










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.51 no.2 Santa Marta July/Dec. 2022 Epub Dec 14, 2022
https://doi.org/10.25268/bimc.invemar.2022.51.2.1104
ARTICULO DE INVESTIGACIÓN
Estado de Lutjanus synagris en tres áreas protegidas del Caribe colombiano: evaluación basada en datos limitados
1
*
 http://orcid.org/0000-0001-8154-7144
http://orcid.org/0000-0001-8154-7144
2
http://orcid.org/0000-0002-0956-0915
1Parques Nacionales Naturales de Colombia, Dirección Territorial Caribe, Calle 17 No. 4-06, Santa Marta, Colombia. hmartinezviloria@gmail.com
2 Parques Nacionales Naturales de Colombia, Dirección Territorial Caribe, Calle 17 No. 4-06, Santa Marta, Colombia. rebeca.franke@parquesnacionales.gov.co
3 Universidad del Magdalena, Carrera 32 No. 22-08, Santa Marta, Colombia. jnarvaez@unimagdalena.edu.co
4 Universidad del Magdalena, Carrera 32 No. 22-08, Santa Marta, Colombia. jdelahoz@unimagdalena.edu.co
With the purpose of evaluating the status of the lane snapper Lutjanus synagris from limited catch length data from the landings of artisanal fishing catches recorded between 2006 and 2012 in Tayrona and Corales del Rosario y de San Bernardo National Parks, and the Vía Parque Isla de Salamanca, simple indicators and the decision tree methodology were applied. it was observed that this indicator mostly configured a selectivity pattern represented in capture of mayoritary immature specimens (<L mat ) and a spawning biomass level lower than the target reference point, affecting the natural renewal of L. synagris stocks. Also, it was ratified the degree of overfishing for growth and recruitment to which L. synagris is subjected. Reducing pressure on snapper resource is a management objective that requires greater protection in the three protected areas for their ability to export the resource, as well as participatory regional fisheries management for the sustainability of the resource. Therefore, it is recommended to reach conservation agreements that promote responsible fishing of mature fish. This objective will allow maximum biomass yield, and better economic income for fishers. We also propose to apply the methodology described in this study to evaluate other fishery resources caught in the protected areas.
Key words: fishing; overfishing; simple indicators; decision tree
Con el propósito de evaluar el estado del pargo rayado Lutjanus synagris a partir de datos limitados de longitudes provenientes de los desembarcos de las capturas de la pesca artesanal registradas entre 2006 y 2012 en los Parques Nacionales Naturales Tayrona y Corales del Rosario y de San Bernardo y la Vía Parque Isla de Salamanca, se aplicaron indicadores simples y la metodología árbol de decisiones. Se observó un patrón de selectividad en la captura mayoritaria de ejemplares inmaduros (<L mat ) y un nivel de biomasa desovante inferior al punto de referencia límite objetivo, afectando la renovación natural de los stocks de L. synagris. También se ratificó el grado de sobrepesca por crecimiento y reclutamiento. Propender por la disminución de la presión sobre el pargo es un objetivo de manejo que requiere mayor protección en las tres áreas protegidas como zonas fuente, así como un ordenamiento regional participativo para la sostenibilidad del recurso. Por tanto, se recomienda llegar a acuerdos de conservación que propendan por una pesca responsable de peces maduros. Esto, permitirá un máximo rendimiento en biomasa y mejores ingresos económicos para los pescadores. Se propone la metodología descrita para la evaluación de otros recursos aprovechados en las áreas protegidas.
Palabras-clave: pesca; sobrepesca; indicadores simples; árbol de decisiones
INTRODUCCIÓN
Históricamente en las áreas protegidas del Sistema de Parques Nacionales Naturales de Colombia adscritas a la Dirección Territorial Caribe (DTCA) ha existido un constante aprovechamiento de los recursos pesqueros por parte de comunidades asentadas en su interior y de aquellas ubicadas en áreas de influencia, ocasionando fuerte presión sobre las especies explotadas (Manjarrés, 2007; Martínez-Viloria, 2014). Como parte del seguimiento a la actividad pesquera realizado por la DTCA en cinco áreas protegidas con jurisdicción marino-costera, se identificaron aproximadamente 1600 pescadores, de los cuales dependen económicamente unas 4700 personas (Martínez-Viloria et al., 2014). Así mismo, se ha documentado que un gran número de especies aprovechadas por la pesca están fuertemente presionadas, aspecto que podría contribuir negativamente en su condición poblacional (López, 2009; Guzmán, 2010; Jaimes, 2011; Martínez-Viloria et al., 2011).
Entre las especies de mayor aprovechamiento pesquero en los Parques Nacionales Naturales Tayrona-PNNT, Corales del Rosario y de San Bernardo-PNNCRSB y Vía Parque Isla de Salamanca-VIPIS, se encuentra el pargo rayado o chino (Lutjanus synagris) (López, 2009; Guzmán, 2010; Pardo, 2011; Martínez-Viloria et al., 2011; Martínez-Viloria, 2014). Esta es una especie demersal, carnívora generalista y oportunista trófica que se distribuye desde Carolina del Norte (EE. UU.) hasta el sureste de Brasil (Allen, 1985; Cervigón et al.,1992; Doncel y Páramo, 2010), incluyendo el Gran Caribe. Es una importante fuente de ingresos para las pesquerías regionales de países como Bermudas, Cuba, Puerto Rico, Venezuela y Colombia (Luckhurst et al., 2000; Gómez et al., 2001; Claro y Lindeman, 2008; Amorim et al., 2018). Algunos aspectos biológico pesqueros indican que L. synagris puede ser considerada una especie vulnerable a la sobreexplotación en Colombia debido a: (i) las características biológicas de la especie como rápido crecimiento durante el primer año de vida y desaceleración en el crecimiento entre el tercer y cuarto año, madurez sexual a partir de su primer año y temporadas de migraciones de agregación reproductiva (Coleman et al., 2000; Luckhurst et al., 2000; Claro y Lindeman, 2008); (ii) la evidente presión de pesca de la flota artesanal y semindustrial (Mejía y Acero, 2002; Manjarrés, 2004; Barreto y Borda, 2009; De La Hoz et al., 2013, 2017; Altamar et al., 2020); (iii) el aumento del esfuerzo pesquero en la región del Caribe debido a innovaciones técnicas y tecnológicas de las flotas artesanales (Manjarrés, 2004); (iv) la disminución histórica en las capturas tanto por la flota artesanal (Correa y Manjarrés, 2004), como por la industrial de arrastre (Viaña et al., 2004); (v) el deterioro de sus hábitats de reclutamiento, desarrollo y residencia, tales como los arrecifes coralinos, lagunas y ciénagas (Mejía y Acero, 2002); y (vi) la disminución de la edad de madurez en sitios con mayor esfuerzo pesquero (Arteaga et al., 2004).
La presencia de L. synagris en las áreas protegidas está relacionada con la disposición de hábitats estratégicos, donde la especie pasa parte del tiempo en su ciclo natural y cumple funciones de depredador contribuyendo al enriquecimiento orgánico de los arrecifes (Claro y Lindeman, 2008). Por lo tanto, el aprovechamiento no debe contemplar la captura de ejemplares por debajo de la longitud de madurez sexual (Lmat), ni tampoco afectarse la cohorte de los megadesovadores, ya que estos últimos aseguran el vigor genético y una mayor descendencia para garantizar la supervivencia de las poblaciones (Longhurst, 2002; Hsieh et al., 2006).
Bajo este contexto el propósito de esta investigación fue evaluar el estado de L. synagris a partir de datos limitados de longitudes provenientes de los desembarcos de la pesca ejercida en PNNT, PNNCRSB y VIPIS. Esta investigación ayudará a dimensionar el efecto de la pesca sobre el pargo rayado en cada área protegida y a suministrar recomendaciones para el manejo que contribuyan al ordenamiento pesquero en el Caribe colombiano, donde se considere a las áreas protegidas como sitios estratégicos que proporcionan condiciones favorables para que las especies pesqueras se mantengan y puedan ser aprovechadas por fuera (Roberts et al., 2001; Di Lorenzo et al., 2016).
ÁREA DE ESTUDIO
El trabajo se realizó en las áreas protegidas PNNT, PNN CRSB y la VIPIS (Figura 1). El PNNT está localizado al nororiente de Santa Marta, departamento del Magdalena (11° 16’ 20” N y 74° 12’ 56” W a 11° 21’ 33” N y 73° 53’ 11” W) (PNNC, 2006). Cuenta 15 000 ha, de las cuales 3 000 hacen parte del área marina. En esta área más de 200 pescadores hacen uso con diferentes artes y/o métodos de pesca (Martínez-Viloria, 2009; Martínez-Viloria et al.,2014). El conflicto de uso por pesca ha sido histórico y la Sentencia T-606 de 2015 de la Corte Constitucional colombiana indica que no se pesque en áreas protegidas, a menos que sea en el marco de la subsistencia.
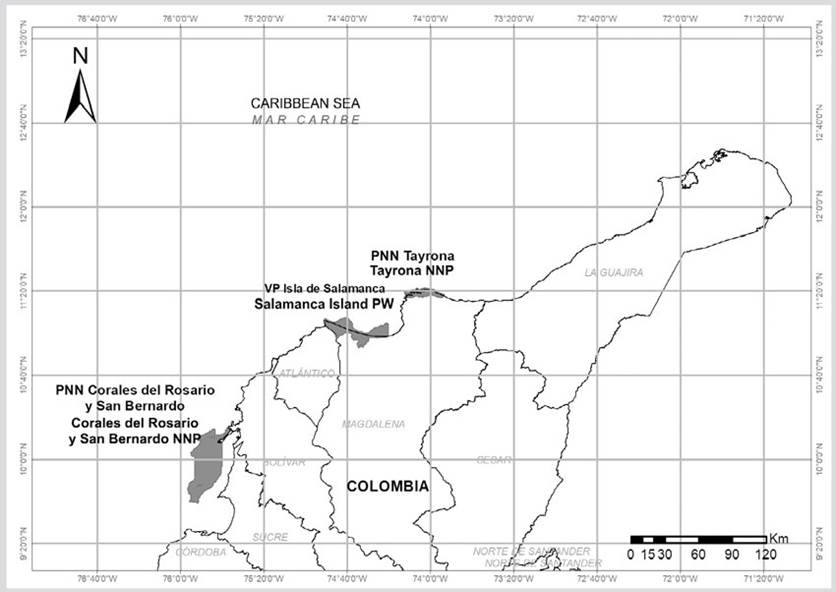
Figura 1 Ubicación de áreas protegidas de la Dirección Territorial Caribe Parque Nacional Natural Tayrona (PNNT), Vía Parque Isla de Salamanca (VIPIS) y PNN Corales del Rosario y de San Bernardo (PNNCRSB).
El PNN CRSB está ubicado a 45 km del suroccidente de la ciudad de Cartagena de Indias y a 30 km al noroccidente de la localidad de Tolú entre los departamentos de Bolívar y Sucre (10º 15’ - 9º 35’ N y 75º 47’ - 75º 50’ W) (PNNC, 2020). De las 120 000 ha de esta área protegida, 118 800 ha (99 %) son marinas. Donde más de 600 pescadores hacen uso de los recursos a través de la pesca (Martínez-Viloria et al., 2011).
La VIPIS se encuentra localizada en el sector costero del departamento del Magdalena, en jurisdicción de los municipios de Pueblo Viejo y Sitio Nuevo (11° 7’ 19” -10° 53’ 07” N y 74° 20’ 34” - 74° 51’ 00” O) (PNNC, 2017). De las 56 200 ha, en esta área protegida 28 200 son marinas. En donde más de 350 pescadores realizan actividades de pesca (Martínez-Viloria, 2009, 2014), utilizando redes de arrastre (Cochrane, 2005).
MATERIALES Y MÉTODOS
Descripción de artes y/o métodos de pesca
Línea de mano. Es un arte de pesca empleado para la captura de especies demersales a través de diferentes métodos, supeditado por el movimiento de la embarcación (Aunap-Unimagdalena, 2014). En las áreas protegidas la línea de mano es el arte de mayor uso (34 %; Martínez-Viloria et al., 2014). Consta de un cordel de nylon tipo monofilamento con resistencias de 3 a 120 kgf, dotado de uno a seis anzuelos en su mayoría tipo jota “J” por línea, que oscilan entre los números (N°) J6 y J12. En el PNNT es empleada como línea madre o principal el nylon de 35 a 120 kgf con un largo que puede oscilar entre 90 a 200 m y anzuelos tipo “J” N° 7, 9 y 10. En el PNNCRSB, se presentan similares características a las del PNNT, siendo además el uso de anzuelos tipo circular o curvos N° C5 y C6.
Red de enmalle fija. Representan 23 % de los artes utilizados en las áreas protegidas (Martínez-Viloria et al., 2014). Consta de uno o varios paños de red que en su mayoría son de monofilamento. Las utilizadas en PNNT registran longitudes hasta 225 m, altura máxima de 17 m y tamaños de malla que oscilan entre 8,89 y 11,43 cm (3,5 y 4,5 pulgadas). Para el caso de PNNCRSB los tamaños de malla oscilan entre 5,08 y 8,89 cm (2,0 y 3,5 pulgadas); mientras que en VIPIS varían entre 5,08 y 7,62 cm (2 y 3 pulgadas).
Buceo. Es un método practicado en el PNNT y en el PNNCRSB, a profundidades entre 3 y 20 m, con arpón para la captura de peces, o sin arpón para capturar caracoles (Eustrombus gigas, Melongena melongena, Cittarium pica), langosta (Panulirus argus), pulpo (Octopus spp.); otras veces se emplea con varilla, ganchos y lazos para la captura de langosta y pulpo.
Palangre. Están constituidos por un cordel principal o línea madre horizontal de nylon monofilamento o multifilamento de 60 a 120 kgf en PNNT y de 20 a 60 kgf en VIPIS, en el cual van unidas líneas secundarias (reinales) a distancias regulares (~1,2 m) en VIPIS y (~ 3,6 m) en PNNT, en cuyos extremos van anzuelos tipo jota “J” que oscilan entre N° 9, 10 y 12 en VIPIS y 7, 8 y 10 en PNNT. Se utilizan con mayor frecuencia en VIPIS hasta 20 m de profundidad y en el PNNT hasta 200 m de profundidad y por lo general oscilan entre 100 y 300 anzuelos para la pesca de peces demersales.
Registro de datos y análisis de información
En el marco de la caracterización de la presión por pesca en las áreas protegidas, se hicieron registros mensuales de la longitud total (cm) de los peces desembarcados entre 2006 y 2012 en el PNNCRSB y la VIPIS, y de 2006 a 2010 en el PNNT; los pargos rayados fueron medidos con un ictiómetro de 0,1 cm de precisión y agrupados en marcas de clase de 0,5 cm. Estos registros de L. synagris fueron discriminados por área protegida, por arte y/o método de pesca y por año, empleando un esfuerzo de muestreo que en todas las áreas fue superior a 30 % de las Unidades Económicas de Pesca activas (metodología Sistema de Información Pesquera del Invemar-Sipein Ver. 3.0, Narváez et al., 2005).
Puntos de referencia biológicos
El análisis de datos fue basado en los siguientes puntos de referencia biológicos (PRB): i) longitud de madurez sexual-longitud donde 50 % de los peces de una población determinada llegan a estar sexualmente maduros por primera vez (Lmat; Froese y Binohlan, 2000; Fontoura et al., 2009; Cope y Punt, 2009), ii) longitud óptima de captura-equivale a la longitud en la que una cohorte no pescada alcanza su máxima tasa de crecimiento y biomasa (Lopt ;Froese et al., 2008); y la longitud de megadesovadores (Lmega)-representa a los ejemplares que en las capturas equivalen a aquellos peces ubicados por encima de 10 % de Lopt (Froese, 2004). También, se utilizó como punto de referencia límite el 0,25 de la biomasa desovante-SB (Cope y Punt, 2009).
Para esta investigación se consideró que en cada área protegida existe una población genéticamente diferenciada de L. synagris (Fst > 0,061, p < 0,05; Narváez et al., 2015). Por lo tanto, se asumieron valores de Lmat independientes, obtenidos de investigaciones previas realizadas en cada área protegida y correspondientes a sexos combinados (PNNT = 33,08 cm, Arteaga et al., 2004; VIPIS = 30,6 cm, Pardo, 2011; PNNCRSB = 24,6 cm, Jaimes, 2011). Los valores de Lopt y Lmega fueron calculados siguiendo las instrucciones de Froese y Binohlan (2000) y Froese et al. (2008).
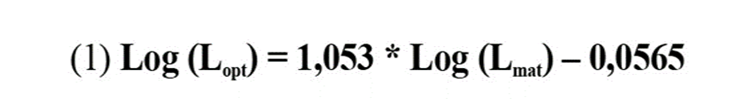
Donde, Lopt es la longitud óptima de captura y Lmat es la longitud de madurez sexual.
Indicadores basados en longitudes de captura de L. synagris
Para evaluar el estado de las poblaciones de L. synagris se utilizaron los indicadores sugeridos por Cope y Punt, (2009): i ) P mat, correspondiente a la proporción de individuos de L. synagris que son capturados por encima de Lmat; ii ) P opt, proporción determinada a partir de ejemplares capturados entre 0,9 y 1,1 de Lopt, iii ) P mega, proporción de ejemplares capturados por encima de Lmega, iv ) P obj, equivalente a la sumatoria de las proporciones de longitudes.
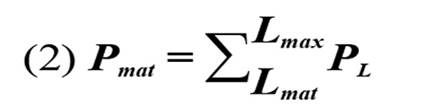
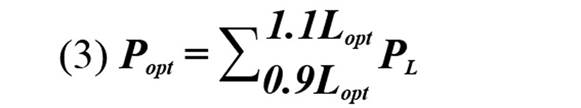
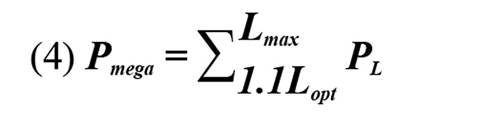
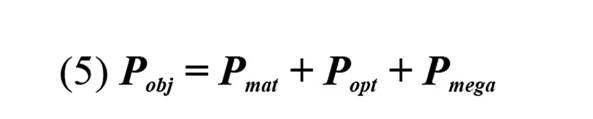
Con el fin de determinar las tendencias del impacto de la actividad pesquera sobre L. synagris se realizó un análisis de regresión con los datos de proporciones de individuos calculada por debajo de Lmat, entre Lmat y Lmega y por encima de Lmega. Para entender si la pesca está generando riesgo de sobrepesca en el pargo rayado se aplicó la metodología de “árbol de decisiones” propuesta por Cope y Punt (2009) la cual indica si la biomasa desovante (SB) de una especie está por encima o por debajo de un punto de referencia límite equivalente a 0,25 SB. El resultado de Pobj se utilizó en conjunto con Popt + Pmega o el valor de Popt para luego determinar el patrón de selectividad como uno de los siguientes: 1) Captura de peces pequeños e inmaduros; 2) Captura de peces pequeños y de tamaño óptimo; 3) Curva de selectividad similar a la ojiva de madurez; 4) Captura de peces de tamaño óptimo y mayor; o 5) Captura de peces de tamaño óptimo.
Dependiendo del tipo de selectividad, se calculó la relación entre la longitud en madurez (Lmat) y la longitud óptima de captura (Lopt). Esto se usó junto con Pmat o Popt para inferir si la biomasa desovante (SB) estaba en o por encima del punto de referencia objetivo (RP). Si el valor de SB estaba por debajo del RP se infirió que la especie está sobreexplotada.
Este análisis complementario, ofrece una visión diferente del estado de la especie a partir de los indicadores de Froese solamente. Si la pesquería captura peces pequeños e inmaduros (selectividad tipo 1), el punto de referencia para Pmat es > 0,25 en el árbol de decisión. La recomendación de Froese (2004), sin embargo, es mucho más conservadora al sugerir que Pmat debe ser igual a cero permitiendo que más peces desoven.
Además se tuvo en cuenta las recomendaciones de sostenibilidad de Froese (2004), las cuales indican que: 1) valores de Pobj menor que uno (< 1) es indicativo de patrones de selectividad que no siguen las recomendaciones de sostenibilidad; 2) valores de Pobj mayor que uno (> 1) siguen los patrones de sostenibilidad; 3) valores de Pobj entre uno y dos (1 y 2) indica que se distinguen patrones de selectividad que contienen algunos peces inmaduros y de tamaños cercanos a la Lopt; 4) valores de Pobj mayor que dos (> 2) indica que se cumplen los patrones de sostenibilidad; y 5) finalmente, si Pobj es menor que uno (< 1) y si Popt + Pmega = 0, indica que solo se pesca individuos inmaduros, por lo que se considera altamente indeseable bajo las recomendaciones de sostenibilidad de Froese (2004).
También se calculó la longitud media de captura (LMC) acumulada, discriminada anualmente por arte y/ó método de pesca por área protegida. Con el fin de determinar diferencias estadísticas de la LMC entre las áreas protegidas y año en cada arte y/ó método de pesca, se realizó un análisis de varianza (Anova) no paramétrico de Kruskal-Wallis debido a que los datos no cumplieron los supuestos de normalidad de los residuos y homogeneidad de las varianzas. Se consideró como variable dependiente la LMC y como factores las áreas protegidas y los artes, con un nivel de significancia de 0,05. En el caso que las pruebas resultaran significativas, se aplicó la comparación a través de un diagrama de cajas y bigotes con muescas. Desde el punto de vista gráfico, la LMC se contrastó con la Lmat, Lopt y Lmega para complementar la evaluación del impacto relativo de la pesca sobre L. synagris.
RESULTADOS
Un total de 33 174 registros de longitud total de L. synagris provenientes del seguimiento a la presión por pesca fueron obtenidos en PNNT, PNNCRSB y VIPIS. El 53,2 % correspondió a registros de los pescadores del sector marino de VIPIS con palangres y redes de enmalle, 40,3 % a registros de pescadores de línea mano, red de enmalle y buceo en el PNNCRSB y 6,5 % observados con línea de mano y red de enmalle en el PNNT.
Al contrastar los resultados de LMC con los puntos de referencia en las tres áreas protegidas, se pudo observar que en todos los casos estuvo por debajo de Lmat, Lopt y Lmega, siendo VIPIS donde los valores estuvieron muy por debajo (Figura 2). Por otro lado, las LMC de L. synagris variaron significativamente entre áreas protegidas (basado en las medianas, Kruskal-Wallis: H = 5026; p < 2,2 × 10-16) (Figura 2). Además, se observó que PNNCRSB fue donde se capturó en promedio los peces más pequeños (25,06 cm) y en PNNT los más grandes (34,2 cm) (Figura 2).
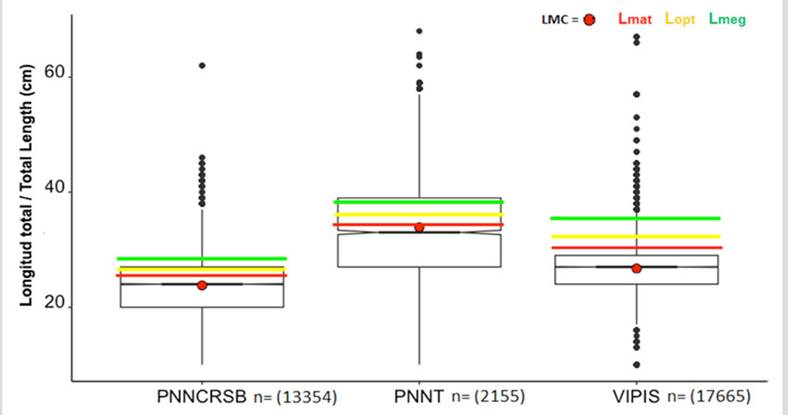
Figura 2 Longitud media de captura (LMC) y diagrama de cajas y bigotes con muescas a partir de datos de L. synagris capturado con todos los artes y contrastada con Lmat, Lopt y Lmega en PNNT, VIPIS y PNNCRSB. PNNCRSB: Lmat = 24,6 cm, Lopt = 25,6 cm y Lmega = 28,2 cm; PNNT: Lmat = 33,08 cm, Lopt = 35,5 cm y Lmega = 38,5 cm; VIPIS: Lmat = 30,6 cm, Lopt = 32,2 cm y Lmega = 35,4 cm.
Efecto de los artes de pesca sobre L. synagris por área protegida
Parque Nacional Natural Tayrona. Al analizar las fracciones de captura de L. synagris, se observó que las tallas más pequeñas que se registraron tanto con línea de mano, como con red de enmalle fija en el PNNT fueron de 10 cm de LT y los más grandes de 68 cm. 50,6 % de los peces capturados con línea de mano estuvieron por debajo de Lmat, 16,7 % fueron capturados entre Lmat y Lmega, y 32,7 % por encima de Lmega. Con red de enmalle, 27 % de los peces fueron capturados por debajo de Lmat, 53,5 % entre Lmat y Lmega y 19,5 %, en longitudes superiores a Lmega (Figura 3). Con relación a LMC, se observó que existe un comportamiento similar en la tendencia de ambos artes de pesca, con una variación interanual significativa de LMC (Kruskal-Wallis: H = 223, p < 2,2 × 10-16 para línea de mano; H = 50,6, p = 5,57 × 10-11 para red de enmalle). El año 2009 fue diferente a los demás en el caso de línea de mano, mientras que para red de enmalle 2006 y 2007 fueron estadísticamente diferentes a 2008 y 2009. No obstante, entre estos últimos años no se detectó diferencias (Figura 3).
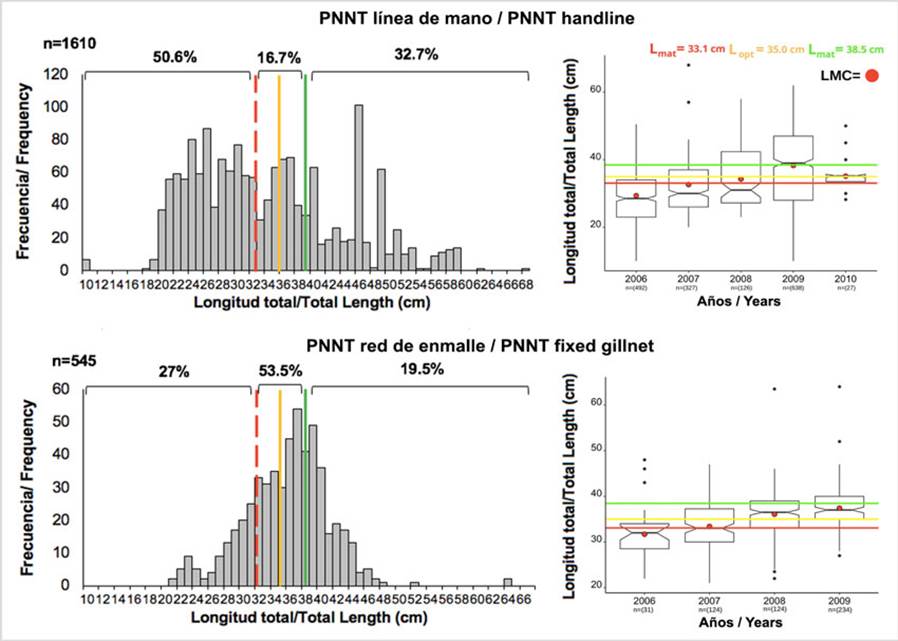
Figura 3 Histograma de frecuencia de longitudes y diagrama de cajas y bigotes con muescas a partir datos de L. synagris capturados con línea de mano y red de enmalle fija entre 2006 a 2009 en el PNNT y LMC anual contrastada con Lmat (33,08 cm), Lopt (35,5 cm) y Lmega (38,5 cm); n indica el tamaño de la muestra.
En todos los casos las regresiones fueron estadísticamente significativas (p < 0,05), con una tendencia hacia la disminución de la captura de individuos inmaduros (Figura 4a); mientras que en la fracción de maduros (Lmat y Lmega), para ambos artes la tendencia fue hacia el aumento (Figura 4b). Con respecto a Lmega (megadesovadores) se observó una tendencia muy leve hacia el aumento de longitudes por encima de este indicador para ambos artes, siendo más pronunciado con red de enmalle (Figura 4c).
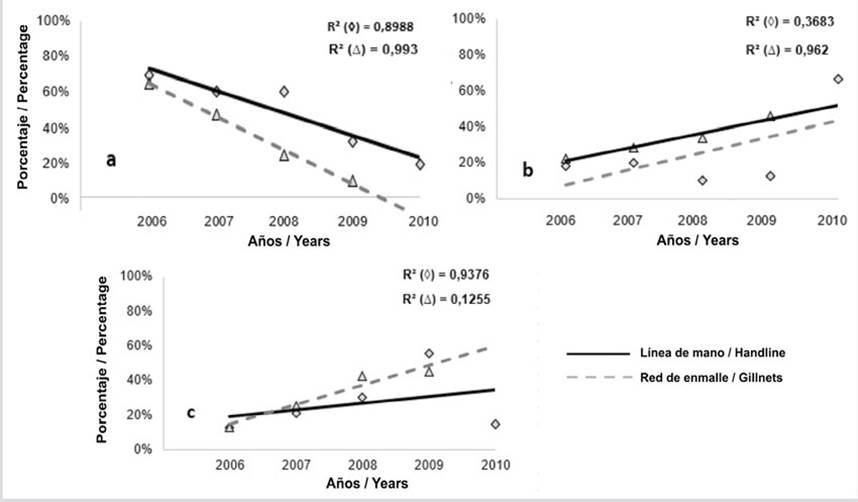
Figura 4 Tendencia de proporciones de longitudes de L. synagris capturados en el PNNT con línea de mano (◊) y red de enmalle (Δ): a) por debajo de la longitud de madurez sexual (Lmat, inmaduros), b) entre la longitud de madurez sexual (Lmat) y la longitud de megadesovadores (Lmega) (maduros) y c) por encima de Lmega (megadesovadores). R2 corresponde al coeficiente de determinación.
Parque Nacional Natural Corales del Rosario y de San Bernardo. 54 % de los individuos capturados con línea de mano estuvieron por debajo de Lmat, 28 % entre Lmat y Lmega y 18 % por encima de Lmega. Con red de enmalle los porcentajes más altos correspondieron a ejemplares capturados entre Lmat y Lmega (42,5 %) y por encima de Lmega (39 %). Así mismo, 35 % de los individuos capturados con buceo estuvieron por debajo de Lmat, 40 % entre Lmat y Lmega y 25 % por encima de Lmega (Figura 5). La LMC calculada por año y correspondiente a la línea de mano en todos los años estuvo por debajo de los puntos de referencia Lmat, Lopt y Lmega, con una variación entre años significativa (Kruskal-Wallis: H = 1085; p < 2,2 × 10-16). En 2006, 2007 y 2008 difirieron significativamente, mientras que 2009 a 2012 fueron estadísticamente iguales (Figura 5). Para red de enmalle se presentó variación entre años (Kruskal-Wallis: H = 107,8; p < 2,2 × 10-16) y se observó que 2010 fue diferente a todos los años, excepto con 2012 (Figura 5). En cuanto al buceo, se observó una variación interanual significativa (Kruskal-Wallis: H = 37,56; p = 3,69 × 10-5); se resalta que por los tamaños de las muestras la comparación sólo fue posible entre 2008, 2009 y 2011. Así mismo, se detectó que 2009 es diferente a 2008 y 2011 (Figura 5). En los tres años, LMC estuvo por debajo de Lmega y sólo en 2009 igualó el valor de Lopt, mientras que en 2009 y 2011 los valores estuvieron ligeramente por encima de Lmat. (Figura 5).
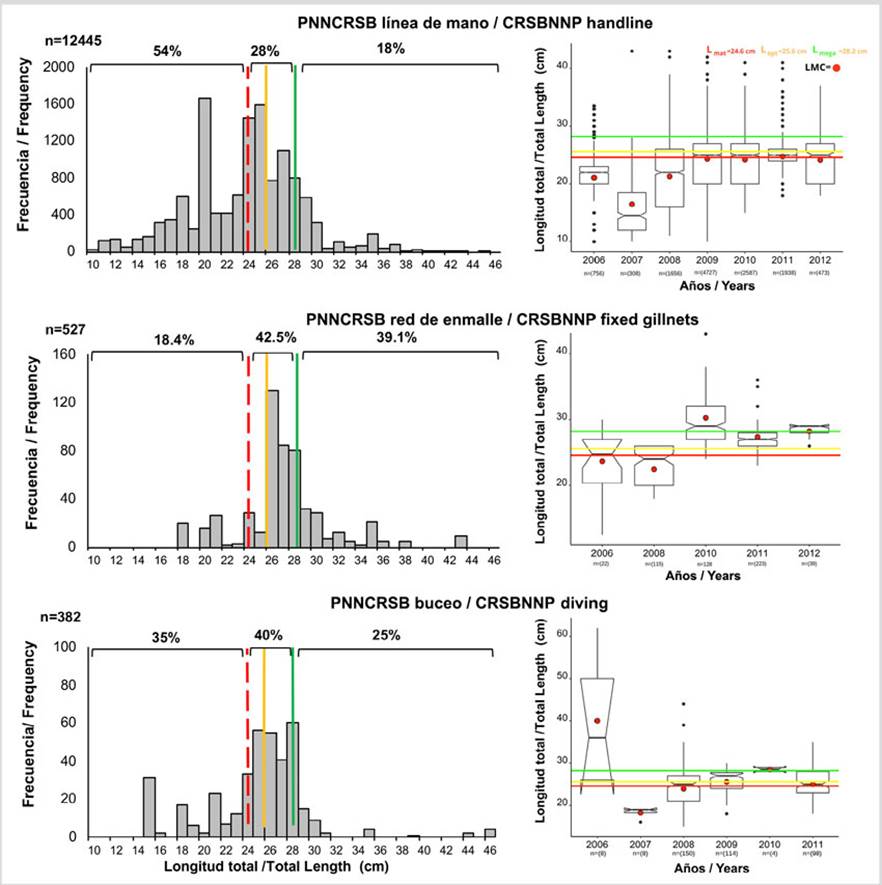
Figura 5 Histograma de frecuencia de longitudes y diagrama de cajas y bigotes con muescas a partir de datos de L. synagris capturados con línea de mano, red de enmalle fija y buceo desde 2006 a 2012 en PNNCRSB y LMC anual contrastada con Lmat (24,6 cm), Lopt (25,6 cm) y Lmega (28,2 cm); n indica el tamaño de la muestra.
En la mayoría de los casos las regresiones fueron estadísticamente significativas (p < 0,05), excepto el buceo (p > 0,05). Al igual que en PNNT, la tendencia fue hacia la disminución de la captura de individuos inmaduros (Figura 6a), mientras que en la fracción de maduros (Lmat y Lmega) la tendencia fue hacia el aumento, excepto buceo (Figura 6b). Con relación a Lmega (megadesovadores) la tendencia fue hacia el aumento para los tres artes, siendo más pronunciado con red de enmalle (Figura 6c).
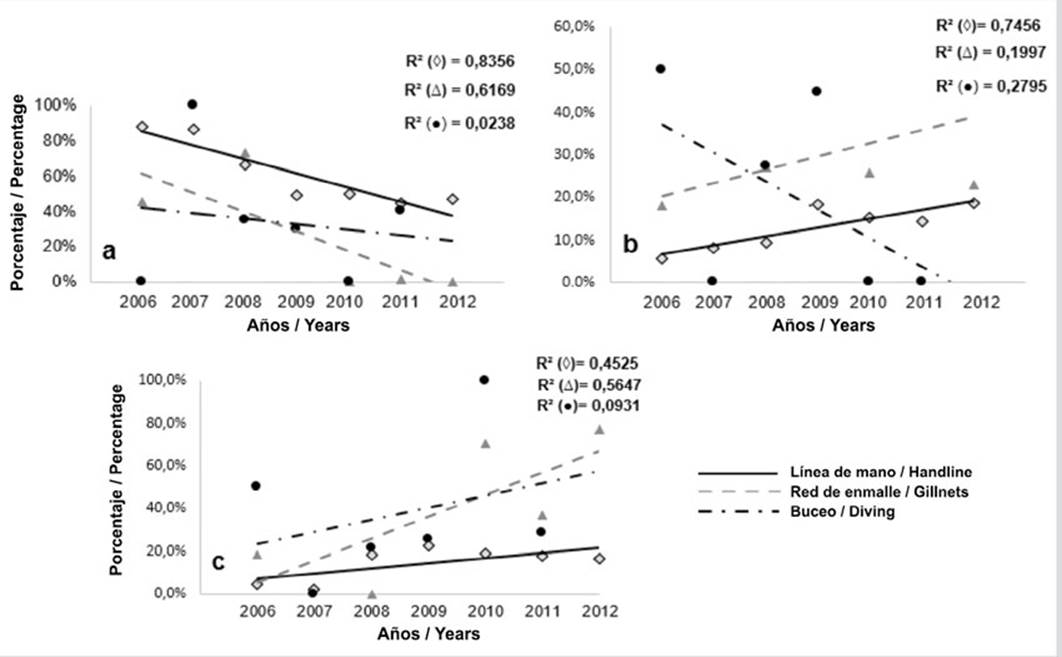
Figura 6 Tendencia de proporciones de longitudes de L. synagris capturados en PNN CRSB con línea de mano (◊), red de enmalle (Δ) y buceo (●): a) por debajo de la longitud de madurez sexual (Lmat) (inmaduros), b) entre la longitud Lmat y Lmega (maduros) y c) por encima de Lmega (megadesovadores). R2 corresponde al coeficiente de determinación.
Vía Parque Isla de Salamanca. 83,4 % de los ejemplares capturados con palangre estuvieron por debajo de Lmat, 15,4 % entre Lmat y Lmega; y 1,3 % estuvo por encima de Lmega. Con respecto a la red de enmalle, 95,4 % de los individuos fueron pescados sin alcanzar la Lmat (Figura 7). El comportamiento de la LMC con palangre presentó una variación interanual significativa (Kruskal-Wallis: H = 2783; p < 2,2 × 10-16), observándose que hubo diferencia entre todos los años, excepto entre 2006 y 2007. La red de enmalle presentó variación interanual significativa (Kruskal-Wallis: H = 464,4; p < 2,2 × 10-16), observándose que en todos los años hubo diferencia, excepto entre 2010 y 2011 (Figura 7). También se encontró que, durante todos los años, en ambos artes de pesca la captura de peces estuvo por debajo de Lmat y, así mismo, por debajo de los otros indicadores (Figura 7).
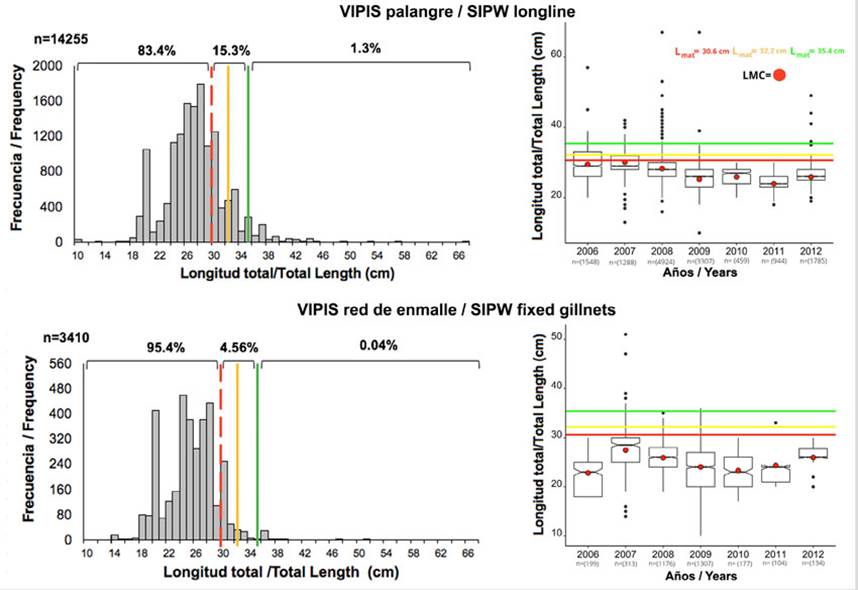
Figura 7 Histograma de frecuencia de longitudes y diagrama de cajas y bigotes con muescas a partir de datos de L. synagris capturados con palangre y red de enmalle fija acumulada desde 2006 a 2012 en VIPIS y LMC anual contrastada con Lmat (30,6 cm), Lopt (32,2 cm) y Lmega (35,4 cm);n indica el tamaño de la muestra.
En la mayoría de los casos las regresiones fueron estadísticamente significativas (p < 0,05), excepto con red de enmalle (p > 0,05). Se observó una tendencia hacía el aumento de la captura de individuos inmaduros con palangre, mientras que para red de enmalle el comportamiento fue constante y cercano a 100 % (Figura 8a). La tendencia de la proporción de maduros (Lmat y Lmega) y de Lmega (megadesovadores) fue similar en la captura con palangre y red de enmalle (Figura 8b y c).
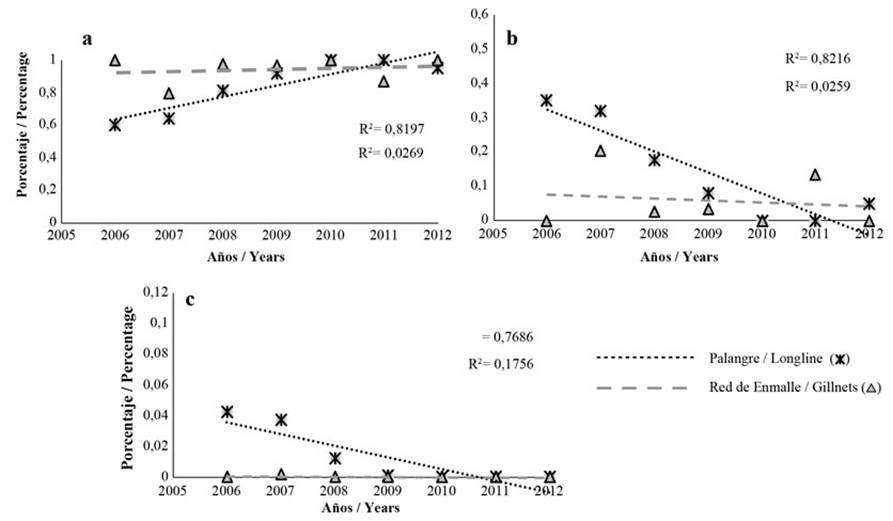
Figura 8 Tendencias de proporciones de longitudes de L. synagris capturados en VIPIS con palangre (*) y red de enmalle (Δ): a) por debajo de la longitud de madurez sexual (Lmat) (inmaduros), b) entre la longitud de madurez sexual (Lmat) y por encima de la longitud de megadesovadores (Lmega) (maduros) y c) por encima de Lmega (megadesovadores). R2 corresponde al coeficiente de determinación.
Aplicación del árbol de decisiones de Cope y Punt (2009)
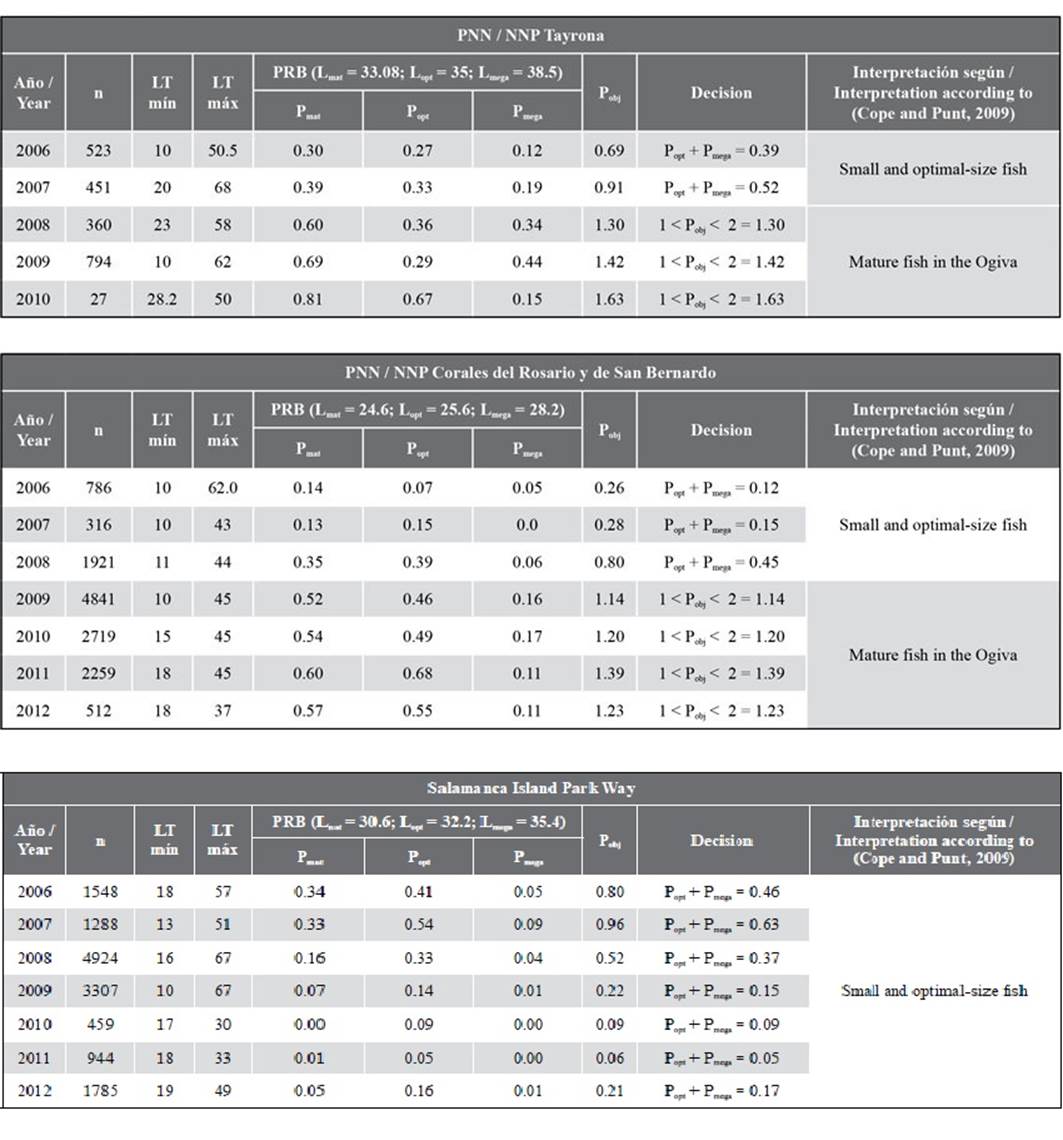
Usando el árbol de decisiones de Cope y Punt, (2009), se observó que en PNNT los dos primeros años evaluados (2006 y 2007) mostraron selectividad tipo 2 (Pobj< 1, pero donde Popt+ Pmega> 0), dado que en estos años la relación Lmat: Lopt y el valor Pmat > 0,25 sugieren que la biomasa desovante fue mayor que el punto de referencia objetivo. No obstante, para los años restantes se evidenció selectividad tipo 3 (donde Pobj estaba entre 1 y 2), en el que la relación Lmat: Lopt fue superior a 0,9; sin embargo, la Pmat fue menor que 0,9, sugiriendo que la biomasa desovante fue menor que el punto de referencia objetivo evidenciando signos de sobrepesca.
Con respecto a PNNCRSB, los tres primeros años mostraron selectividad tipo 2 (Pobj< 1, pero donde Popt+ Pmega> 0), mientras que los cuatro restantes mostraron selectividad tipo 3 (donde Pobj estaba entre 1 y 2). En estos últimos cuatro años, la relación Lmat: Lopt y el valor de Pmat0,9 sugieren que la biomasa desovante era menor que el punto de referencia objetivo y por lo tanto se observaron signos de sobrepesca. Sólo en 2008 se observó una relación Lmat: Lopt y Pmat > 0,25 indicando que la biomasa desovante fue mayor que el punto de referencia objetivo y en consecuencia no se presentaron signos de sobrepesca.
En VIPIS para todos los años el tipo de selectividad fue de tipo 2 (Pobj< 1, y Popt + Pmega > 0); con una relación Lmat: Lopt y Pmat < 0,25 indicando sobrepesca en el período 2008 a 2012; sin embargo, de los siete años evaluados sólo en 2006 y 2007 se observó una relación Lmat: Lopt y un valor Pmat que sugirió que la biomasa desovante fue mayor que el punto de referencia objetivo (Tabla 1).
Tabla 1. Proporciones de longitudes de L. synagris calculadas a partir de los puntos de referencia biológicos-PRB por área protegida y año (los ítems de decisión e interpretación corresponden a lo sugerido por Cope y Punt, 2009). n: número de individuos; LT: longitud total; mín: longitud mínima; máx: longitud máxima.
Tabla 1 Proporciones de longitudes de L. synagris calculadas a partir de los puntos de referencia biológicos-PRB por área protegida y año (los ítems de decisión e interpretación corresponden a lo sugerido por Cope y Punt, 2009). n: número de individuos; LT: longitud total; mín: longitud mínima; máx: longitud máxima.
DISCUSIÓN
A nivel mundial, se ha documentado que 58,1 % de las poblaciones de peces alcanzó su aprovechamiento máximo sostenible y 31 % se encuentran sobrepescadas; además, en los últimos 15 años los stocks pesqueros sobrepescados aumentaron en 20 % (FAO, 2018). Este comportamiento también se ha reflejado en las pesquerías costeras del Caribe colombiano, en las que se ha observado niveles máximos de aprovechamiento y sobrepesca por crecimiento y reclutamiento (Martínez-Viloria et al., 2006; García et al., 2007; Narváez et al., 2008; Barreto y Borda, 2009). Esta situación no es ajena a las áreas protegidas, ya que en el presente estudio se evidenció sobrepesca de crecimiento y reclutamiento en las poblaciones de L. synagris aprovechadas en PNNCRSB, PNNT y VIPIS.
Variación de la longitud media de captura-LMC y contraste con los puntos de referencia
Si bien la LMC de L. synagris estuvo por debajo en las tres áreas protegidas, fue en el PNNCRSB donde se capturaron en promedio los peces más pequeños, situación que también ha sido documentada por Guzmán (2009), Jaimes (2011) y Martínez-Viloria et al. (2011). Así mismo, se observó que en el PNNT la LMC estuvo por debajo de Lmat y por debajo de Lopt y Lmega, lo cual indica la magnitud del impacto que se está generando sobre la especie (López, 2009). Bajo este panorama, se puede inferir que el pargo rayado ha sido aprovechado de manera insostenible en las tres áreas protegidas, lo cual agrava la situación de la especie ya que la forma de aprovechamiento por los pescadores es común en el Caribe colombiano y ha sido empleada por décadas (Manjarrés, 1993; Barreto y Borda, 2009; López, 2009; De La Hoz et al., 2013, 2017; Narváez et al., 2013)
Efecto de los artes de pesca por área protegida
Dado que en el PNNT se evidenció que los artes de pesca línea de mano y red de enmalle concentran su captura mayoritaria en individuos inmaduros y maduros, se requiere que en el área protegida se fortalezca la función de protección a la estructura de tamaños de la especie (López, 2009). Este asunto genera preocupación debido a que algunos estudios han documentado que la situación de los stocks de L. synagris aprovechados en el Caribe de Colombia están sobrepescados y en el mejor de los casos cerca al límite de su explotación (Manjarrés, 2004; Manjarrés et al., 2004; Barreto y Borda, 2009; López, 2009; De La Hoz et al., 2013, 2017; Narváez et al., 2013).
Para el PNNCRSB se observó que el arte de pesca línea de mano tiene mayor incidencia sobre la captura de individuos pequeños e inmaduros en proporciones que superan 53 % de los ejemplares, mientras que la red de enmalle y el método buceo dirigen su esfuerzo a la extracción de ejemplares más grandes, lo cual sugiere la importancia de desarrollar estrategias para disminuir la presión acorde con la normatividad establecida para PNNC. En un escenario favorable para la especie esas estrategias podrían orientarse a que sólo se permita la captura de peces ubicados entre Lmat y Lmeg (maduros), en cuya fracción se obtiene el máximo rendimiento en biomasa y a su vez los mejores ingresos económicos para los pescadores (Froese et al., 2008, 2016).
En la VIPIS se evidenció que los artes de pesca palangre y red de enmalle dirigen su esfuerzo a la captura de individuos inmaduros en proporciones que supera 83 %, lo cual configuró que la LMC en todos los casos estuviera por debajo de los puntos de referencia Lmat, Lopt y Lmega. Esta situación es el reflejo de un patrón de selectividad basado en la captura mayoritaria de peces pequeños (Froese et al., 2004)
A pesar de lo anterior y el esfuerzo de PNNC por alcanzar los objetivos de conservación de las áreas protegidas, sus ecosistemas y especies, se convierte en un reto el manejo de la presión que ejerce la pesca, ya que fue evidente que los artes de pesca están seleccionando peces pequeños (inmaduros). Situación relacionada con el hecho de que los pescadores han diseñado sus artes de pesca con anzuelos y tamaños de malla pequeños, lo cual también ha sido registrado por López (2009) y Jaimes (2011). Todo lo anterior, explica por qué en PNNT se capturan peces más grandes que en PNNCRSB y VIPIS. Además, el stock de L. synagris del PNNT está representado por peces de mayor tamaño en el Caribe norte colombiano (Gómez-Canchong et al., 2004). Lo preocupante de la situación descrita es que los pescadores artesanales de la región tienden a cambiar las características de las artes de pesca para aumentar las capturas (Martínez -Viloria et al., 2006; Narváez et al.,2008). Para el caso de PNNT, en la década de los noventa del siglo pasado se documentó que las redes de enmalle disminuyeron los tamaños de malla, pasando de 7,62-19,32 cm (3-7,6 pulgadas) en 1993 a 7,62-10,16 cm (3-4 pulgadas) en 1999 (López, 2009). Es claro que estos cambios traen como consecuencia efecto de selectividad sobre los tamaños de la especie objetivo (Pope et al., 1983; Altamar et al., 2020).
En cuanto al impacto de los artes de pesca en el tiempo, a través de los porcentajes de captura de longitudes, se observó que para la línea de mano y red de enmalle en el PNNT y el PNNCRSB existe una tendencia hacia la disminución de las capturas de ejemplares inmaduros y una tendencia en aumento de la captura de individuos maduros y megadesovadores mucho más marcada con red de enmalle. Esta situación llama la atención y sugiere la necesidad de revisar el impacto que se está generando sobre los individuos megadesovadores, ya que los porcentajes de longitudes en las capturas (hasta 56 %) muestra señales que indican que la parte superior de la estructura de tallas puede estar erosionada como consecuencia de la remoción de los individuos más grandes, aspecto que afectaría la capacidad de resiliencia de la población (Longhurst, 2002; López, 2009). En el PNNCRSB, la tendencia de la red de enmalle y el método buceo es más marcada hacia la captura de ejemplares maduros y megadesovadores, en porcentajes que llegan a ser 50 % con buceo y superior a 75 % con red de enmalle. Esta situación sugiere que una evaluación más profunda sobre los megadesovadores es necesaria, ya que se puede estar comprometiendo la alta fecundidad y transferencia genética que los megadesovadores aportan al ecosistema (Froese y Binohlan, 2000; Bent-Hooker, 2006). Así mismo, se observó que la línea de mano es el arte que más captura ejemplares inmaduros, aspecto que debe ser revisado ya que se podría afectar la renovación del stock que utiliza las condiciones que ofrece el área protegida (Jaimes, 2011; Martínez-Viloria et al., 2011). Todo lo anterior, contrasta con lo observado en la VIPIS donde la captura de ejemplares inmaduros con los artes de pesca palangre y red de enmalle, evidencia el grado de sobrepesca por crecimiento, lo cual deja ver que ha sido continuo en el tiempo y coincide con Pardo (2011).
Estado de explotación de L. synagris segúnCope y Punt, (2009)
El árbol de decisiones de Cope y Punt infirió que en la mayoría de los años y sitios evaluados la especie tenía una biomasa desovante por debajo de los puntos de referencia, lo que indica la condición de sobrepesca observada a través de los análisis descritos. La condición de sobrepesca es teóricamente imposible si todos los peces hubieran podido reproducirse antes de la captura (Myers y Mertz, 1998). Froese sugiere un objetivo entre 90 y 100 % donde los peces que alcancen la madurez, permitiendo que cada pez se reproduzca al menos una vez antes de ser capturado (Froese, 2004). En este estudio, en ninguno de los sitios evaluados se alcanzó el umbral objetivo de Froese de 90 %. Esto sugiere que la mayoría de los peces capturados no han podido reemplazarse a sí mismos en la población y éste es un resultado bastante alarmante.
Los ecosistemas marinos son complejos y es probable que los cambios en los componentes individuales de los sistemas tengan consecuencias en otros lugares de los mismos sistemas (Scheffer et al., 2005). La sobrepesca no solo afecta a las poblaciones de peces, las redes tróficas y ecosistemas, sino también afecta a las personas que dependen de la pesca ya sea por su sustento económico, provisión e identidad cultural (Cinner et al., 2012; McClanahan et al., 2015; Purcell et al., 2016).
Todo lo anterior indica que la condición de efecto de desborde que se debe propiciar por la función de protección de las especies al interior de las áreas protegidas podría estar comprometida, ya que al ser removidos los ejemplares juveniles y los más grandes de la población no le estarían aportando la suficiente biomasa a los sectores externos de las áreas (Froese et al., 2004). Esta situación se agrava para L. synagris, dado que en la mayoría de los desembarques pesqueros del Caribe de Colombia su pesca se centra también en un alto porcentaje de peces inmaduros (Narváez et al., 2013; Duarte et al., 2018), con consecuencias que la han llevado a una disminución de su captura por unidad de esfuerzo (Duarte et al., 2018) y a la pérdida de la variabilidad genética (Landínez et al., 2009; Narváez et al., 2015).
Finalmente, es conveniente anotar que al interior de las áreas protegidas se deben fortalecer las medidas de manejo frente al uso y aprovechamiento de L. synagris, principalmente cuando se conoce que en estas áreas no sólo se protegen hábitats importantes para su alimentación y reproducción, sino que son un seguro contra posibles errores en el manejo pesquero por fuera de ellas (Austermühle, 2004). El número de pescadores que frecuentan estas áreas (~1150 entre las tres áreas; Martínez-Viloria et al., 2014) y los asociados al sector marino adyacente dificultan las acciones de control y vigilancia del personal técnico de PNNC y de la autoridad pesquera (AUNAP). Esto es un aspecto mundial de las pesquerías artesanales que con la creciente sobreexplotación pesquera (Muallil et al., 2014; Gough et al., 2020), se han convertido en un gran reto interinstitucional en los países en desarrollo, como Colombia, para poder administrarlas y minimizar los impactos pesqueros que generan sobre los recursos. Por lo tanto, el manejo pesquero de los recursos al interior y por fuera de las áreas protegidas debe darse en el marco de acciones conjuntas en las que se involucren las autoridades autónomas regionales, las de carácter policivo, marítimo y ambiental, los entes territoriales, la autoridad pesquera y Parques Nacionales Naturales. En esas acciones deberán considerarse aspectos técnicos-normativos, económicos y sociales que propendan por beneficiar a los pescadores ante la prohibición o regulación de la pesca comercial artesanal y del uso de artes que lesionan al recurso pargo rayado dentro y fuera de las áreas protegidas.
Con lo anterior en mente y teniendo en cuenta el efecto de desborde de las áreas protegidas como estrategia de conservación (Roberts et al., 2001; Kaunda-Arara y Rose, 2004; Di Lorenzo et al., 2016) se contribuye al aprovechamiento de los recursos pesqueros por fuera de las áreas, mediante diferentes actividades, siempre y cuando se realicen de manera responsable (FAO, 1995). Por tanto, se plantea que se fortalezcan acciones de acuerdo con los usos permitidos al interior de cada área protegida como: i) Prevención, control y vigilancia con apoyo de las autoridades que tienen dentro de sus funciones la protección del medio ambiente, a través de acciones conjuntas entre PNNC, Armada Nacional, AUNAP, para evitar las capturas de pargo rayado inmaduros y megadesovadores, ii) promover planes de monitoreo basados en la metodología descrita en este estudio que incorpore los resultados obtenidos como línea base, que permita validar acciones de manejo a través del seguimiento a las capturas de L. synagris, iii) incentivar la investigación que genere información pertinente para la toma de decisiones dentro y fuera de las áreas protegidas, iv) desarrollar estrategias de comunicaciones y educación ambiental que contribuyan a mantener informado al público sobre la condición de la especie y por qué no se debe consumir ejemplares juveniles, ni los más grandes que están presente en las áreas protegidas, v) generar acuerdos de conservación para la pesca de subsistencia (acorde con la reglamentación de PNNC) y vi) fortalecer el relacionamiento con otras autoridades en la región en busca de alternativas que contribuyan a la disminución de la presión sobre el recurso.
CONCLUSIONES
Al evaluar el estado de Lutjanus synagris en las áreas protegidas PNNT, PNNCRSB y VIPIS se evidenció que la especie se encuentra en una situación de sobrepesca por crecimiento y reclutamiento.
Los indicadores simples basados en las longitudes de captura configuraron un patrón de selectividad representado en la captura mayoritaria de ejemplares inmaduros (< Lmat) y en menor grado de individuos ubicados en el rango óptimo de longitud, lo que condujo a un nivel de biomasa desovante inferior al punto de referencia (25 % de SB), de acuerdo con lo establecido en el árbol de decisión propuesto por Cope y Punt (2009).
Los artes de pesca están generando un efecto negativo sobre las poblaciones de L. synagris, al capturar mayoritariamente individuos a la longitud media de captura-LMC por debajo < Lmat, los cuales son considerados inmaduros, aspecto que es más marcado en PNNCRSB y VIPIS. Además, existe una tendencia sobre los porcentajes de longitudes de captura que muestra señales que no benefician la condición de la especie al ser removido también individuos megadesovadores.
Los resultados obtenidos en este estudio evidencian que los indicadores basados en longitudes y el árbol de decisiones de Cope y Punt son una opción para que PNNC replique el modelo en otras áreas protegidas y generar respuestas de manejo en situaciones de datos limitados.
AGRADECIMIENTOS
Este trabajo hace parte de la tesis de HMV en la Maestría en Acuicultura y Ecología Acuática Tropical de la Universidad del Magdalena. Los autores agradecen a PNNC por el apoyo al facilitar el uso de la información proveniente del seguimiento a la presión por pesca realizado en las áreas protegidas entre 2006 - 2012
REFERENCIAS
Allen, G.R. 1985. Snappers of the world. An annotated and illustrated catalogue of lutjanid species known to date. FAO Spec. Cat. Fish. Synop.,125 (6): 208 p. [ Links ]
Altamar, J., J. Wong-Lubo, J. de la Hoz-M. e I. Martínez-Dallos. 2020. Evaluación de la selectividad de redes de enmalle y líneas de mano para la captura de cojinoa (Caranx crysos) en áreas de influencia marina del Parque Nacional Natural Tayrona. Bol. Invest. Mar. Cost., 49 (Supl. Esp.): 209-222. [ Links ]
Amorim, P., P. Sousa, M. Westmeyer and G.M. Menezes. 2018. Generic Knowledge Indicator (GKI): A tool to evaluate the state of knowledge of fisheries applied to snapper and grouper. Mar. Pol., 89: 40-49 [ Links ]
Arteaga, E.A., M.I. Críales-Hernández, D.Y. Rodríguez, P. Gómez-Canchong y L. Manjarrés. 2004. Dinámica reproductiva de los stocks de pargo rayado Lutjanus synagris (Linnaeus, 1758) en el área norte del Caribe colombiano. 297-315. En: Manjarrés, L. (Ed.). Pesquerías demersales del área norte del mar Caribe de Colombia y parámetros biológico-pesqueros y poblacionales del recurso pargo. Univ. Magdalena, Santa Marta. 317 p. [ Links ]
Aunap-Unimagdalena. 2014. Caracterización de los principales artes de pesca de Colombia y reporte del consolidado del tipo y número de artes, embarcaciones y UEP´s empleadas por los pescadores vinculados a la actividad pesquera. Univ. Magdalena, Santa Marta. 72 p. [ Links ]
Austermühle, S. 2004. Áreas marinas protegidas el futuro de la costa peruana. Mundo Azul, Conservación Internacional, Lima. 44 p. [ Links ]
Barreto, C. y C. Borda. 2009. Evaluación de recursos pesqueros colombianos. Inst. Col. Agropec. Subger. Pesca Acuic., Bogotá. 131 p. [ Links ]
Bent-Hooker, H.C. 2006. Biología, ecología y pesquería de la barracuda Sphyraena barracuda (Walbaum, 1792) (Pisces: Perciformes: Sphyraenidae) en la isla de San Andrés y los cayos Bolívar y Albuquerque, Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 63 p. [ Links ]
Cervigón, F., R. Cipriani, W. Fischer, L. Garibaldi, M. Hendrickx, A.J. Lemus, R. Márquez, J.M. Poutiers, G. Robaina y B. Rodríguez. 1992. Guía de campo de las especies marinas y de aguas salobres de la costa septentrional de Sur América. FAO, Roma, 513 p. [ Links ]
Chong, L., T. K. Mildenberger, M.B. Rudd, M.H. Taylor, J.M. Cope, T.A. Branch, M. Wolff and M. Stäbler. 2019. Performance evaluation of data-limited, length-based stock assessment methods. ICES J. Mar. Sci.,77(1): 97-108. [ Links ]
Cinner, J.E., T.R. McClanahan, N.A.J. Graham, T.M. Daw, J. Maina and S.M. Stead. 2012. Vulnerability of coastal communities to key impacts of climate change on coral reef fisheries. Glob. Environ. Chang., 22: 12-20p. [ Links ]
Claro, R. y K. Lindeman. 2008. Biología y manejo de los pargos (Lutjanidae) en el Atlántico occidental. Inst. Oceanol., CITMA, La Habana. 472 p. [ Links ]
Cochrane, K.L. (Ed.). 2005. Guía del administrador pesquero. Medidas de ordenación y su aplicación. FAO Doc. Tec. Pesca, 424: 231p. [ Links ]
Coleman, F., G.R. Koenig, J.A. Huntsman, M. Musick, J.C. Eklund, R.W. McGovern, G. Chapman, R. Sedberry and C.B. Grimes. 2000. Longlived reef fishes: the grouper-snapper complex. Fisheries, 25: 14-20. [ Links ]
Cope, J.M. and A.E. Punt. 2009. Length-based reference points for data-limited situations: applications and restrictions. Mar. Coast. Fish., 1(1): 169-186. [ Links ]
Di Lorenzo, M., J. Claudet and P. Guidetti. 2016. Spillover from marine protected areas to adjacent fisheries has an ecological and a fishery component. J. Nat. Cons., 32: 62-66. [ Links ]
Doncel, O. y J. Páramo. 2010. Hábitos alimenticios del pargo rayado, Lutjanus synagris (Perciformes: Lutjanidae), en la zona norte del Caribe colombiano. Lat. Am. J. Aquat. Res., 38(3): 413-426. [ Links ]
Dowling, N.A., A.D.M. Smith, D.C. Smith, A.M. Parma, C. M. Dichmont, K. Sainsbury and J.R. Wilson. 2019. Generic solutions for data-limited fishery assessments are not so simple. Fish Fish., 20: 174-188. [ Links ]
Duarte, L.O. and C.B. García. 1999. Diet of the lane snapper, Lutjanus synagris (Lutjanidae), in the Gulf of Salamanca, Colombia. Caribb. J. Sci., 35 (1-2): 54-63. [ Links ]
Duarte L.O., L. Manjarrés-Martínez, J De la Hoz-M., F. Cuello y J. Altamar. 2018. Estado de los principales recursos pesqueros de Colombia. Análisis de indicadores basados en tasas de captura, tallas de captura y madurez. AUNAP, Univ. Magdalena. Santa Marta. 37 p. [ Links ]
FAO. 1995. Code of conduct for responsible fisheries. Food and Agriculture Organization of the United Nations, Rome. 41 p. [ Links ]
FAO. 2018. El estado mundial de la pesca y la acuicultura. Cumplir los objetivos de desarrollo sostenible. Licencia: CC BY-NC-SA 3.0 IGO. Food and Agriculture Organization of the United Nations, Roma. 233 p. [ Links ]
Fontoura, N.F., A.S. Braun and P.C.C. Milani. 2009. Estimating size at first maturity (L50) from gonadosomatic index (GSI) data. Neotrop. Ichthyol., 7(2): 217-222. [ Links ]
Froese, R. 2004. Keep it simple: three indicators to deal with overfishing. Fish Fish, 5: 86-91. [ Links ]
Froese, R. and C. Binholan. 2000. Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. J. Fish Biol., 56: 758-773. [ Links ]
Froese, R., A. Stern-Pirlot, H. Winker and D. Gascuel. 2008. Size matters: how single-species management can contribute to ecosystem-based fisheries management. Fish. Res., 92: 231-241. [ Links ]
Froese, R., H. Winker, D. Gascuel, R. Sumaila and D. Pauly, 2016. Minimizing the impact of fishing. Fish. Res., 17: 785-802. [ Links ]
García, C.B., L.O. Duarte, J. Altamar and L. Manjarrés. 2007. Demersal fish density in the upwelling ecosystem off Colombia, Caribbean Sea: historic outlook. Fish. Res., 85: 68-73. [ Links ]
Gómez, G., R. Guzmán y R. Chacón. 2001. Parámetros reproductivos de Lutjanus synagris en el golfo de Paria, Venezuela. Zootec. Trop., 19 (3): 335-357. [ Links ]
Gough, C.L.A., K.M. Dewar, B.J. Godley, E. Zafindranosy and A.C. Broderick. 2020. Evidence of overfishing in small-scale fisheries in Madagascar. Front. Mar. Sci., 7: 317. [ Links ]
Guzmán, Y. K. 2010. Contribución al conocimiento biológico-pesquero de las principales especies de peces capturadas en el Parque Nacional Natural Corales del Rosario y de San Bernardo, sector Archipiélago Nuestra Señora del Rosario, Caribe colombiano. Trabajo Grado Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá. 152 p. [ Links ]
Hsieh, C., C.S. Reiss, J.R. Hunter, J.R. Beddington, R.M. May and G. Sugihara. 2006. Fishing elevates variability in the abundance of exploited species. Nature, 443: 859-862. [ Links ]
Jaimes, L. 2011. Aspectos biológicos-pesqueros de las principales especies ícticas capturadas en el sector de San Bernardo, Parque Nacional Natural Corales del Rosario y de San Bernardo, Caribe colombiano. Trabajo Grado Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá. 179 p. [ Links ]
Landínez-García, R.M., S.P. Ospina-Guerrero, D.J. Rodríguez-Castro, R. Arango and E. Márquez. 2009. Genetic analysis of Lutjanus synagris populations in the Colombian Caribbean. Cienc. Mar., 35(4): 321-331. [ Links ]
Longhurst, A. 2002. Murphy’s law revisited: longevity as a factor in recruitment to fish populations. Fish. Res., 56: 125-131. [ Links ]
López, A. 2009. Estado de explotación de peces con estrategias de vida contrastantes en el mar Caribe de Colombia. Análisis espacial y temporal de indicadores biológicos simples. Trabajo Grado Biol., Univ. Magdalena, Santa Marta. 69 p. [ Links ]
Luckhurst, B., J.M. Dean and M. Reichert. 2000. Age growth and reproduction of the lane snapperLutjanus synagris(Pisces: Lutjanidae) at Bermuda. Mar. Ecol. Prog. Ser., 203: 255 - 261. [ Links ]
Manjarrés, L. 1993. Eficiencia y selectividad de redes de enmalle en el área de Santa Marta. Proyecto integral de investigaciones y desarrollo de la pesca artesanal en el área de Santa Marta, INPA-CIID-Univ. Magdalena. Inf. Técn. Final, Santa Marta, 23 p. [ Links ]
Manjarrés, L. 2004. Pesquerías demersales del área norte del mar Caribe de Colombia y parámetros biológico-pesqueros y poblacionales del recurso pargo. Univ. Magdalena, Santa Marta. 318 p. [ Links ]
Manjarrés, L. (Ed). 2007. Caracterización participativa de las pesquerías de pequeña escala de las áreas protegidas de la Territorial Caribe. Inf. Técn. Final, Univ. Magdalena -PNNC-DTCA, Santa Marta. 86 p. [ Links ]
Manjarrés, L., J.C. Arévalo, D.J. Rodríguez y P. Gómez-Canchong. 2004. Dinámica poblacional y manejo del stock de pargo rayado (Lutjanus synagris) (Linneaus, 1758) de La Guajira (Caribe colombiano). 297-315. En: Manjarrés, L (Ed). Pesquerías demersales del área norte del mar Caribe de Colombia y parámetros biológico-pesqueros y poblacionales del recurso pargo. Univ. Magdalena, Santa Marta. 315 p. [ Links ]
Martínez-Viloria, H. 2009. Análisis regional del estado de los recursos hidrobiológicos al interior de las áreas protegidas a cargo de la Dirección Territorial Caribe-Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales. Inf. Técn., PNNC-DTCA, Santa Marta. 51p. [ Links ]
Martínez-Viloria, H. 2014. Análisis sobre la presión por pesca en áreas protegidas con jurisdicción marino-costeras adscritas a las Dirección Territorial Caribe de Parques Nacionales Naturales de Colombia. Inf. Técn., PNN-DTCA, Santa Marta. 64 p. [ Links ]
Martínez-Viloria, H., J.C. Narváez, R. Rivera y O.D. Solano. 2006. Evaluación de la selectividad del trasmallo en la pesquería artesanal de la zona deltaico estuarina del río Sinú, Caribe colombiano. Intrópica, 3: 33-41. [ Links ]
Martínez-Viloria, H., L. Martínez, A. Vargas-Pineda y J.C. Narváez. 2011. Efectos de la pesca sobre los recursos hidrobiológicos del Parque Nacional Natural Corales del Rosario y de San Bernardo. 273-289. En: Zarza-González, E. (Ed). El entorno ambiental del Parque Nacional Natural Corales del Rosario y de San Bernardo. PNNC, Bogotá. 416 p. [ Links ]
Martínez-Viloria. H, R. Franke, P. Saldaña, M. Cano-Correa, L.E. Angarita, C. García, L. Martínez, A. Castro, S. Posada, C. Gómez, E. Brugés, J.C. Narváez, E. Viloria y C. López. 2014. Caracterización del uso y aprovechamiento de recursos hidrobiológicos en áreas protegidas de Parques Nacionales Naturales del Caribe de Colombia. Bol. Invest. Mar. Cost., 43(2): 277-306. [ Links ]
McClanahan, T., E.H. Allison and J.E. Cinner. 2015. Managing fisheries for human and food security. Fish Fish., 16: 78-103. [ Links ]
Mejía, L. E. y A. Acero (Eds.). 2002. Libro rojo de peces marinos de Colombia. Serie Libros rojos de especies amenazadas de Colombia. Inst. Cienc. Nat., Univ. Nal. Colombia, Invemar y Min. Medio Ambiente, Bogotá. 174 p. [ Links ]
Muallil, R.N., S.S. Mamauag, R.B. Cabral, E.O. Celeste-Dizon and P.M. Aliño. 2014. Status, trends and challenges in the sustainability of small-scale fisheries in the Philippines: Insights from FISHDA (Fishing Industries’ Support in Handling Decisions Application) model. Mar. Pol., 44: 212-221. [ Links ]
Myers, R.A. and G. Mertz. 1998. The limits of utilization: a precautionary approach. Ecol. Appl., 8: 165-169 [ Links ]
Narváez, J.C., M. Rueda, E. Viloria, J. Blanco, J.A. Romero y F. Newmark. 2005. Manual del Sistema de Información Pesquera del Invemar (Sipein V.3.0): Una herramienta para el diseño de sistemas de manejo pesquero. Serie de documentos generales del Invemar, 18. Santa Marta. 128 p. [ Links ]
Narváez, J.C., F. Herrera y J. Blanco. 2008. Efecto de los artes de pesca sobre el tamaño de los peces en una pesquería artesanal del Caribe colombiano. Bol. Invest. Mar. Cost., 37 (2): 163-168. [ Links ]
Narváez , B.J.C., J. de La Hoz, J. Blanco, F. Bolívar, R. Rivera, T. Álvarez, A. Mora, O. Bran y C. Riascos. 2013. Tallas mínimas de captura para el aprovechamiento sostenible de las principales especies de peces comerciales de Colombia. Univ. Magdalena, Santa Marta. 36 p. [ Links ]
Narváez, J.C., G. Orozco B., J.C. Aguirre P., E. Muñoz, J. Quintero, F. Bolívar M., T. Narváez B., R. Mendoza U., L. Castro G. y L.O. Duarte. 2015. Estado genético y conectividad de la población del pargo rayado Lutjanus synagris en sitios dentro y fuera de las áreas marinas protegidas (AMP) de Parques Nacionales Naturales de la Territorial Caribe. Inf. Técn. Final. Patrimonio Natural y Univ. Magdalena, Santa Marta. 25 p. [ Links ]
Pardo, R. 2011. Efecto de las artes de pesca sobre la estructura de tallas de pargo rayado (Lutjanus synagris) en el golfo de Salamanca, mar Caribe de Colombia. Tesis Ing. Pesq., Univ. Magdalena, Santa Marta. 98 p. [ Links ]
PNNC. 2006. Plan de manejo del Parque Nacional Natural Tayrona, Santa Marta. 298 p. [ Links ]
PNNC. 2017. Plan de manejo Vía Parque Isla de Salamanca 2017-2022, Santa Marta. 227 p. [ Links ]
PNNC. 2020. Plan de manejo Parque Nacional Natural Corales del Rosario y de San Bernardo, Cartagena. 228 p. [ Links ]
Pope, J.A., A.R. Margetts, J. Hamley y E.F. Akyüz. 1983. Manual de métodos para la evaluación de las poblaciones de peces. Parte 3. Selectividad del arte de pesca. FAO Doc. Téc. Pesca, 41(1): 1-56. [ Links ]
Purcell, S.W., P. Ngaluafe, S.J. Foale, N. Cocks, B.R. Cullis and W. Lalavanua. 2016. Multiple factors affect socioeconomics and wellbeing of Artisanal Sea cucumber fishers. PLoS One, 11:165633. [ Links ]
Roberts, C., J. Bohnsack, F. Gell, J. Hawkins and R. Goodridge. 2001. Effects of marine reserves on adjacent fisheries. Science, 294: 1920-1923. [ Links ]
Rodríguez, J., J. Arévalo y L. Manjarrés. 1999. Aspectos biológico-pesqueros de los pargos rayado (Lutjanus synagris) y cebal (L. analis). Bol. Cient. INPA, 6: 53-75. [ Links ]
Scheffer, M., S. Carpenter and B. de Young. 2005. Cascading effects of overfishing marine systems. Trends Ecol. Evol., 20: 579-581. [ Links ]
Sparre, P. y S. Venema. 1995. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1: Manual. FAO. Doc. Técn. Pesca, 306/1, 420 p. [ Links ]
Viaña, J., A. Medina, M. Barros, L. Manjarrés, J. Altamar y M. Solano. 2004. Evaluación de la ictiofauna demersal extraída por la pesquería industrial de arrastre en el área norte del Caribe Colombiano (enero 2000-junio 2001). 115-151p. En: Manjarrés, L. (Ed.). Evaluación de las pesquerías demersales del área norte del Caribe colombiano y parámetros biológico-pesqueros y poblacionales del recurso pargo. Univ. Magdalena, Santa Marta. 151 p. [ Links ]
Recibido: 09 de Agosto de 2020; Aprobado: 19 de Abril de 2022
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
