










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.12 no.1 Manizales Jan./Dec. 2008
Julián A. Salazar E.1
*Recibido 27 de junio de 2007, aceptado 22 de mayo de 2008.
1MVZ. Museo de Historia Natural. Universidad de Caldas. Manizales, Colombia. julianadolfoster@gmail.com
Este trabajo pretende demostrar la gran importancia taxonómica que tiene el estudio de los palpos labiales en mariposas tropicales de la familia Charaxidae. Los caracteres más relevantes son discutidos por su examen bajo el microscopio electrónico de barrido. Se da información adicional relacionada con los diseños del patrón alar y hábitos de los adultos pertenecientes a géneros de reciente descripción.
Lepidóptera, palpos, clasificación, Charaxidae, nuevos géneros, neotrópico, taxonomía, hábitos, patrones alares, Reuter, Rydon.
The intention of this work is to demonstrate the taxonomic importance of the study of cephalic palpi in neotropical butterflies of the Charaxidae family. Relevant characteristics are discussed regarding their structures examined under the electron microscope. Additional information is given on alar design patterns and adult habits belonging to some genera that have been recently described.
Lepidoptera, palpi, classification, Charaxidae, new genera, neotropical, taxonomy, habits, alar patterns, Reuter, Rydon.
INTRODUCTIONIn the taxonomic field, the Lepidoptera, especially the suborder Rhopalocera has been recently receiving plentiful contributions centered around the description of new species and varieties; the creation of new genera from others that had acted as an "omnibus" containing numerous species, morphologically similar in their mature state, but different in taxonomic characters related to their immature states (MUYSHONDT, 2005): the anatomy of the adults' alar venation; or the structure of the genital armor present in both sexes. Such topics have been treated by diverse authors (GOSSE, 1883; MÜLLER, 1886; CHAPMAN, 1916; SNODGRASS, 1935; FORBES, 1939; HERRERA & BEIRNE, 1942; HOWARTH, 1966; STEKOL´NIKOV, 1967; RYDON, 1971). Equally, other structures of outstanding taxonomic implication have been studied, (JORDAN, 1923; TYLER, BROWN & WILSON, 1994; ROBBINS, 1989) that also involve the suborder Heterocera (SNODGRASS, 1935; HEPPNER, 1993; SCOBLE, 1995; OLIVARES & ANGULO, 1996). The case of certain groups of neotropical butterflies, such as Riodinidae or Lycaenidae, have not been unaware of this phenomenon centered around phylogenetic analyses, whose basis is the study of the eggs, the sternites, the genitalia and the setae of immature states (JOHNSON et al., 1997; CALLAGHAN, 2000; D'ABRERA & BÁLINT, 2001; HALL, 2001, 2002, 2003; HALL & HARVEY, 2001; HALL & CALLAGHAN, 2003).
RECORDSA character that is still considered in these publications is the examination of the cephalic palpi located in the entrance to the mouth, organs that have not received appropriate treatment and whose meticulous attention was due to the pioneer works carried out by Enzio REUTER in 1896 and 1898 (Fig. 1). This author studied the variation of the length of the segments of the labial palpi in a comparative way, comparing the existing ones in many genera of butterflies; all of this, to discover certain patterns that repeat in representatives of different families, especially, in the morphometry of the joints or segments. According to VILORIA (2001), Reuter found correspondence in the data of his morphological studies used to differentiate certain sections within the families of butterflies, which led him to devise a taxonomic classification model and to propose a number of tribes. Such an effort was soon modified since he worked with a limited quantity of taxons that facilitated the generation of his conclusions. For groups of butterflies such as the Satyridae, the study of the lengths of the palpi segments is quite variable and it does not allow the subtraction of characters of important taxonomic value at any hierarchical level, mainly to establish homologies, since said structures could have been originated in multiple and independent ways in the cladogenesis of the Rhopalocera (VILORIA, Op. cit). Nevertheless, in recent times, several authors have ventured to rescue the value of the palpi as an important part within anatomical studies for some genera and species of neotropical origin (ORFILA, 1950; RYDON, 1971; CASAGRANDE, 1979; BILOTA, 1993; BRAVE et al., 2003; MIELKE et al., 2004), although they have not been considered as substantial factors in more modern taxonomic classification proposals (vg. SCOTT, 1985; HILL, 1990; NIJHOUT, 1991; MILLER & WENZEL, 1995; ACKERY et al., 1999; BROWER, 2000; VANE-WRIGHT, 2001; WILLMOTT, 2004; FREITAS & BROWN, 2004; WAHLBERG et al., 2005), the palpi are superficially included in the analysis of certain groups of Rhopalocera and Heterocera but referred to as an anatomical character considered close to venation and the forelegs (HEPPNER, 1993; SCOBLE, 1995; HALL, 1999a; CRANSTON & GULLAN, 2002; VANE-WRIGHT, 2003).

The Charaxidae (RYDON, 1971; SALAZAR & CONSTANTINO, 2001) is a group of tropical cosmopolitan butterflies that can be distinguished, according to ACKERY, DE JONG and VANE-WRIGHT (1999), due to the shared characteristic homologous conditions: partial parapatagia or entirely sclerotized and shortly anastomosed Rs3 and Rs4 venation (RYDON, 1971).
This article seeks to highlight the importance of the palpi in the diagnosis of some genera of neotropical Charaxids of recent description (SALAZAR & CONSTANTINO, 2001), by comparing certain regions of these organs between similar groups. Said factor was not considered as outstanding when being described.
MATERIALS & METHODSThe basis of this study was the extraction of palpi of 15-20 samples of species belonging to Rydonia, Annagrapha, Pseudocharaxes, Muyshondtia and Zikania, genera described by SALAZAR & CONSTANTINO (2001). Recently, LAMAS (2004) unified the first two genera as Memphis (Hübner, 1819). The remaining ones were fused in Polygrapha, Staudinger (1887). In spite of this approach and for effects of the present work, the generic status of such entities is conserved, also based on the conclusions collected by RYDON (1971), NEILD (1996) and that, COMSTOCK (1961) ended up seeing. It should be then clarified, that this posture has not been considered (or ignored) by several modern authors (HARVEY, 1991; BROWER, 2000; FREITAS & BROWN, 2004).
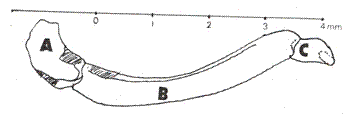
Additionally, the pencilled drawings previously made by A. H. B. RYDON in 1970 (Fig. 2) a pioneer on the systematic of Charaxids, using 25 neotropical species are included in this work (RYDON, Op. cit.). The isolated palpi underwent an extreme magnification achieved with the help of a sweeping electron microscope (Spot Magr. Acc. V type), in order to analyze certain basic structures. On the other hand, coloration patterns and adult habits were studied, taken in live samples (in situ) by means of a Pentax K-1000 camera.

A. CONFIGURATION AND DIFFERENTIATION OF THE LABIAL PALPUS
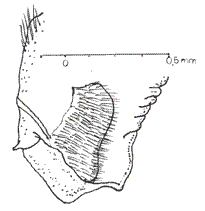
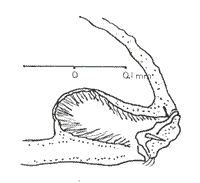
The palpi in butterflies are cephalic appendages relatively conspicuous associated with the external part of the oral sucking apparatus, as well as being, invariably, made up of three sequential articulated segments, or joints (SNODGRASS, 1935; EHRLICH & EHRLICH, 1961) (Fig. 3). The first segment, the basal one, is usually as long as it is wide in proportion; the second is almost always long and arched; and, the third segment, having somewhat of a lobe-like appearance, is as long as it is wide, similar to the basal joint. In the neotropical environment, the labial palpi in the Charaxidae have been studied in detail by several authors: ORFILA (1950) analyzed the Prepona Bsd. (sensu lato) genus; RYDON (1971) in his global analysis of the Charaxidae and more recently by MIELKE et al. (2004) studying the external anatomy of Zaretis (Hbn) and Agrias (Dbl.) (Figs. 4 & 5). However, as it was already mentioned, almost 110 years ago the German naturalist E. Reuter examined the palpi thoroughly, proposing a phylogenetic system for some butterfly groups (ACKERY, 1984). Reuter, when studying the anatomical characteristics of the palpi, established a series of differences centered on certain regions of the joints, especially the basal and the apical ones. Within the genera of the family Charaxidae, elevated to this rank by GUENÈE (1865), Reuter had at hand the palpi of the neotropical Hubnerian genera Hypna, Siderone, Zaretis and Anaea.


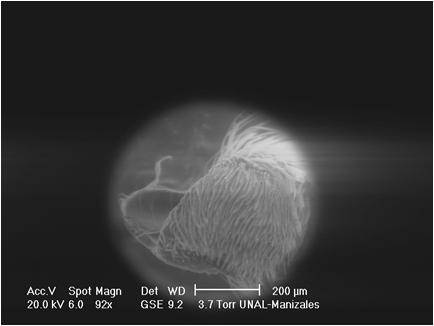
This last one based on "Anaea" iphis Latreille, 1813, that according to COMSTOCK (1961), belongs to the Memphis Hübner genus, as well as three Prepona Bsd. species that were placed by FRUHSTORFER (1916) in his Archaeoprepona genus. For the purpose of this work, the joints of the palpi will be analyzed by focussing on their more distinctive regions highlighted in the following manner:
1. Reuter´s sensitive patch:
It is an area located in the internal face of the basal joint near the union with the posgena, it is also called the "Basalfleck" or "sensorial area" (REUTER, Op. Cit; ORFILA, 1950). It is arranged as a plate provisioned with erect pectinate bristles (Fig. 6).
2. Second joint:
Basal projections of frontal hairs.
3. Gustatory crypt:
It is present in the third or last segment or joint. There is also a well-known fissure named "vom Rath's organ" (Fig. 7). According to REUTER (Op. cit.), peculiar dermal appendages also appear, more or less conical in form, and with tiny cavities or grooves. Such formations suggest sensorial organs similar to those described by other authors (Kreepelin, Foren, Flauser).
The sensitive patch is quite uniform in the diverse groups of Heterocera, but the Rhopalocera show great variation in its extension, form and in the level of development of its chitinous appendages.
In his beginnings, Reuter took into account three factors for palpi comparison:
a. The external form of the whole palpus.
b. The scale covering of the palpus.
c. The sensitive basal patch or "Basalfleck".
The first two, especially the first one, are frequently employed as generic characters of smaller or auxiliary taxonomic value (see HALL & WILLMOTT, 1996; CALLAGHAN, 2000; HALL, 2001, 2002; HALL & HARVEY, 2001), while the form and disposition of the basal sensitive patch is a specialized condition (REUTER, 1898; RYDON, 1971). It also constitutes an important factor when trying to establish morphological comparisons since, as it was indicated above, it varies in form or in structure.
For purposes of this work, segments of the palpi with their more distinctive regions found in the genera described by SALAZAR & CONSTANTINO (2001) will be treated as follows:
I. Genus MUYSHONDTIA Salazar & Constantino, 2001
Type species: "Papia" tyrianthina Salvin & Godman (1868)
This genus was created to include a single species, P. tyrianthina, which mainly inhabits the joint Andean area of Peru and Bolivia. The conformation of the palpus (Fig. 8) has an erect appearance, projected exactly above the head. The basal joint according to Rydon (com. pers.), lacks cones or scales, showing a naked sensitive patch. The cones are restricted more toward the union with the posgena, the medium segment has a length three and a half times greater than that of the basal one.
It is almost dorsally free of scales, except toward the ventral region, that under electron microscopy exhibits some lengthened scales, clear ones, with other dark ones that are bifid in the tips (Fig. 8b). The appearance of these scales is thinner and narrower than those of the genus Pseudocharaxes.


II. Genus PSEUDOCHARAXES Salazar & Constantino, 2001
Type species: "Paphia" xenocrates Westwood, 1850
The genus Pseudocharaxes was created in order to include the Ps. xenocrates species located near the Amazon basin. The conformation of the palpus in this species (Fig. 9) is erect with alternate dark and light regions, and ventrally depressed. The Reuter´s sensitive patch is scutiform (shield-like) and distally pointed. The medium segment is thick, with a length three and a half times greater than that of the basal segment. Its scales are microscopically wider, bifid with rounded tips. They tend to be more lengthened toward the basal segment (Fig. 9b).
III. Genus "ZIKANIA" Salazar & Constantino, 2001
Type species: "Anaea" suprema Schaus, 1920
Monotypic genus designated to include the species A. suprema. Nevertheless, in agreement with LAMAS (2004: 225), the name of Zikania is not being used, since it was conceived originally by Borgmeier in 1925 to include species belonging to the order Diptera. Consequently I propose the name of PROZIKANIA repl. name that considers this species endemic to Brazil. The configuration of the palpus is marked (Fig. 10), lightly haired, of a dorsally dark brown tonality with dark or clear grooves in the ventral region.



Under the electron microscope the sensitive patch is present in two separate parts; the first, with tight and abundant cones similar to hair, are thin and equally long (Figs. 10b, 10c). The other part is more spaced, somewhat hollow and with short sensilla hair. The medium segment is three times as long as the basal segment. It exhibits a multitude of bifid hairs with blunt and not so pointed tips as in M. tyrianthina (Fig. 10d).


IV. Genus RYDONIA Salazar & Constantino, 2001
Type species: "Papia" pasibula Doubleday, 1849
This genus includes two Andean species that, according to SALAZAR & CONSTANTINO (2001: 351), were previously included in the Memphis genus. Both the palp of R. pasibula and of its relative, R. falcata Hopffer 1875, have short configuration, with dark or clear tonality in the back with ventrally longitudinal bands of scales. Its appearance is similar to the palpi of the Anaea Hübner genus but with a more notable "S" form (Fig. 11). The electron microscope shows a triangular-shaped sensitive basal patch with a series of lengthened pectinate cones (Fig. 11b), that are defined by an area of wide scales of three or four tips each (Fig. 11c). The medium segment is three times as long as the basal one, and the distal segment offers a blunt, obtuse aspect, replete with longitudinal and abundant scales that end in two or three tips separated by notches of variable depth (Fig. 11d).
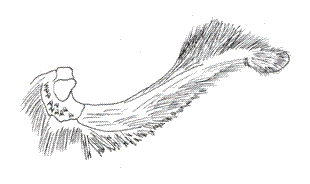
V. Genus ANNAGRAPHA Salazar & Constantino, 2001
Type species: "Paphia" aureola Bates, 1864
According to SALAZAR & CONSTANTINO (2001), this genus consists of five species separated from Memphis due to anatomical characteristics described in its original description, and that especially inhabit the humid tropical forests of America. The palpus in some of them, as for example A. anna (Stgr.), is of a dark color in the back and ventrally piebald with whitish scales. The sensitive patch of the basal segment is triangular-shaped with a nearby flat area. The medium segment is four times as long as the basal one and the distal segment has a blunt and obtuse appearance (Fig. 12). A. polyxo (Druce) has the same palpus configuration, but the sensitive patch is oblong, upholstered with cone-shaped sensilla (Fig. 13). With the microscopic examination, the palpi of the type species: A. aureola Bates present the sensitive patch provisioned with cone-shaped lengthened scales (Fig. 14), differentiated from the other typical basal and medium segment scales because they are wider, crossed with a longitudinal band that finishes off in two or three blunt and cut tips (Figs. 14b, c). This same type of scales is present in A. elina (Stgr.) (Figs. 15a, b). The distal segment is obtuse with bifid or trifid scales with variable width and length, but smaller toward the tip. Vom Rath's organ is imperceptible.



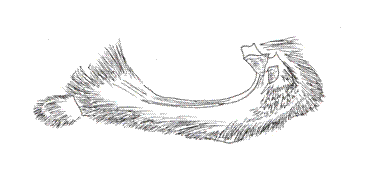
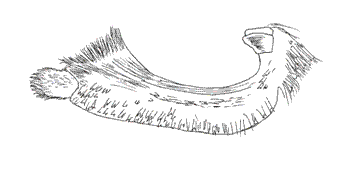

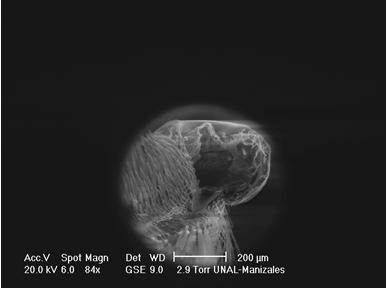



VI. Genus MEMPHIS Hübner, 1819
Type species: "Papilio" odilia Stoll (= polycarmes Fab.)
Twenty-five (25) species of this genus are registered for Colombia. The disposition of the palpal segments have been analyzed by Rydon (com. pers.), who studied the palpi of M. polycarmes (F.), M. boliviana (Dce.) and Fountainea (Memphis) sossipus (Latr.) (Figs. 16, 17, 18).
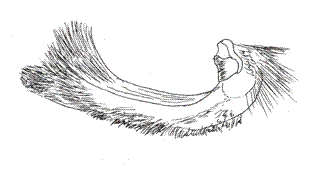
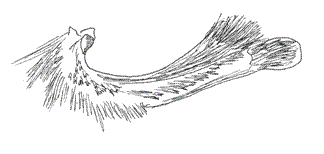
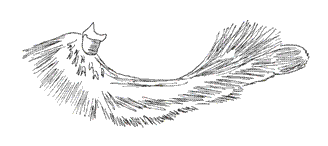
The configuration of the sensitive basal patch in these species is similar, of square shape. The medium segment is three and a half times as long as the basal one. The distal or terminal segment is obtuse and blunt. For the benefit of this paper, the microscopic aspect of the Memphis lyceus (Druce) palp is shown. Its sensitive basal patch´s disposition is square, with shorter scales than those of the Fountainea species (e.g. F. nessus (Latr.), for comparison) (Figs. 19, 20). The rest of the basal segment has similar scales to those of A. aureola, bifid or trifid crossed with more than 50 longitudinal grooves. The distal segment is obtuse with more lengthened and narrower scales, which are usually bifid in the tips (Figs. 21, 22).


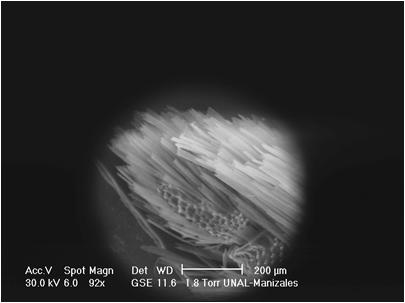

VII. Genus FOUNTAINEA Rydon, 1971
Type species: "Anaea" phidile (Geyer)
This genus was proposed by A. H. B. Rydon in 1971. It includes nine Colombian species. In two of the species analyzed by this author, F. ryphea Cr., and F. phidile Gy., the palpus configuration is similar: the basal segment shows a sensitive triangular patch. The medium segment is three and a half times longer than the basal one. The latter is oval and erected (Figs. 23, 24). Under the sweeping electron microscope, the palpi of F. nessus (Latr.), another species of Andean origin, are configured with a sensitive patch full of conical sensilla that are very lengthened and thinner than in other species, such as M. lyceus or A. aureola (Figs. 25, 26). Their insertion bases are curved and have a sequential disposition (Fig. 26b). The scales that underline the medium segment are longer and narrower and equally crossed with longitudinal bands.
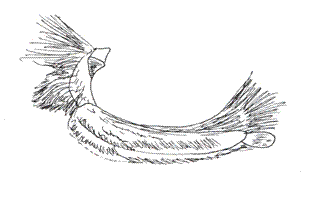
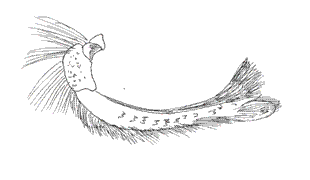


VIII. Genus POLYGRAPHA Godman & Salvin, 1887
Type species: "Paphia" cyanea Godman & Salvin, 1887
In a strict sense, Polygrapha only consists of P. cyanea species characteristic of the Amazon regions. Based on RYDON (1971: 340), this genus does not include "Polygrapha" suprema, "P." tyrianthina and "P." xenocrates, they have different genitalia, alar venations and the sensorial patches of the palpi. The palpus in this species has the Reuter sensitive patch, which is scutiform: wide and without the conical formations that are present in tyrianthina (Fig. 28). In suprema, they are limited to certain regions. The medium segment presents, abundant lengthened scales ventrally, which can be clear, dark and with bifid tips (Fig. 28b). Its length is three and a half times greater than that of the basal one. This last segment is obtuse and pointed.

Such differences, among others, noticed by RYDON (1971) were not considered by LAMAS (2004) in his Checklist of neotropical butterflies, who places Prozikania, (= Zikania), Muyshondtia and Pseudocharaxes in the same genus as Polygrapha. The same thing happens with Annagrapha, which was incorporated by said author in Memphis. Due to what was stated above, it is necessary to make other detailed studies, including DNA tests, in order to be able to elucidate the situation outlined by SALAZAR & CONSTANTINO (2001). Besides the immature stages are totally unknown in these new genera compared to those that are already well known and described by J. Hübner and A. Rydon.

B. SCALE STRUCTURE AND FORM
When attentively examining the palpi of certain Charaxids, three types of scales can be differentiated: two of them especially present in the medium and distal segments; the other, having a sensitive nature, stands out in the internal side of the basal segment (REUTER, 1896; MIELKE et al., 2004; BIZARRO et al., 2003). The first two types of scales (Figs. a and b) can vary in length and width, finishing in a bifid tip for those located in the medium and distal segments, while others are wide and with trifid tips in the dorsal area of the basal and medium segments. The cone-shaped scales are unique and are located within the sensitive patch. They are the principal characteristic with which REUTER (1896) tried to create a Rhopalocera genealogical tree, using the palpi of 302 genera and 670 species.
The scales almost cover the entirety of the palpal segments and they are often projected forward, similar to a fan, while others have a pectinate disposition, others that are more specialized exist in the distal segment. The latter have a special structure: the gustatory crypt (ORFILA, 1950), which occupies the central part of the third segment with an oral pore in the ventral part. In the superior margin, it has a lock of sensitive hair and laterally, a trichoid sensilla of tactile function. This specialized region is also known as vom Rath's organ (SCOBLE, 1995; MIELKE et al., 2004), which seemingly works as a kind of receiver for levels of carbon dioxide in the atmosphere (SCOBLE, Op. cit.). As it was said before, the form of the scale should be kept in mind, since Kellog's preliminary study (1894) is being used as a base. This is because he distinguished its structure as being hair, on one side, which is generalized in almost all the Rhopalocera, and on the other side, of being foliaceous and having a specialized appearance. In many cases, the variation in the shape of the scales is very minimal among groups of species phylogenetically related, mainly those of foliaceous type (VILORIA, 2001).
C. NOTE ON ADULT PATTERN COLORATIONThis topic has already been sufficiently treated by several authors, among which this work highlights DESCIMON (1983, 1986), NIJHOUT (1990, 1991), NIJHOUT and WRAY (1986), VILORIA (2001), etc. They had as a basis the preliminary works carried out by SCHWANWITSCH (1924, 1930a, 1930b, 1956), who established a basic Nymphalid type model that has worked as a fundamental explanation in order to know the coloration pattern and drawings of the butterflies and their possible derivative variations, understood as a diversification of bands and ocelli. According to DESCIMON (1986), for the Memphis genus that concerns the present study, the most frequent dorsal wing pattern is blue and black, with the exception of Cymatogramma and Fountainea which have a tawny bottom with black designs and violet gleams (F. nessus and allies). The ventral face of the Memphis possesses a characteristic design of fringes that acts as a deceiving appearance. According to VILORIA (2001), the above-mentioned is due to a stereo-effect phenomenon that substantially improves the cryptic defense of these butterflies when imitating a dry leaf or parts of a tree bark. These fringes are usually organized in a typical directed pattern almost the same in Memphis (sensu stricto). According to DESCIMON (1976), the ventral face of this genus belongs to the protective coloring pattern, or similar to a withered leaf. The deflective designs also appear on Fountainea and Polygrapha. The Memphis, Fountainea and Rydonia genera present an associate, primitive and static behavior when resting under the mid-day sun, when they settle on the leaves and branches of the trees and bushes. The genus Annagrapha is an exception, since it is involved in a singular Müllerian ring demonstrated by SALAZAR (2004), and interpreted by DESCIMON (Op. cit.) as a "pas évolutif" when imitating other genera (Callicore, Temenis). This happens in A. anna and A. elina, since A. aureola, polyxo and dia have developed a coloration pattern similar to that of certain Memphis.
D. NOTE ON THE HABITS AND ACTIVITIES OF FLIGHT IN ADULTSGood part of the Charaxidae species is assiduous visitors of the hill summits and other wild promontories that stand out of the environment in the diverse tropical landscapes from sea-level up to 2800 meters. This activity is best known as "Hilltopping", aids the males to fly over these emergent places to settle in alert exhibiting territorial behavior and to look for individuals of the opposite sex for mating (SCOTT, 1968; GUPPY, 1969; RUTOWSKI, 1984). Such places, according to ALCOCK (1987), mentioned by PINHEIRO (1990), generally do not harbor nutritious resources for adults and immature specimens, or they are not birth focuses of female virgins. Their only role is as a specialized habitat for sexual activities. In tropical areas, in order to have access to the study of these places it is necessary to ascend to the tops of trees with binoculars, or to use Rydon traps with bait and entomologic nets with very long handles (Figs. 29, 30, 31). In Colombia such habits of some Charaxidae species have been reported by SALAZAR (1996, 2001, 2002 and 2003). They can be categorized in the following way based on the observations carried out by PINHEIRO (1990) on Brazilian Papilionids:



1. Intra-specific Aggression:
• Observed among individuals of the same species.
• It consists of persecutions, fights in the air and falls toward the ground in twirls as it occurs in Memphis lyceus Druce and Rydonia pasibula Dbl. (*)
2. Inter-specific Aggression:
• It consists of single persecutions among individuals of different species, and it is usually quicker in comparison to the previous one. This has been verified in Fountainea nessus Latr., and F. glycerium Dbl., in the coffee-growing areas.
3. Restricted patrolling/Innkeepers:
• The flight carried out inside a small and well-defined area of the Hilltop. The great majority of the Charaxidae are included, since, although they are vigorous flyers, they have dominance over a small area, which they watch over with aggressive attitude over the most excellent foliage in the summits.
4. Twirls/Circled-flight:
• Carried out when drifting permanently over the place they have dominance of.
• The flight is done in a circular way.
• It is more frequent in the Papilionidae than in Charaxidae.
(*) A Note by A.H.B. Rydon:
The Behaviour of certain African Charaxids- Nymphaloids in the Field
"I was standing on top of a rocky hill in South Africa in April 1970 with a friend standing just below me with a long-handled net in his hand, when we saw two males of Charaxes pelias Cr., whirling rapidly up the hill-side as they battled with each other over territorial rights. When they reached where we were standing my friend made a sweep of his net and, much to our surprise, netted both males! Another African Charaxes, notoriously difficult to capture on the wing, is the attractive Ch. lactetinctus Karsch, the males of which are sharp-eyed and very aggressive towards other species of Charaxes and also towards birds, and when two males fight over territory they will go whirling upwards in decreasing circles until they are almost out of sight (Some collectors, in desperation to obtain specimens of lactetinctus, end up by shooting them down with reduced bird-shot.). In contrast to this aggressiveness in some, but no all Charaxids, are members of the African genus Euxanthe Hübner which are large, round-winged, forest butterflies. Two males will often be seen to pass many hours flying together to and fro at 2 or 3 m above the ground, performing a kind pf pas-de-deux. As these males often share the same tree in a forest, it is more likely that they are playing rather than fighting. There are also some other African Nymphaloid genera that are aggressively territorial among such being Cymothoe Hübner and Pseudathyma Hübner (both Limenitids). These two taxa have the convenient habit (for collectors, that is) of descending earthwards when their males battle over favorite perches on understory trees in ever decreasing circles, reminiscent of the neotropical Rydonia pasibula Dbl. and Memphis lyceus Druce, as reported above in this paper".
The species recognized as territorial and studied in the present work include the following:
Subtropical Humid forest (coffee belt)
• Memphis lyceus (Druce, 1877) (Fig. 32).
• Memphis philumena indigotica (Salvin, 1869).
• Cymatogramma perenna austrina (Comstock, 1961) (Fig. 33).
• Cymatogramma xenocles fissilis (Hall, 1935).
• Fountainea glycerium comstocki (Witt, 1972) (Fig. 34).
• Fountainea centaurus (C. & R. Felder, 1867).
• Fountainea nessus (Latreille, 1813).
• Rydonia pasibula (Doubleday, 1849) (Fig. 35).
Tropical Humid forest (Biogeographical Chocó)
• Memphis laura caucana (Joicey & Talbot, 1922).
• Memphis cleomestra (Hewitson, 1869).
• Memphis offa gudrun (Niepelt, 1924).
• Annagrapha aureola (Bates, 1866) (Fig. 36).
• Annagrapha dia (Godman & Salvin, 1884).
• Annagrapha anna elina (Staudinger, 1897).
The butterflies indicated above, include fruit-eating species, which are stratigraphically distributed in the canopy of the forest and the understory, just as it was verified by DE VRIES (1988) for five species of Costa Rica: cleomestra, laura, aureola, morvus and xenocles; showing their predilection for the tree tops (canopy), and very little for the understorey. Equally, this distribution has been studied by FERMON et al. (2000, 2003), and WILLIAMS (1973) in African Charaxids.





Another interesting phenomenon that can be applied to the Charaxidae of the New World is that some species present "Niche Segregations", as demonstrated by PRIETO (2005) and PRIETO & DAHNERS (2006) in detailed studies on Lycaenids (hairstreaks butterflies) that fly over the Western Colombian Mountain Range. These authors conclude that there is an elevated overlap in the daily spatial distribution of the species, showing competition preferences toward the mid-day. Although the Charaxidae behavior of this habit has not been well-analyzed yet, significant differences can present themselves in the use of the resources or segregation. The above mentioned can be due, not only to the inter-specific competition, but to each species' biological factors, such as the thermoregulation that goes according to the size of the species and the environmental temperature when they are activated to fly; or even, the host plants located at different heights in relatively close places to the niche (BLACKISH, 2005). The environmental temperature is what mostly defines the activity spectrum and depends on the presence of the sun (SALAZAR, 1996).
CONCLUSIONS• The analysis of the palpi in butterflies should be taken into account at the taxonomic level, in order to specify some degree of affinity or difference among some genera of neotropical origin of uncertain kinship.
• Nevertheless in the above-mentioned, the study of the morphometry as such, does not lead, in great measure, to the establishment of a strong form homology, since according to VILORIA (2001), the labial palpi could have been originated in a multiple and independent way in the Rhopalocera cladogenesis.
• The patterns of alar coloration in Charaxidae are the product of an evolutionary mimetic adaptation that seeks to resemble the environment they inhabit, or to simulate color patterns that appear in other species of unpleasant flavor for their predators.
• The adult habits in these butterflies, within their ecological niche, probably obey a use of the resource related to the segregation. This involves species or individuals that are limited to a territorial area of small extension, as occurs in the Lycaenidae family.
ACKNOWLEDGEMENTSThis article was greatly improved thanks to useful suggestions, criticisms, comments on the manuscript and financial support from Dr. Arthur H. B. Rydon (Sussex, UK). I thank Mauricio Arroyave, Rogelio Ospina and Dr. Alfonso Devia Cubillos () at Fisica del Plasma Laboratory (National University of Manizales, COLOMBIA) for expert microscope assistance. Thanks are also due to José I. Vargas, Gabriel Rodríguez (Envigado) and Dr. Olaf H. Mielke (Universidade Federal do Paraná, BRAZIL) for company and butterfly palps and specimens.
BIBLIOGRAPHY ACKERY, P. R., DE JONG, R. & VANE-WRIGHT, R. I., 1999.- 16. The Butterflies: Hedyloidea, Hesperioidea and Papilionoidea: 263-300. (in) KRISTENSEN, N. (ed.) Lepidoptera, Moths and Butterflies, vol. 1: Evolution, Systematics and Biogeography. Walter de Gruyter, Berlin, N.Y. [ Links ]
BEIRNE, B. P., 1942.- The morphology of the male genitalia of the Lepidoptera. The Ent. Record, 15 (3): 17-22, 37-39. [ Links ]
BIZARRO, J. M., CASAGRANDE, M. M. & MIELKE, O. H., 2003.- Morfología externa de Thyridia psidii cetoides (Rosenberg & Talbot). I. Cabeca e apendices (Lep: Nymphalidae, Ithomiinae). Rev. Brass. Zool., 20 (2): 279-284. [ Links ]
BROWER, A. V. Z., 2000.- Phylogenetic relationships among the Nymphalidae (Lep.) inferred from partial sequences of the wingless gene. Proc. Royal Soc. of London, 267: 1201-1211. [ Links ]
CALLAGHAN, C. J., 2000.- The genus Hyphophylla Boisduval, 1836 (Riodinidae) with descriptions of new taxa. J. Lepid. Soc., 54 (4): 119-130. [ Links ]
CHAPMAN, J. A., 1916.- VI. On the pairing of the Plebeiidi blue butterflies (Lycaenidae: Plebeiidi). Trans. Ent. Soc. London, part I: 156-180, pls. LXII-XVIII. [ Links ]
DESCIMON, H., 1986.- L'évolution de la coloration chez les Charaxidae nèotropicaux: strategies, adaptatives et cladogenese (Lepidoptera: Rhopalocera). Bull. Soc. Ent. Fr., 111: 261-296. [ Links ]
DE VRIES, P. J., 1988.- Stratification of fruit-feeding Nymphalid butterflies in a Costa Rican rainforest. J. Res. Lepid., 26 (1-4): 98-108. [ Links ]
EHRLICH, P. & EHRLICH, A., 1961.- How to Know the Butterflies: 2 W. C. Brown Comp. Publ. Dubuque, Iowa. [ Links ]
FERMON, H., WALTERT, M., LARSEN, T. B., DALL, U. & MÜHLENBERG, M., 2000.- Effects of forest management on diversity and abundance of fruit-feeding Nymphalid butterflies in South-eastern Cónte d'Ivoire. J. Insect Conservation, 4: 173-189. [ Links ]
FERMON, H., WALTERT, M. & MÜHLENBERGT, M., 2003.- Movement and vertical stratification of fruit-feeding butterflies in a managed West-African rainforest. J. Insect Conservation, 7: 7-19. [ Links ]
FORBES, W. T. M., 1939.- The muscles of the Lepidopterous male genitalia. Ann. Ent. Soc. Amer., 32 (1): 1-10. [ Links ]
FREITAS, A. V. & BROWN, K. S., 2004.- Phylogeny of the Nymphalidae (Lepidoptera). Systematic Biology, 53 (3): 363-383. [ Links ]
GOSSE, P. H. 1883.- On the clasping-organs ancillary to generation in certain groups of the Lepidoptera. Trans. Linn. Soc. London, 2 (6): 265-345, pls. 26-33. [ Links ]
GUPPY, R., 1969.- Further observations on "Hilltopping" in Papilio zelicaon. J. Res. Lepid., 8 (3): 105-117. [ Links ]
HALL, J. P., 2002.- A Review of the new Riodinid butterfly Genus Panaropsis (Lep: Riod. Symmachiini). Proc. Ent. Soc. Wash., 104 (1): 63-72. [ Links ]
HALL, J. P., 2002.- A Review of the new Riodinid butterfly Genus Dachetola (Lep: Riodinidae). J. NY. Ent. Soc., 109 (2): 183-195. [ Links ]
HALL, J. P., 2003.- A Phylogenetic reassessment of the five forewing radial-veined tribes of the Riodininae (Lep: Riodinidae). Systematic Entomology, 28: 23-37. [ Links ]
HALL, J. P. & HARVEY, D. J., 2001.- A Reassessment of Calociasma with the description of a new Genus and a new species (Lep: Riodinidae: Nymphidiini). J. NY. Ent. Soc., 109 (2): 196-205. [ Links ]
HALL, J. P. & CALLAGHAN, C. J., 2003.- A Revision of the new Riodinid butterfly Genus Pseudotinea (Lep: Riodinidae). J. Nat Hist. 37: 821-837. [ Links ]
HARVEY, D. J., 1991.- Appendix B. Higher Classification of the Nymphalidae: 255-273. (in) NIJHOUT, H. F. (ed.). The Development and Evolution of butterfly wing patterns. Smith. Inst. Press. [ Links ]
HEPPNER, J. B., 1993.- Keys to families of Lepidoptera. Tropical Lep. Suppl., 3: 1-28. [ Links ]
HERRERA, J. & HOWARTH, T. G., 1966.- Genitalia de los tipos de Satyridae de Chile depositados en el British Museum. Publ. Centr. Est. Ent: 73-126. [ Links ]
JORDAN, K., 1923.- On a sensory organ found on the head of many Lepidoptera. Nov. Zool., 30: 155-158. [ Links ]
LAMAS, G., 2004.- Charaxinae: 224-234 (in) Atlas of Neotropical Lepidoptera. Checklist: Part 4A Hesperioidea- Papilionoidea: 439 pp. Ass. Trop. Lep. Scient. Publ. Gainesville, Fl. [ Links ]
MIELKE, C. G., MIELKE, O. H. & CASAGRANDE, M. M., 2004.- Estudo comparado da morfologia externa de Zaretis itys itylus (Westwood) e Agrias claudina anneta (Gray) (Lep: Nymphalidae, Charaxinae). I Cabeca, apèndices e regiao cervical. Rev. Bras. Zool., 21 (2): 357-369. [ Links ]
MILLER, J. S. & WENZEL, J. W., 1995.- Ecological characters and Phylogeny. Ann. Rev. Ent., 40: 389-415. [ Links ]
MUYSHONDT, A., 2005.- Notas sobre el ciclo y la Historia Natural de algunas Mariposas de El Salvador: 455 pp. + figs. Impr. Universitaria U. de El salvador, San Salvador. [ Links ]
OLIVARES, T. S. & ANGULO, A. O., 1996.- El Órgano Timpánico en la clasificación de Lepidóptera: Noctuidae. Bol. Ent. Venez. N.S., 11 (2): 155-183. [ Links ]
OTERO, L. D., 1990.- Estudio de algunos caracteres para su uso en la clasificación de Eurytelinae (Lep: Nymphalidae). Bol. Ent. Venez. N.S., 5 (16): 123-138. [ Links ]
ORFILA, R. N., 1950.- Las especies Argentinas de "Prepona" Boisduval (Lep: Nymph.) Rev. Inst. Nac. Invest. Cienc. Nat. Mus. Arg. "Bernardino Rivadavia" 1 (7): 273-321. [ Links ]
PINHEIRO, C. E., 1990.- Territorial Hilltopping behavior of three swallowtail butterflies (Lep: Papilionidae). J. Res. Lepid., 29 (1-2): 134-142. [ Links ]
PRIETO, C., 2005.- Segregación de Nicho en una comunidad de Licénidos de montaña del Valle del Cauca (Lep: Lycaenidae, Theclinae). XXXII. Congr. Soc. Col. Ent. Resúmenes: 58. Ibagué, Tolima. [ Links ] PRIETO, C. & DAHNERS, H., 2006.- Sección morfología, comportamiento, ecología, y sistemática de Eumaeini (Lep. Lyc.) del cerro San Antonio. Dinámica de la riqueza y comportamiento. Rev. Col. Ent., 32 (2): 179-187 + pls. [ Links ] REUTER, E., 1896.- Über die Palpen der Rhopaloceren ein beitrag zur erkenntnis der verwandtschaftlichen beziehungen unter den Tagfaltern. Acta Soc. Scientiarum Fennicae, 22 (1): xvi + 1-577 & V. Taffs. [ Links ]
REUTER, E., 1898.- On a New classification of the Rhopalocera. The Entomologist´s Rec. J. Var., 10 (2): 25-26, 75-77, 95-98 + 1 pl. [ Links ]
ROBBINS, R. K., 1989.- Systematic implications of butterfly leg structures that clean the antennae. Psyche, 96 (3-4): 209-222. [ Links ]
RUTOWSKI, R. L., 1984.- Sexual selection and the evolution of butterfly mating behavior. J. Res. Lepid., 23 (2): 125-142. [ Links ]
RYDON, A. H. B., 1964.- Notes on the use of butterfly traps in East Africa. J. Lepid. Soc., 101: 51-58, 4 figs. [ Links ]
RYDON, A. H. B., 1971.- The Systematics of the Charaxidae (Lep: Nymphaloidea). Entomologist's Rec., 83: 219-233, 283-287, 310-316, 336-341, 384-388. [ Links ]
SALAZAR, J. A., 1996.- Sobre la concentración de Lepidópteros Rhopaloceros en la cumbre de un cerro del Noroccidente de Caldas, Colombia (Insecta: Lepidóptera). Shilap, 24 (94): 183-195. [ Links ]
SALAZAR, J. A., 2001.- Nuevas Observaciones sobre la concentración de Lepidópteros Rhopaloceros en las cumbres de algunos cerros de Caldas y Antioquia, Colombia (Insecta: Lepidóptera). Bol. Cient. Mus. Hist. Nat. U. Caldas, 5: 66-90. [ Links ]
SALAZAR, J. A., 2004.- Patrones de coloración en algunos Charaxidae de Colombia y sus modelos Mullerianos (Lep: Nymphaloidea). Lambillionea, 104 (1): 109-120. [ Links ]
SALAZAR, J. A. & LÓPEZ, C., 2002.- Predicting the overall butterfly species richness in a tropical montane rainforest in the Colombian Chocó. Bol. Cient. Mus. Hist. Nat. U. Caldas, 6: 111-145. [ Links ]
SALAZAR, J. A., GIRALDO, M. & VARGAS, J. I., 2003.- Más Observaciones sobre la concentración de mariposas territoriales en cumbres de cerros colombianos y especies residentes en el Bosque de Bavaria (Villavicencio, Meta) su distribución espacial y trofismo. Bol. Cient. Mus. Hist. Nat. U. Caldas, 7: 255-317. [ Links ]
SALAZAR, J. A. & CONSTANTINO, L. M., 2001.- Synthesis of the Colombian Charaxidae and description of new Genera for South America: Rydonia, Annagrapha, Pseudocharaxes, Muyshondtia, Zikania (Lep: Nymphaloidea). Lambillionea, 101. Suppl. 2: 344-369. [ Links ]
SCHWANWITSCH, B. N., 1930.- Studies upon the wing-pattern of Prepona and Agrias two genera of South American Nymphalid butterflies. Acta Zool., 11: 290-421+ 4 pls. [ Links ]
SCHWANWITSCH, B. N., 1930b.- Wing pattern of Cattagramma and related South American Nymphalid butterflies. Trans. Zool. Soc. London, 21 (2): 186-286 + 12 pls. [ Links ]
SCOBLE, M. J., 1995.- The Lepidoptera: Form, Function and Diversity: 404 pp. The Nat. Hist. Mus. And Oxford Un. Press. [ Links ]
SCOTT, J. A., 1968.- Hilltopping as a mating mechanism to aid the survival of low density species. J. Res. Lepid., 7 (4): 191-204. [ Links ]
SCOTT, J. A., 1984.- The Phylogeny of Butterflies (Papilionoidea and Hesperioidea). J. Res. Lepid., 23 (4): 241-281. [ Links ] SNODGRASS, R. E., 1935.- Chapter VII. The Head Appendages: 130-155 (in) Principles of Insect Morphology. McGraw Hill Book comp. Inc. NY. London. [ Links ]
STEKOL'NIKOV, A., 1967.- Phylogenetic relationships within the Rhopalocera on the basis of the functional morphology of the genital apparatus. Ent. Rev., 46 (1): 1-11. [ Links ] VANE-WRIGHT, R. I., 2001.- Taxonomy, Methods Of (in) Encyclopedia of Biodiversity, 5: 589-606. Academic Press, London. [ Links ]
VILORIA, A. L., 2001.- Sistématica práctica: recuento de morfología externa, reconocimiento y uso de las homologías en los Rhopalocera adultos (Insecta: Lepidóptera): 15 pp. + figs. 1er. Dipl. de Marip. Trop. U. del Valle, Cali, Colombia. [ Links ]
WILLIAMS, J. G., 1973.- A Field Guide to the Butterflies of Africa: 238 pp. + 24 pls. Collins clear, London. [ Links ]
WILLMOTT, K., 2004.- Brush-footed Butterflies (Lep: Nymphalidae) (in) CAPINERA, L. (ed.) vol. 1A-E: 379-384. Dordrecht-London, Kluw. Ac. [ Links ]
