Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.11 n.1 Manizales ene./dic. 2007
Liliana Arango-B.1
José Mauricio Montes-R.2
Diego Alejandro López-P.3
José Orlando López-P.4
1 Ingeniera Agrónoma, Universidad de Caldas, Manizales. E-mail: liliana.arango@gmail.com
2 Ingeniero Agrónomo, Universidad de Caldas, Manizales. E-mail: jmpamplonman@gmail.com
3 Estudiante de Ingeniería Agronómica, Universidad de Caldas, Manizales. E-mail: diegolopez@colombia.com
4 Estudiante de Derecho, Universidad de Caldas, Manizales. E-mail: hiroyui.jose@latinmail.com
Resumen
Las mariposas diurnas, escarabajos coprófagos y hormigas son útiles para obtener información rápida y eficiente sobre la diversidad de un ecosistema. Con este objetivo se realizó su caracterización en pastizales y relictos boscosos del Ecoparque Alcázares–Arenillo (EAA), ubicado en Manizales, departamento de Caldas. Las mariposas se colectaron con trampas Van Someren Rydon y jama; los escarabajos coprófagos, con trampas de caída cebadas con excremento, y las hormigas, mediante captura manual, trampas Winkler, cebos epigeos y trampas de caída instaladas en transectos lineales. Los resultados indicaron que el 43% de las especies de mariposas colectadas son exclusivas de los fragmentos boscosos y que la similitud en la composición entre los componentes del paisaje del EAA es baja. Los escarabajos coprófagos presentaron los valores más altos de abundancia y riqueza de especies en el fragmento más grande y los menores en potrero. Las hormigas presentaron diversidad similar y baja similitud entre los transectos, y se destacó que 19 de las 57 especies colectadas obtuvieron tan solo un registro en todo el muestreo. Estos resultados permitieron concluir que los relictos boscosos del EAA representan un refugio para algunas especies de estos grupos de insectos, que cada elemento del paisaje contribuye a la diversidad de la zona y que deben tomarse medidas oportunas que permitan la conservación, en especial de las especies asociadas con los fragmentos boscosos y en peligro de extinción local.
Palabras clave
Ecoparque Alcázares, Biodiversidad, Hormigas, Mariposas, Escarabajos coprogfagos.
Abstract
Diurnal butterflies, dung beetles and ants are useful in obtaining fast and efficient data on the diversity of an ecosystem. The latter was the objective of the characterization carried out on these groups of insects in forest fragments and pastures in the Alcázares–Arenillo Ecopark (EAA) located in Manizales in the Caldas department. The butterflies were collected with Van Someren Rydon traps and an entomological net; the dung beetles with pitfall traps baited with excrement; and the ants by means of manual capture, epigeous baits, pitfall traps and Winkler traps installed in linear transects. The results showed that 43% of the collected butterflies species are exclusive of the forest fragments and that the similarity in the composition between the components of the EAA landscape is low. The dung beetles obtained the highest values of species abundance and richness in the biggest fragment, and the lowest in pasture. The ants presented similar diversity and low similarity among the transects, showing that 19 out of the 57 collected species obtained just one entry in all the sampling. These results led to the conclusion that EAA is a refuge for some species of these biological groups, each element of the landscape contributes to the diversity of the zone. Therefore, immediate actions must be taken to conserve, especially species associated with forest fragments and that are susceptible of disappearing in the future.
Key words
Alcazares Ecopark, Biodiversity, Ants, Butterflies, Dung Beetles.
INTRODUCCIÓNColombia, con aproximadamente el 10% de las especies descritas en el mundo (VILLAREAL et al., 2004), ve amenazada su biodiversidad por factores como la deforestación y el aprovechamiento insostenible de los recursos naturales, factores que ocasionan la pérdida de gran parte del área boscosa y la extinción masiva de especies. Este fenómeno es particularmente grave en los trópicos, donde se concentra la mayor biodiversidad del planeta (ANDRADE, 1993).
A pesar de la disminución de los paisajes naturales, aún se conservan refugios de flora y fauna que han tolerado la intervención del hombre. En el municipio de Manizales uno de estos lugares lo constituye el Ecoparque Alcázares-Arenillo, el cual posee fragmentos de bosque que deben conservarse por su importancia ambiental.
Para la conservación de ecosistemas es necesario evaluar los componentes de la biota. Sin embargo, esto requiere de grandes inversiones de tiempo y dinero, y hace necesario enfocar estos estudios en grupos que generen información rápidamente y con demandas económicas mínimas; a dichos grupos se les ha denominado bioindicadores. Según FAGUA (1996), el uso de especies indicadoras se constituye en una herramienta de primera mano para el análisis ambiental.
Dentro de los insectos, las mariposas diurnas (FAGUA, 1996, 1999), los escarabajos coprófagos (ESCOBAR, 2004) y las hormigas (ALONSO & AGOSTI, 2000; ARMBRECHT & CHACON, 1997; OLIVER & BEATTIE, 1996) han sido reconocidos como bioindicadores útiles en trabajos de monitoreo y evaluación ambiental, principalmente por la sencillez y rapidez de su muestreo y por su taxonomía relativamente bien conocida con respecto a otros grupos de insectos.
Con base en lo anterior se desarrolló el presente trabajo, con el objetivo de caracterizar las poblaciones de mariposas diurnas, escarabajos coprófagos y hormigas presentes en los diferentes elementos del paisaje del Ecoparque Alcázares–Arenillo.
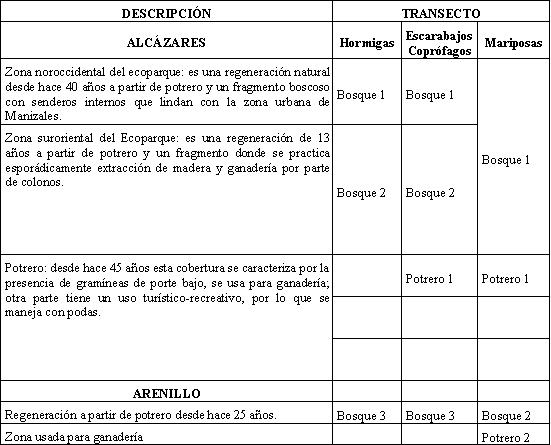
MATERIALES Y MÉTODOS Zona de estudio
El Ecoparque Alcázares-Arenillo (EAA) tiene una extensión total de 70,55 ha, se ubica en el municipio de Manizales, departamento de Caldas, en la vertiente occidental de la Cordillera Central a los 5º 04 Norte y 75º 32 Oeste en un rango altitudinal entre 1.730 y 1.960 msnm y con precipitación de 2.223 mm, características que lo ubican según HOLDRIDGE (1982) en la zona de vida correspondiente a bosque húmedo premontano. El ecoparque limita al norte con el barrio Alcázares y al sur con la vereda el Arenillo.
En el EAA se reconocen principalmente dos elementos del paisaje: uno está compuesto de zonas boscosas con períodos de regeneración entre 10 y 40 años, donde las especies vegetales son básicamente especies pioneras; el otro elemento está compuesto por pasturas destinadas a la producción de ganado y por zonas verdes o jardines con plantas ornamentales.
Los muestreos se realizaron en los meses de febrero y marzo del año 2005. La metodología utilizada para cada grupo se describe a continuación:
Mariposas
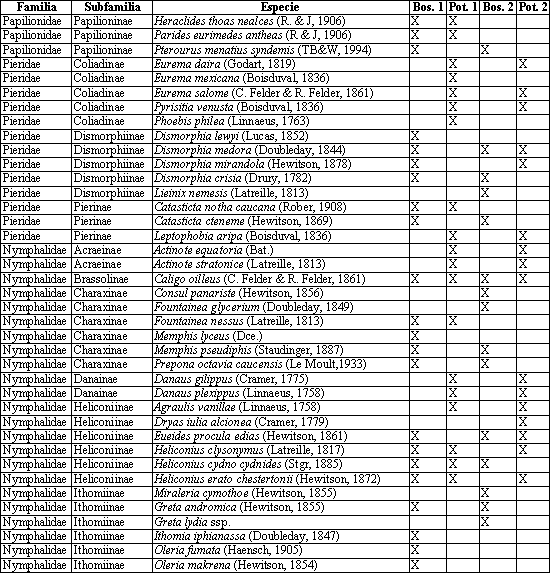
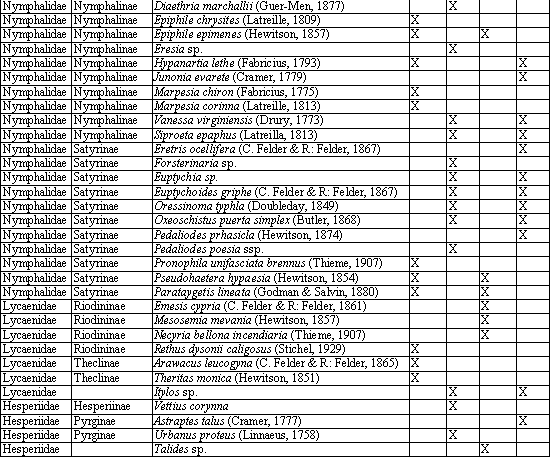
Para capturar las mariposas se identificaron, inicialmente, las zonas boscosas y las áreas abiertas de Alcázares y el Arenillo (Tabla 1). Además, se colectaron las mariposas con red entomológica y se dedicó un esfuerzo de muestreo de 36 horas a cada zona para un total de 144 horas. Las mariposas se depositaron en sobres de papel milano rotulado con datos de colecta. La identificación se realizó con base en las ilustraciones de DEVRIES (1987), DABRERA (1981, 1984, 1987, 1988, 1994), PARRA et al. (2000) y VALENCIA et al. (2005), y se actualizaron los nombres científicos con base en el atlas de los Lepidoptera neotropicales (LAMAS, 2004) y con la asesoría de los especialistas Julián Salazar (Museo de Historia Natural de la Universidad de Caldas) y Efraín Henao (Profesor de entomología de la Universidad de Caldas). Con los datos registrados se realizó un análisis descriptivo basado en la determinación porcentual de cada subfamilia, el cálculo del número de especies únicas, exclusivas, totales y comunes, y el índice de similitud de Jaccard (LUDWING & REYNOLDS, 1988).
Escarabajos coprófagos
Se establecieron 4 transectos (Tabla 1), constituidos por 10 trampas de caída Pitfall cebadas con excremento humano (VILLARREAL et al., 2004), separadas por 20 m una de la otra. Las trampas se recolectaron a las 72 horas después de su instalación.
Los individuos se identificaron hasta el nivel de género con la clave de MEDINA & LOPERA (2000) y hasta el nivel taxonómico de especie con la ayuda de claves específicas como las de HOWDEN & YOUNG (1981) y VITOLO (2000). Para evaluar la diversidad, se calculó el índice de Shannon evaluado por la prueba t propuesta por HUTCHESON (1970), citado por ZAR (1996), y el índice de similitud de Jaccard (LUDWING & REYNOLDS, 1988).
Finalmente, se clasificaron los escarabajos coprófagos en pequeños (≤ 10 mm) y grandes (≥ 10 mm), según lo propuesto por ESCOBAR (2004) y según el tipo de nidificación en rodadores y cavadores. Para el análisis de estos grupos, al igual que para el número de individuos totales por transecto, se realizó un análisis de varianza por rangos y una prueba de Tukey modificada para información no paramétrica (ZAR, 1996).
Hormigas
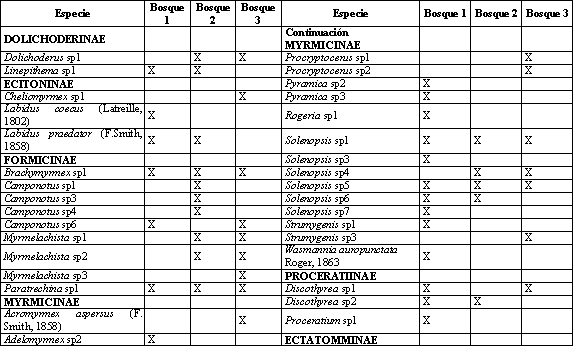
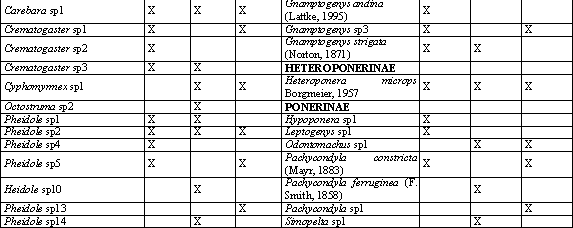
Se establecieron 3 transectos (Tabla 1). Cada uno de estos estuvo compuesto por 10 estaciones de muestreo separadas por 10 m una de la otra, en las cuales se aplicaron los siguientes métodos de captura: trampas Winkler, trampas de caída y captura manual; se aplicó, además, un esfuerzo de muestreo de 10 minutos en cada punto y cebos de atún al nivel del suelo, que se colectaron después de 2 horas (VILLAREAL et al., 2004).
El material colectado se identificó hasta el nivel de género utilizando las claves para la región neotropical de PALACIO & FERNÁNDEZ (2003), y hasta de especie con la ayuda de claves específicas (PALACIO, 1999). Se calculó el índice de diversidad de Shannon evaluado por una prueba t propuesta por HUTCHESON (1970), citado por ZAR (1996), y el índice de similitud de Jaccard (LUDWING & REYNOLDS, 1988). Para el análisis se tomó como abundancia el número de registros en cada estación de muestreo en lugar del número de individuos.
Las mariposas, los escarabajos coprófagos y las hormigas colectadas se procesaron según VILLAREAL et al. (2004) y se depositaron en el laboratorio de entomología de la Universidad de Caldas.
RESULTADOS Y DISCUSIÓN
Mariposas
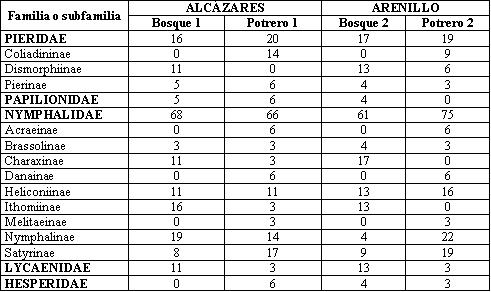
Se capturó un total de 79 especies pertenecientes a cinco familias, 17 subfamilias y 60 géneros. En Alcázares se encontraron 35 y 37 especies en Potrero 1 y en Bosque 1, respectivamente. En el Arenillo se colectaron 52, 32 y 23 especies en Bosque 1, en Potrero 2 y en Bosque 2, respectivamente (Anexo 1). Puede observarse que para Alcázares el número de especies entre las zonas abiertas y el bosque es similar, lo cual se debe a que en Alcázares los fragmentos de bosque son pequeños, dispersos y ubicados dentro de una matriz de potrero, coincidiendo con DEVIRIES (1987) en que la riqueza de mariposas en bosques y bordes de bosque es ligeramente diferente entre ambos hábitats.
En la tabla 2 se presenta la distribución porcentual de familias y subfamilias en la comunidad de mariposas, puede observarse que aunque las familias Pieridae y Nymphalidae tienen un porcentaje similar dentro de los transectos evaluados, sus subfamilias presentan una tendencia diferente. En el caso de Pieridae, la subfamilia Coliadinae presenta mayor porcentaje en zonas abiertas; en el caso de Dismorphinae, en fragmentos de bosque, mientras que Pierinae es similar en todos los transectos. En el caso de Nymphalidae, las subfamilias Ithomiinae y Charaxinae poseen una mayor proporción en fragmentos de bosque; en Ithomiinae esto se debe a que la mayoría de especies son de hábitos umbrófilos y no requieren mucha energía calórica para volar (GARCÍA et al., 2002), mientras que en Charaxinae se debe a que las plantas nutricias son principalmente árboles (DEVRIES, 1987). Por el contrario, las subfamilias Satyrinae, Acraeinae, Danainae, Melitaeinae son más abundantes en zonas abiertas; esta relación se debe a que la mayoría de las especies encontradas de estas subfamilias presenta hábitos heliófilos y a que se alimentan de plantas típicas de zonas abiertas. Al analizar los porcentajes que ocupan las subfamilias Heliconiinae, Nymphalinae y Brassolinae, en los transectos no se encontró una relación con el lugar donde se encontraron.
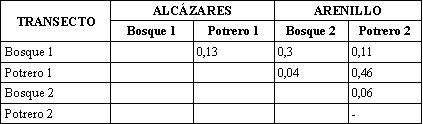
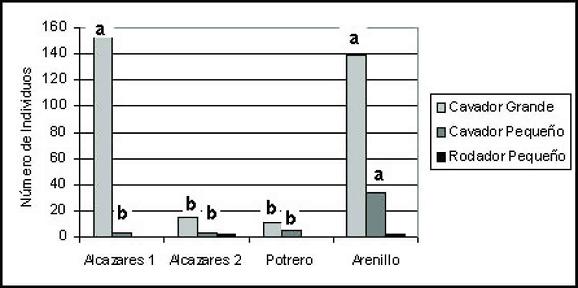
Al calcular el porcentaje de especies comunes entre dos sitios mediante el coeficiente de Jaccard, se encontró que la mayor similitud se presentaba al comparar las zonas abiertas de Alcázares y Arenillo con un valor medio de 0,46, lo cual se explica por la cercanía de estos lugares, la poca diferencia altitudinal y la similitud de los ecosistemas en cuanto a la incidencia de radiación solar. La similitud de los transectos de zonas boscosas de Alcázares y Arenillo fue de 0,3, las áreas boscosas y los potreros presentan las mayores diferencias en la composición de especies, indicando que existe complementariedad entre los elementos del paisaje.
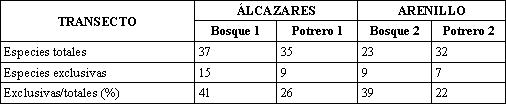
El mayor número de especies exclusivas se encontró en Bosque 1 con 15, seguido de Potrero 1, Bosque 2 y Potrero 2 con 9, 9 y 8 respectivamente. El porcentaje de especies exclusivas en relación con el total de cada transecto es de 41 y 39 para Bosque 1 y 2, respectivamente, y de 26 y 22% para las Potrero 1 y 2 (Tabla 4). Lo anterior indica que si los fragmentos de bosque desaparecieran, entonces se perdería el 41% de las mariposas de Alcázares y el 39% de las mariposas de Arenillo encontradas en esta investigación.
Coleópteros coprófagos
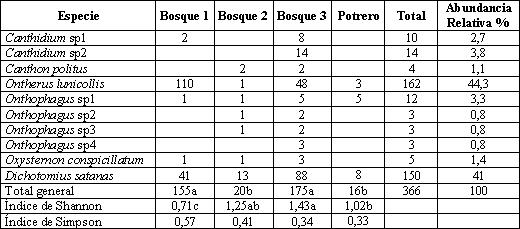
Se colectaron 366 individuos distribuidos en seis géneros y 10 especies (Tabla 5). Dichotomius satanas, Ontherus lunicollis y Onthophagus sp.1 se encontraron en todos los transectos, además, los dos primeros obtuvieron la mayor abundancia relativa con el 44,3% para O. lunicollis y 41% de los especímenes colectados para D. satanas. MEDINA et al., (2002) encontró un resultado similar en cuanto a la abundancia de O. lunicollis en el parque Ucumarí.
La menor abundancia se encontró en los transectos Potrero 1 y Bosque 2, en donde se colectaron 20 y 16 individuos respectivamente. Estos transectos no presentaron diferencias estadísticas entre sí, pero ambos fueron significativamente diferentes (P ≥ 0.05) en los transectos Bosque 1 con 155 y Bosque 3 con 175 individuos (Tabla 5). Las diferencias en abundancias pueden ser debidas a la cercanía con la población urbana y rural del EAA en Bosque 1 y 3, que garantiza a los escarabajos coprófagos disponibilidad de alimento representado en excremento humano y de animales domésticos, lo que favorece el aumento de las poblaciones de estos insectos. Esta afirmación está basada en lo mencionado por HALFFTER & MATTHEWS (1966): La presencia o ausencia de una abundante fuente de excremento es uno de los factores que pueden determinar la presencia y abundancia de Scarabaeinae.
Otro factor que gobierna la distribución de escarabajos coprófagos es el tipo de suelo (HALFFTER & MATTHEWS, 1966), por lo cual debe tenerse en cuenta que las zonas Potrero 1 y Bosque 2 (regeneración desde hace 13 años) han sido afectadas por procesos erosivos y de compactación producto del pisoteo de ganado, factores que pueden haber generado dificultades en el establecimiento de los escarabajos coprófagos, lo que explica su menor abundancia en estas zonas.
En el transecto Bosque 1 se registraron cinco especies, en Bosque 2 siete, en Potrero 1 tres y en el Bosque 3 se colectaron todas las especies reportadas en el estudio (10 en total). Onthophagus sp4 y Canthidium sp2 solo se registraron en este transecto.
En cuanto a la riqueza, se encontró en la zona de Potrero 1 el menor número de especies (3) de los transectos. ESCOBAR (1997) y MEDINA et al. (2002) también encontraron baja riqueza de escarabajos coprófagos en los potreros en comparación con los bosques. Esto puede deberse a algunas características ambientales de los potreros tales como la mayor temperatura y menor humedad en comparación con las zonas boscosas cercanas que desfavorecen el establecimiento de estos insectos (ESCOBAR, 1994), también a otras condiciones microclimáticas más variables (BUSTOS & LOPERA, 2003) y a la desecación del alimento que lo inhabilita para el uso de los escarabajos coprófagos (ESCOBAR, 1997).
Al comparar los índices de diversidad de Shannon (Tabla 5), se encontró que el Bosque 3 obtuvo el mayor valor con 1,43, siendo diferente significativamente (p ≤ 0,001) de los transectos Bosque 1 (0,71) y Potrero 1 (1,02) (P < 0.001), pero no del Bosque 2 (1,25) (P ≥ 0.5). Sin embargo, debe tenerse en cuenta que en Bosque 3 se presentó una mejor distribución de la abundancia, lo cual es confirmado por el menor valor del índice de Simpson (Tabla 5); ambos índices indican que hay mayor diversidad en este transecto.
Según AMAT et al. (1997), el área relictual más grande de un bosque altoandino presenta la mayor riqueza y diversidad, lo cual coincide con lo encontrado en el Bosque 3, ya que es el fragmento más grande con condiciones microclimáticas que favorecen el establecimiento de un mayor número de especies.
Letras diferentes representan diferencia estadística Pruebas Tukey (K= 3; α < 0.05)
El coeficiente de similitud de Jaccard (Tabla 6) presentó valores medios y altos en la mayoría de los datos, excepto entre Potrero 1 y Bosque 3 con un valor de (0,30), los cuales tienen la menor y mayor riqueza. Coincidiendo con ESCOBAR (1997, 2004) en que las zonas más conservadas y las de potrero presentan las mayores diferencias en la composición de la comunidad de Scarabaeinae.
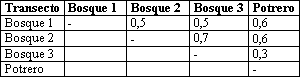
Al observar las diferencias en abundancia, riqueza y diversidad se puede deducir que gran parte de los escarabajos coprófagos del EAA prefieren habitar en el bosque, siguiendo la tendencia de la mayoría de especies en el Neotrópico (GILL, 1991), las cuales se ven afectadas por cambios en tamaño, distribución (ESCOBAR, 1994) y presencia de la cobertura arbórea (HALFFTER & ARELLANO, 2002) (HALFFTER & MATTHEWS, 1966) de zonas boscosas, cambios que imponen nuevas tendencias en la diversidad, estructura y composición de este grupo de insectos (ESCOBAR, 2004).
Al clasificar los coprófagos según el tipo de nidificación y tamaño se encontró que el 87% de los individuos colectados eran cavadores grandes (Fig. 1) de las especies O. lunicolis, D. satanas y Oxystenon conspicillatum, el número de individuos de este grupo para los transectos Bosque 1 y Bosque 3 fue de 152 y 139 mientras para Bosque 2 y Potrero 1 presentó valores de 15 y 11 respectivamente. La prueba Tukey (α ≤ 0.05) no presentó diferencias significativas entre Bosque 1 y Bosque 3, sin embargo se presentaron diferencias al comparar estos transectos con los dos últimos. La mayor abundancia puede estar favorecida por la cercanía a la población que representa una mayor cantidad de recursos alimenticios (excremento de animales domésticos) en Bosque 1 y Bosque 3 y las diferencias en el tipo de suelo explicadas anteriormente.
Los cavadores pequeños se agruparon en seis especies representando el 12% de los individuos; se colectaron tres en Bosque 1 y Bosque 2, cinco en Potrero 1 y en Bosque 3, 34 individuos presentando diferencias significativas con los demás transectos (α < 0.05). La mayor abundancia de escarabajos coprófagos de especies pequeñas se presenta generalmente en áreas boscosas (ESCOBAR, 2004) explicando de esta manera el mayor número de especies de este grupo en Bosque 3.
Los rodadores pequeños con sólo 1 % estuvieron representados por la especie Canthon politus, registrada por AMAT et al. (1997) y MEDINA et al. (2002) sólo en bosques; la baja abundancia de esta especie en el EAA podría estar indicando que es sensible a procesos de fragmentación.
Hormigas
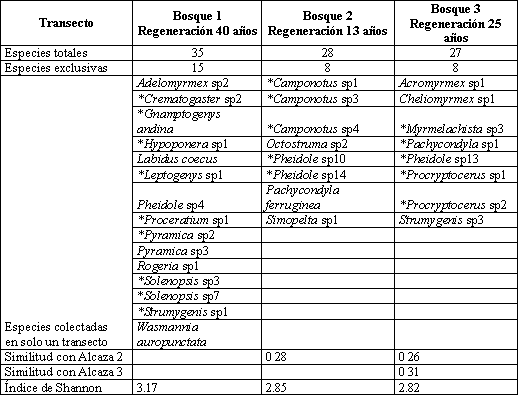
Se encontró un total de 57 especies pertenecientes a 30 géneros y ocho subfamilias, 35 se colectaron en Bosque 1, 28 en Bosque 2, y 27 en el Bosque 3 (Tabla 7), La subfamilia Myrmicinae presentó los mayores abundancia en los tres transectos, seguido por Ectatomminae en Bosque 1, Dolichoderinae en Bosque 2 y Formicinae en el Bosque 3.
Los géneros Pheidole y Solenopsis habitantes comunes de hojarasca (FERNANDEZ, 2003), presentaron la mayor abundancia en todos los transectos a excepción de Bosque 2 en donde Linepithema que es un género abundante y característico en áreas abiertas y perturbadas (KING, et al., 1998) ocupa la segunda posición lo cual ocurre posiblemente por las actividades de extracción de madera y ganadería practicadas allí por parte de colonos, adicionalmente este fragmento tiene menor tiempo en regeneración (13 años) que Bosque 1 y 3 (40 y 25 años respectivamente), resultados similares se reportaron por ESTRADA & FERNÁNDEZ (1999) quienes encontraron que la composición de la comunidad de hormigas varía en un gradiente sucesional.
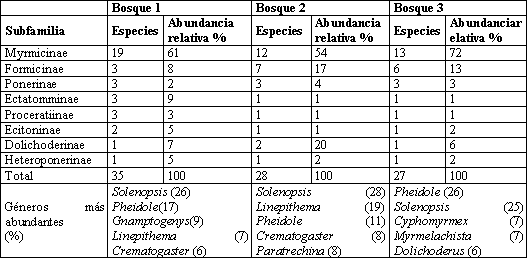
Los sacos Winkler registraron el mayor número de géneros y especies en comparación con los demás métodos de captura (Tabla 8), con 23 de los 30 géneros y 38 de las 57 especies colectadas confirmando lo planteado por DELABIE et al. (2000) en que los sacos Winkler son el método más eficiente para evaluar hormigas de hojarasca, con este método la mayor frecuencia de captura se presentó en los géneros Solenopsis, Pheidole, y Heteroponera éste último con sólo una especie Heteroponera microps. Con los cebos se capturaron hormigas que generalmente son abundantes y conspicuas y que también se colectaron con los otros métodos. La captura manual contribuyó a complementar el muestreo con géneros de hábitos arborícolas como Crematogaster, Myrmelachista, Procryptocerus y Camponotus.

Los índices de similitud presentaron valores bajos (Tabla 9). La fragmentación y el disturbio del ambiente unido a las condiciones topográficas crean microhábitats con diferencias en temperatura, disponibilidad de sitios para anidar, suministro de comida y estructura del microhábitat que afectan la distribución de las poblaciones de hormigas, según los requerimientos ambientales de cada especie ANDERSEN (2000). De está manera, la heterogeneidad del paisaje presente en el EAA parece reflejarse también en la composición de hormigas.
De las 57 especies, 31 se encontraron en sólo un transecto de los cuatro evaluados (exclusivas), y de estas, 19 poseen un único registro (únicas). Esto indica una baja abundancia de estas especies, lo que constituye un problema si se tiene en cuenta que el área en potreros que separa los fragmentos de bosque restringe el intercambio genético entre ellos, dificultando la conservación de estas poblaciones debido a que ciertas especies pueden disminuir o incluso extinguirse, si el área del fragmento de hábitat es muy pequeña o está muy alejada de otros hábitats fuente (ANDRADE, 1993). Resultados similares encontró ARMBRECHT et al. (2001) en fragmentos de bosque, por lo que concluyó que cada uno constituye un refugio para preservar la biodiversidad.
El índice de Shannon (Tabla 9) no presentó diferencias significativas, lo cual indica que cada transecto posee una diversidad similar y esto, sumado a la baja similitud de la composición de hormigas entre los transectos, indica que todos los fragmentos son importantes para la conservación de su diversidad en el EAA.
CONCLUSIONES
Fragmentos como los presentes en el EAA contribuyen a reducir el efecto local que produce la deforestación y la destrucción del ambiente en la biodiversidad, debido a que refugian especies de escarabajos coprófagos, hormigas, mariposas diurnas y probablemente a otros grupos biológicos asociados a bosques.
La abundancia, riqueza y distribución de las mariposas, escarabajos coprófagos y hormigas del EAA es diferente en cada elemento del paisaje y estas diferencias se deben principalmente al tipo de vegetación.
AGRADECIMIENTOSLos autores expresan su agradecimiento a Misael Salgado y Gustavo Zabala, al Centro Nacional de Investigaciones en Café Cenicafé, a la Universidad de Caldas, a la ONG Pangea y a Julián A. Salazar E. (MVZ), por sus correcciones en la taxonomía de las mariposas.
BIBLIOGRAFÍA ALONSO, L. E. & AGOSTI, D., 2000.- Biodiversity studies, Monitoring, and ants: An Overview: 1-8 (en) AGOSTI, et al., Ants: Standard Methods for measuring and monitoring biodiversity. Smithsonian Institution Press. Washington. 280 p. [ Links ]
AMAT, G.; LOPERA T., & AMEZQUITA,, S. J., 1997.- Patrones de distribución de escarabajos coprófagos (Coleoptera: Scarabaeidae) en relicto del bosque altoandino, cordillera oriental de Colombia. Caldasia, 19 (1-2): 191-204. [ Links ]
ANDRADE, G. I., 1993.- Biodiversidad y Conservación en Colombia: 25-42 (en) CÁRDENAS, S. & CORREA, H. D. (eds.). Nuestra Diversidad Biológica. Fundación Alejandro Ángel Escobar. Editorial Presencia. Bogotá, Colombia: 296 p. [ Links ]
ARMBRECHT, I. & CHACON, P., 1997.- Composición y diversidad de hormigas en bosques secos relictuales y sus alrededores, en el Valle del Cauca, Colombia. Revista Colombiana de Entomología, 23 (1-2): 45-50. [ Links ]
ARMBRECHT, I.; TISCHER, I. & CHACON, P., 2001.- Nested subset and partition in ant assemblages (Hymenoptera, Formicidae) of Colombian dry forest fragments. Pan-Pacific Entomologist, 77 (3): 196-209. [ Links ]
BUSTOS, L. F. & LOPERA, A., 2003.- Preferencia por cebo de los escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de un remanente de bosque seco tropical al norte del Tolima (Colombia): 59-65. (en) ONORE, G.; REYES, P. & ZUNINO, M. (comp). Escarabaeidos de Latinoamérica: Estado del conocimiento. Monografías Tercer Milenio. Sociedad Entomológica Aragonesa. Zaragoza. 86 p. [ Links ]
DABRERA, B., 1981.- Butterflies of the Neotropical Region. Parte I. Papilionidae y Pieridae Lansdowne Editions. 172 p. [ Links ]
__________., 1984.- Butterflies of the Neotropical Region. Parte II. Danaidae, Ithomiinae, Heliconiinae y Morphidae Hill House Editions. 382 p. [ Links ]
__________., 1987.- Butterflies of the Neotropical Region. Parte III. Brassolidae, Acraeidae & Nymphalidae. Hill House Editions. 525 p. [ Links ]
__________., 1988.- Butterflies of the Neotropical Region. Parte IV. Nymphalidae & Satyridae. Hill House Editions. 877 p. [ Links ]
__________., 1994. Butterflies of the Neotropical Region. Parte VI. Riodinidae. Hill House Editions. 1096 p. [ Links ]
DELABIE, et al., 2000.-. Sampling effort and choice of methods: 145-154 (en) AGOSTI, et al., Ants: Standard Methods for measuring and monitoring biodiversity. Smithsonian Institution Press. Washington. 280 p. [ Links ]
DEVRIES, J. P., 1987.- The Butterflies of Costa Rica and Their Natural History. Papilionidae, Pieridae, Nymphalidae. Princeton University Press. United States. 327p. [ Links ]
ESCOBAR, F., 1994.- Excremento, coprófagos y deforestación en bosques de montaña al suroccidente de Colombia: Tesis, Universidad del valle, Departamento de Biología, Cali. 68p. [ Links ]
__________., 1997.- Estudio de la comunidad de coleópteros coprófagos (Scarabaeidae) en un remanente de bosque seco al norte del Tolima, Colombia. Caldasia, 19 (3): 419 – 430. [ Links ]
__________., 2004.- Diversity and composition of dung beetle (Scarabaeinae) assemblages in a heterogeneous Andean landscape. Tropical Zoology, 17: 123-136. [ Links ]
ESTRADA, C. & FERNANDEZ, F., 1999.- Diversidad de hormigas (Hymenoptera: Formicidae) en un gradiente sucesional del bosque nublado (Nariño, Colombia). Revista de Biología Tropical, 47 (1-2): 189-201. [ Links ]
FAGUA, G., 1996.- Comunidad de Mariposas y Artropofauna Asociada con el Suelo de Tres Tipos de Vegetación de la Serranía del Tairarà (Vaupés, Colombia). Una Prueba del Uso de las Mariposas como Bioindicadores. Revista Colombiana de Entomología, 22 (3): 143-151. [ Links ]
__________. 1999. Variación de las mariposas y hormigas de un gradiente altitudinal de la cordillera oriental (Colombia): 317-362. (en) AMAT, G.; ANDRADE-C., G. & FERNÁNDEZ, F. (eds.). Insectos de Colombia. Volumen II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras. No. 13. Editora Guadalupe Ltda. Bogotá, Colombia. 541 p. [ Links ]
FERNÁNDEZ, F. ,2003.- Subfamilia Myrmicinae: 307-330. (en) FERNÁNDEZ, F. (ed). Introducción a las hormigas de la región Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, Colombia. 398 p. [ Links ]
GARCÍA R., et al., 2002. - Mariposas comunes de la Cordillera Central de Colombia. Guía de campo. Impresora Feriva S. A. Cali, Colombia. 130 p. [ Links ]
GIL, B. D., 1991.- Dung Beetles in American Tropical Forest: 211-229. (en) HANSKI, I. & CAMBEFORT, Y. Dung Beetle Ecology. Princeton University Press. New Jersey, E. U. 481p. [ Links ]
HALFFTER, G. & MATTHEWS, E. G., 1966.- Natural History of dung beetles of the subfamily Scarabaeinae (Coleoptera: Scarabaeidae). Folia Entomologica Mexicana. Reprint Medical Books. México. 313p. [ Links ]
HALFFTER, G. & ARELLANO. 2002.- Response de Dung Beetle Diversity to Human- induced Changes in a Tropical Landscape. Biotropica, 34 (1): 144-154. [ Links ]
HOLDRIDGE, L. R., 1982.- Ecología basada en zonas de vida. Instituto Interamericano de Cooperación de la Agricultura. San José de Costa Rica. 216p. [ Links ]
HOWDEN, H. F. & YOUNG, O. P., 1981.- Panamian Scarabaeinae: Taxonomy, Distribution, and habits (Coleoptera, Scarabaeidae). Contributions of the American Entomological Institute, 18 (1): 204 p. [ Links ]
KING, J. R.; ANDERSEN, A. N.& CUTTER, A. D., 1998.- Ants as bioindicators of habitat disturbance: validation of the functional group model for Australias humid tropics. Biodiversity and Conservation, 7: 1627-1638. [ Links ]
LAMAS G. (ed.), 2004.- Atlas of Neotropical Lepidoptera. Checklist: Part 4. Hesperoidea – Papilionoidea. p. 439. [ Links ]
LUDWING, J. A. & REYNOLDS, J. F., 1988.- Statistical Ecology. Jhon Wiley & sons. San Diego, California. 337 p. [ Links ]
MEDINA, C. A.& LOPERA T., A., 2000.- Clave ilustrada para la identificación de géneros de escarabajos coprófagos (Coleoptera: Scarabaeinae) de Colombia. Caldasia, 22 (2): 299-315. [ Links ]
MEDINA, C.; ESCOBAR, F. & KATTAN, G. H., 2002.- Diversity and Habitat Use of Dung Beetles in a Restored Andean Landscape. Biotropica, 34(1): 181-187. [ Links ]
OLIVER, I. & BEATTIE, A. J., 1996.- Defining a cost- effective invertebrate survey: A test of methods for rapid assessment of biodiversity. Ecological Applications, 6 (2): 594-607. [ Links ]
PALACIO, E., 1999. Hormigas legionarias (Hymenoptera: Formicidae: Ecitoninae) de Colombia: 117-189 (en) AMAT, G.; ANDRADE-C., G. & FERNÁNDEZ, F. (eds.) Insectos de Colombia. Volumen II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras. No. 13. Editora Guadalupe Ltda. Bogotá, Colombia. 541 p. [ Links ]
PALACIO, E.; FERNÁNDEZ, F., 2003.- Clave para las subfamilias y géneros: 233-260. (en) FERNÁNDEZ, F. (ed) Introducción a las hormigas de la región Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, Colombia. 398 p. [ Links ]
PARRA H., M. L.; VARGAS, J. I. & TABARES P., M., 2000.- Mariposas de Manizales. Artes gráficas TIZAN Ltda. Manizales, Caldas. 117 p. [ Links ]
VALENCIA M., C. A.; GIL P., Z. N. & CONSTANTINO, L. M., 2005. - Mariposas diurnas de la zona central cafetera Colombiana. Guía de Campo. Cenicafé. Chinchina, Colombia. 244 pp. [ Links ]
VILLAREAL, et al., 2004.- Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de Inventarios de Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, Colombia. 236 p. [ Links ]
VITOLO, A., 2000.- Clave para la identificación de los géneros y especies phaenaeinas (Coleoptera: Scarabaeidae: Corpinae: Phanaeini) de Colombia. Rev. Acad. Colomb. Cienc. 24 (93): 591-601. [ Links ]
ZAR, J. H., 1996.- Biostatistical analysis. Prentice may. New Jersey. 480p. [ Links ]