero-Pérez et al., 2012). As a result of these studies, about 280 species of echinoderms (55 echinoids) are recorded for the Colombian Caribbean, being the third country with the highest echinoderm richness in the Caribbean Sea, overcome by Cuba and Mexico (Alvarado and Solís-Marín, 2013).
The family Echinothuriidae Thomson, 1872, is integrated by seven existing genera, including HygrosomaMortensen, 1903, that mostly inhabit deep waters, considered an important part of deep benthos (Kroh and Mooi, 2021). It is characterized by a thin and flexible test; a series of ambulacral plates completely covering each peristome radius; the oral side with club-shaped or hoof-shaped primary spines; and miliary spines usually provided with poison glands (Mortensen, 1927). Inside this family, only the genus Araeosoma Mortensen, 1903 has been reported for Colombia with the species Araeosoma belli Mortensen, 1903 and Araeosoma fenestratum (Wyville-Thomson, 1872) (Borrero-Pérez et al., 2012).
The genus Hygrosoma presents three species, of which Hygrosoma petersii (A. Agassiz, 1880) is the only one distributed in the Atlantic Ocean. This species has a complex taxonomic history that includes several synonyms and transfers. It was described by Alexander Agassiz under the name of Phormosoma petersii; however, due the short diagnosis presented by the author, there have been misidentifications and, consequently, several reports of their geographic and bathymetric distribution are erroneous (Mortensen, 1927 and 1935). This sea urchin is characterized by the presence of tridentate pedicellariae that has a widened spoon-shaped tip with a straight edge finely toothed or serrated (Mortensen, 1903). Although the shape of pedicellariae is a very stable taxonomic character, it was considered of little morphological value, being rejected as a character of generic importance in this family for several years (Mortensen, 1935); However, many authors have highlighted the importance of using these structures in the identification of sea urchins, especially with similar species that are commonly confused (Mortensen, 1903, 1940; Coppard and Campbell, 2006; Coppard et al., 2010).
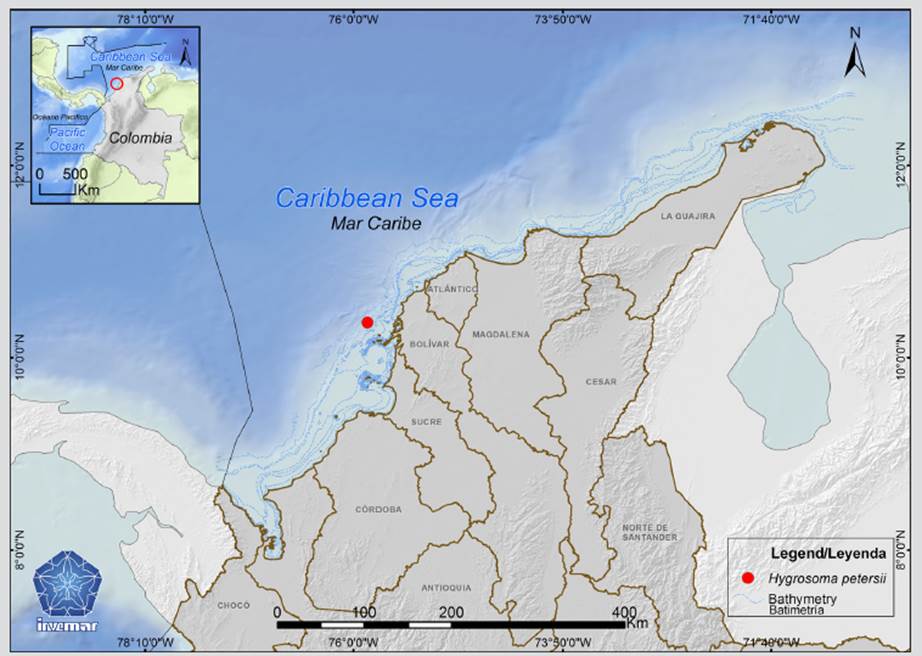
Hygrosoma petersii is widely distributed throughout the North Atlantic. In the present study, this species was found on the continental shelf off Tierra Bomba Island in the Colombian Caribbean. Two organisms were collected at 323 m depth in the RC 4/5/6/7 (E 287) exploration area, using a trawl net during the “Invemar-ANH II” research campaign in 2009, on board the B/I Ancón. There, organisms were relaxed with magnesium sulfate and then fixed in 70 % ethanol (Invemar-ANH, 2010). The specimens were identified under a stereomicroscope following the keys and descriptions of Mortensen (1927, 1935). Digital and SEM photographs of the largest specimen were taken. Specimens are deposited in the echinoderm collection (INV EQU) of the Museum of Natural Marine History of Colombia (MHNMC) Makuriwa at the Invemar. SEM photographs of H. petersii were compared with photographs of their Caribbean congeners Phormosoma placenta (ICML-UNAM 4.126.23) and Araeosoma fenestratum (Thomson, 1872) (ICML-UNAM 4.75.5) from the National Collection of Mexican Echinoderms “Dra. Ma. Elena Caso Muñoz” at the National Autonomous University of México, UNAM.ç
Hygrosoma petersii (A. Agassiz, 1880)
Material examined: Two specimens INV EQU4721, Off Tierra Bomba Island, Colombian Caribbean (10° 21’ 54” N-75° 51’ 19” W) at 323 m depth. Test diameter: 3.6 ˗ 4.7 cm.
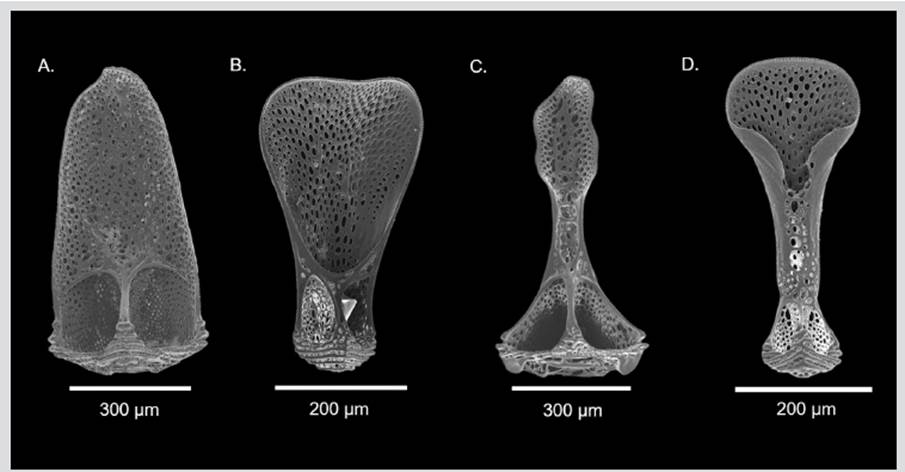
Description: Test very flexible, low and hemispherical, with the oral side almost flat, although slightly rounding towards the ambitus. Interambulacra with large tubercles on the oral side (Figure 1A); on the aboral side, the tubercles are much smaller, scattered, and quite irregularly arranged. Secondary tubercles of the ambitus inconspicuous (Figure 1B). Ambulacra with pore-pairs forming only two series; pore-pair of the primary plate rudimentary, much less developed than that of the secondary plates. Periproctal plates of the apical system well developed, not separated by wide membranous spaces. Genital plates and eye plates well formed, the first ones are very remarkable, although they are not very large; genital pore in the membranous prolongation. These specimens have all the spines broken, but in alive specimens, the larger spines on the peristome and on the proximal part of the oral side are curved at the end, like a hockey stick (Mortensen 1927, 1935). There are large and small tridentate pedicellariae with a neck and curved valves that have a spoon-shaped tip with serrated edge, the smallest have a more rounded tip, while the largest have a long head (Figures 1C˗D). Large tridentate pedicellariae are distributed mainly in the oral area, most of them in the peristome; while the small are distributed throughout the test, occurring in smaller amounts in the aboral zone. Also, there are triphyllous pedicellariae distributed throughout the test, they have curved valves with widened tip, giving the appearance of a spatula with a serrated edge (Figure 1E). The sphaeridia in this species are distributed in pairs, close to tube-feet pores; they are completely round with smooth surface, unlike other echinothurioids that have a very varied shape, often longer with ridges or surface roughness (Figure 1F).
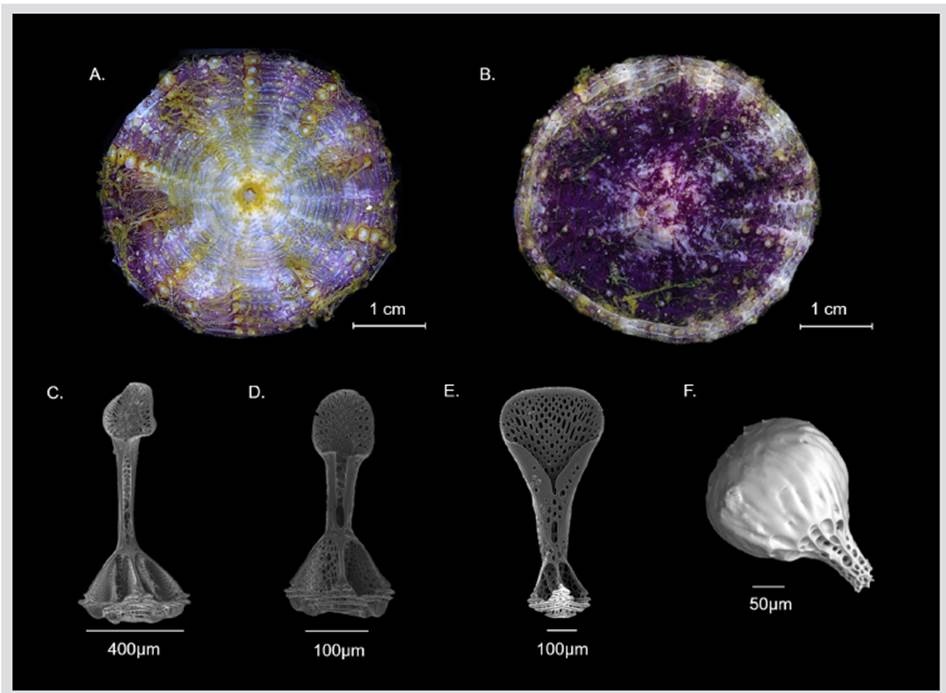
Figure 1 Hygrosoma petersii (INV EQU4721): A. Oral side; B. Aboral side; C. Large tridentate pedicellaria; D. Small tridentate pedicellaria; E. Triphyllous pedicellaria; F. Sphaeridium.
Biology: Hygrosoma petersii lives in soft bottoms (muddy and sandy bottoms) (Grassle et al., 1975; Pawson, 1982). This species was observed alive and described as a “large black urchin with long and thin spines, very active that moves quickly walking on ventral spines with white stout and truncate tips” (Grassle et al., 1975). In relation to feeding habits, the intestinal content of some studied specimens consisted almost exclusively of Sargassum and Thalassia fragments, suggesting an herbivorous diet; however, the occasional presence of foraminifera skeletons and pteropod shells indicate that some of the sediment is ingested (Pawson, 1982).
Distribution: Off Southwest Ireland to the Azores; Senegal; off New Jersey, USA; Mexico; Cuba; Dominican Republic; and Cape Peninsula, South Africa, between 200 to 3700 m depth (Mortensen, 1935; Mooi et al., 2004; Alvarado y Solís-Marín, 2013), also off Tierra Bomba Island, Colombia, at 323 m depth (present paper) (Figure 2).
Comments: Inside the order Echinothurioida, the shape of the oral spines is a very important character that helps to separate the genera within the Echinothuriidae and to separate them from the Phormosomatidae. Species of the genus Araeosoma have oral spines ending in a hyaline helmet with expanded helmet-shaped, while the genus Hygrosoma is characterized by the presence of curved primary spines on the oral side and Phormosoma placenta has oral spines wrapped in a thick “bag” (Mortensen, 1935). However, during offshore sampling, the test of these sea urchins usually collapses being completely flattened, in addition, they are very susceptible to damage within trawl nets, thus the primary spines often break down impeding their correct taxonomic identification (Mortensen, 1927).
Consequently, the pedicellariae and sphaeridia have also been implemented as diagnostic characters of great taxonomic importance, even at a specific level, since they have very characteristic and distinctive shapes for each of the species of Echinothuriidae and Phormosomatidae families (Guerrero-Correa, 2017). Therefore, the echinothurioids of the Colombian Caribbean can be differentiated in the following way: Phormosoma placenta has tridentate pedicellariae with the tip almost as wide as the base, without a neck, and triphyllous pedicellariae spoon-shaped with distal part extended (Figures 3A-B). Inside the Echinothuriidae, H. petersii is characterized in addition to the curved primary spines on the oral side (scattered in the ambitus), by the presence of large areolas, pedicellariae tridentate with a spoon-shaped valves with distal part extended, and triphyllous pedicellariae of elongated leaflets with the base distal part closed in a tube, and the upper section enlarged funnel-shaped (Figures 1C-E). While the genus Araeosoma is distinguished by the curved primary spines with a hyaline white hull end, small areolas, and tridentate, triphyllous, and dactylous pedicellariae. In this case, Araeosoma fenestratum has big tridentate pedicellariae with shorter neck and elongated valves, and bigger triphyllous pedicellariae than H. petersii (Figures 3C-D).