










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.13 no.1 Manizales Jan./june 2009
1 Semillero de Investigación en Biodiversidad y Recursos Fitogenéticos, Facultad de Ciencias Agropecuarias, Universidad de Caldas. A.A 275. E-mail: valentina-r-v@hotmail.co
2 Herbario Universidad de Caldas (FAUC), Grupo de Investigación en Biodiversidad y Recursos Fitogenéticos, Facultad de Ciencias Agropecuarias, Universidad de Caldas. E-mail: david.sanin@ucaldas.edu.co, luis.alvarez@ucaldas.edu.co
3 Corporación Flora de Caldas. E-mail: corporacionfloradecaldas@yahoo.com
Resumen
Con el objetivo de analizar el crecimiento de helechos arbóreos en la Cordillera Central, entre marzo de 2006 y febrero de 2007 se realizó el monitoreo de la población de Dicksonia sellowiana en la Reserva Río Blanco (Manizales, Caldas), registrando trimestralmente el DAP, la altura total, el número de hojas emitidas y su estado reproductivo. Esta información se relacionó con la precipitación y se comparó con los ejemplares depositados en los herbarios COL, HUA y FAUC. El diámetro se incrementó en 1,2 cm, especialmente en la primera temporada del 2007. El crecimiento en altura del tallo fue de 6,7 cm, con la misma tendencia. La emisión de hojas se mantuvo estable, permaneciendo las 9 hojas iniciales. Únicamente las plantas con alturas mayores a 4 m se encontraron fértiles. Se concluye que D. sellowiana responde a un patrón fenológico estacional bimodal determinado por la precipitación, y coincide con la mayor esporulación registrada en los ejemplares de herbario. Igualmente, se encontró que el crecimiento en el tallo fue similar a otras especies de Cyatheales, principalmente en Alsophila. La emisión y acumulación de las hojas se presentó en módulos continuos "duplos foliares", lo que representa una estrategia adaptativa en los ecosistemas alto andinos, donde la materia orgánica acumulada y de lenta descomposición estabiliza el microclima. Estos argumentos aportan bases para el desarrollo de programas de conservación a partir del estudio y el manejo sostenible de las poblaciones naturales.
Palabras clave: Estimación del Crecimiento, Dicksonia sellowiana, Reserva Río Blanco, fertilidad en Colombia.
AbstractIn order to analyze the growth of tree ferns in the Cordillera Central, between March 2006 and February 2007, the population of Dicksonia sellowiana in the Rio Blanco Reserve (Manizales, Caldas) was monitored. The DAP, total height, leaf number and reproductive status were recorded quarterly. This information was related to precipitation and compared with specimens deposited in the COL, HUA and FAUC herbaria. The diameter increased by 1.2 cm, especially in the first 2007 season. The height growth of the stem was 6.7 cm, with the same trend. The leaf number was stable with 9 initial leaves. Only plants with a height greater than 4 m were fertile. The results showed that D. sellowiana answers to a bimodal seasonal phenological pattern determined by rainfall, coinciding with the increased sporulation recorded in herbarium specimens. Similarly, the growth in the stem was similar to other Cyatheales species, mainly Alsophila. The emission and accumulation of leaves were seen in continuous "double leaf" modules, an adaptive strategy in the high Andean ecosystems, where the accumulated organic matter of slow decomposition stabilizes the microclimate. These arguments provide a basis for the development of conservation programs from the study and sustainable management of natural populations.
Key words: Growth Estimation, Dicksonia sellowiana, Rio Blanco Reserve, fertility in Colombia.
INTRODUCCIÓNLa floración, fructificación y esporulación afectan los procesos de sucesión vegetal, principalmente por eventos reproductivos como la sincronización, polinización, dispersión, germinación y posterior patrón de distribución propiciado por el banco de semillas o esporas. En este sentido, los factores abióticos han sido pobremente registrados en relación al crecimiento y desarrollo de las plantas del Neotrópico (WRIGTH & CALDERÓN, 1995), limitando el entendimiento de muchos procesos que influyen en la regulación y dinámica de los bosques.
La fenología permite conocer la tasa de crecimiento y desarrollo y las fases de vida de las especies en relación con las características medioambientales. Dada la abundancia y visibilidad de las Angiospermas, estos estudios se han centrado principalmente en estas plantas (CROAT, 1975; MEHLTRETER & RÍOS-PALACIOS, 2003), debido a que presentan considerables tasas de crecimiento, mayor diversidad y estructuras reproductivas conspicuas "las flores" (MEHLTRETER & RÍOS-PALACIOS, 2003). Por otro lado, algunos estudios realizados en Monilophyta (helechos verdaderos) (SEILER, 1981, 1995) registran tasas lentas de crecimiento, su diversidad representa el 10% de la riqueza de cualquier flora en el neotrópico (KRAMER, 1976) y la producción de estructuras reproductivas en los helechos (soros, esporangios y esporas) es poco vistosa. De manera contrastante, existen especies que presentan rápido crecimiento, generalmente no dependen de polinizadores y de vectores animales para su dispersión (MEHLTRETER & RÍOS-PALACIOS, 2003). Así, los factores abióticos juegan el rol más importante en los patrones de temporalidad fenológica (MEHLTRETER & RÍOS-PALACIOS, 2003).
Existen estudios en fenología de helechos arborescentes, principalmente de Cyatheaceae J.S. Kaulf. (CONANT, 1976; TANNER, 1983; ASH, 1987; SEILER, 1981, 1995; BITTNER & BRECKLE, 1995; SCHMITH & WINDISCH, 2006), los cuales se han enfocado en la generación de programas de conservación, debido al peligro de extinción presente en estos grupos por la explotación comercial a la cual son sometidos (SCHMIT & WINDISCH, 2006).
Dicksonia sellowiana Hook. es un helecho arborescente nativo de Suramérica (MURILLO, 1988), habita ecosistemas silvestres localizados desde los bosques montanos y premontanos hasta el páramo (MORAN & RIBA, 1995). Su sistema radicular es característico del Orden Cyatheales (Monilophyta) por la ausencia de una raíz primaria axonomorfa, que es reemplazada por innumerables raicillas secundarias de origen adventicio, es decir, que surgen de la base del tallo (FONT-QUER, 1982). Constituyendo un sustrato ideal para la siembra de plantas epífitas conocido como "Sarro", por cuanto es permeable, permite el intercambio de gases, es aséptico dada su lenta descomposición (ELEÚTERIO, 2006), su alto contenido de taninos (KORALL et al., 2006) y sus intrincadas formas que brindan resistencia y belleza al sustrato (ELEÚTERIO, 2006).
Esto ha provocado que se realicen extracciones selectivas, donde la práctica consiste en el sacrificio de individuos adultos, cortando el sistema radicular y dejando el tallo y las hojas expuestas, con el desperdicio de estos órganos y la posterior disminución de las poblaciones naturales (CONSTANTINO et al., 1999). Por esto, actualmente en Colombia se reconoce bajo la categoría de Vulnerable (VU) (CALDERÓN, 1997). La Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES), la reconoce En Peligro (EN).
La información que se ha publicado para la especie en Colombia, se limita al tratamiento taxonómico realizado para Dicksoniaceae (C. Presl) Bower (MURILLO, 1988) y a experimentos dirigidos a la propagación masiva in-vitro (CONSTANTINO et al., 1999). No existen registros fenológicos para Dicksonia sellowiana, por lo tanto, aspectos reproductivos, de establecimiento y desarrollo, aún son desconocidos, lo que limita la generación de programas de conservación, uso y manejo sostenible (CROAT, 1975). Estos son la base para el diseño y la implementación de estrategias para el aprovechamiento de los recursos vegetales, en acuerdo con su conservación in situ y la interacción con su ecosistema (RICKER & DALY, 1997). Basados en estos argumentos, presentamos una comparación de la fenología de D. sellowiana, realizada en la Reserva de Río Blanco, con registros colombianos.
MATERIALES Y MÉTODOSDicksoniaceae (C. Presl) Bower, está compuesta por tres géneros y 30 especies (SMITH et al., 2006). Representa un grupo monofilético (KORALL et al., 2006), incluido en el Orden Cyatheales (SMITH et al., 2006), localizado evolutivamente junto con las familias Cyatheaceae Kaulf., Cibotiaceae Korall, Culcitaceae Pichi-Sermolli, Loxsomataceae C. Presl., Thyrsopteridaceae Korall, Plagiogyriaceae Bower y Metaxyaceae C. Presl. (KORALL et al., 2006). Orden que se distribuye desde el Este de Asia, Australia, el Neotrópico, hasta Santa Helena y los Estados Unidos (KORALL et al., 2006). Se caracteriza por la presencia de rizomas con dictiostelas policíclicas, con los ápices de los tallos y la unión de los pecíolos cubiertos por pelos uniseriados; láminas largas (mayores a 2 m.), 2-3 pinnadas, venas simples o divididas, libres; soros marginales y bibalvados; anillos de los esporangios oblicuos, receptáculos levantados, parafisos algunas veces presentes, filiformes y esporas globosas triletes (SMITH et al., 2006).
Dicksonia sellowiana Hook. (Tomado de MORAN & RIBA, 1995)
Helecho arborescente, tallo 5-8 (-10 m) x 0,15-0,3 m (incluyendo la densa cubierta de raíces adventicias); hojas 2-3 m, esparcidas a ascendentes; pecíolo de 10 a 30 x 1,5 cm, pardo oscuro a pajizo; lámina ampliamente lanceolada, reducida hasta el ápice y la base, coriácea a subcoriácea; raquis liso a ligeramente muricado, comúnmente pajizo, ligeramente peloso como el pecíolo; pinnas hasta 70 x 20 cm, subsésiles a cortamente pediculadas (ca. 1 cm), agudas a acuminadas; costa y cóstulas densamente pelosas adaxialmente, los tricomas principalmente pardo oscuro, rígidos y antrorsos, escasa a abundantemente pelosas abaxialmente, los tricomas amarillos a parduscos, rígidos, antrorsos a dispersos, últimos segmentos 3-5 x 2-5m, crenado-aserrados a profundamente lobados, glabros, no o ligeramente glaucos, ligera a fuertemente revolutos, el ápice agudo a cuspidado; nervaduras 5-7 (-8) o más pares por segmento, simples o bifurcadas, los segmentos profundamente lobados, 1-3 bifurcadas.
Dicksonia sellowiana se encuentra distribuida desde Centroamérica hasta el Norte de Uruguay, habitando ecosistemas de selvas bajas, bosques de niebla, bosques húmedos en laderas, quebradas, sotobosques, bosques de galería, matorrales achaparrados, bosques premontanos muy húmedos, paramos y bosques caducifolios, entre los 1900-3200 m (MORAN & RIBA, 1995). En Colombia, la especie ha sido registrada desde los 1500 hasta 3800 m (MURILLO, 1988), en los departamentos de Boyacá, Cauca, Cesar, Cundinamarca, Huila, Magdalena, Nariño, Putumayo, Santander, Norte de Santander y Tolima (MURILLO, 1988). ÁLVAREZ-MEJÍA et al. (2007) registran la presencia de la especie en el departamento de Caldas, precisamente en los bosques premontanos y montanos de las Reservas Río Blanco y Bosques de la CHEC.
Aspectos generales de la zona de estudio
La quebrada Olivares ha sido la fuente de agua para la ciudad de Manizales desde el primer cuarto del siglo pasado, por ello el municipio en 1904 inició la compra de predios en el área de influencia. Hacia 1951, cuando la zona protegida alcanzó las 1000 ha, Enrique Pérez Arbeláez presentó un plan de manejo y ordenación de la cuenca. En febrero de 1966 se sumaron a la reserva los predios: La Arenosa, Buenos Aires, Pinares, Martinica y La Elvira, con un total de 614 ha, donde se establecieron plantaciones forestales, predominando el cultivo de Aliso (Alnus acuminata Kunth). Hacia 1989 se otorga la propiedad a las Empresas Públicas y el 25 de julio de 1990 la Junta Directiva del INDERENA, mediante acuerdo No. 0027, crea la Reserva Forestal Protectora de las cuencas hidrográficas de Río Blanco y Olivares, avalada por el Ministerio de Agricultura mediante la resolución ejecutiva No. 66 de abril de 1992.
Descripción Biofísica
La Reserva Forestal Protectora de Río Blanco se ubica al Nororiente del municipio de Manizales, sobre la vertiente Occidental de la cordillera Central colombiana, entre los 2150 y 3700 m. Tiene un área aproximada de 4932 ha (Figura 1), comprende las veredas Alto del Guamo, Buenavista, El Desquite, El Paraíso, La Esperanza, Las Palomas, Río Blanco y Sinaí.
El clima en el área es típicamente intertropical, caracterizado por pequeñas oscilaciones de la temperatura interanual, pero con grandes fluctuaciones diarias y una distribución bimodal de la lluvia durante el año, influenciada por la Zona de Convergencia Intertropical (ZCIT) y por el carácter montañoso de la región (WITTE, 1995).
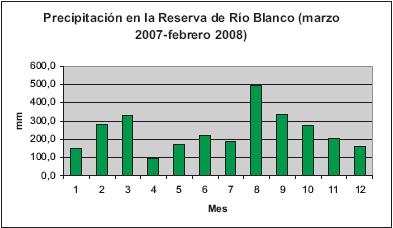
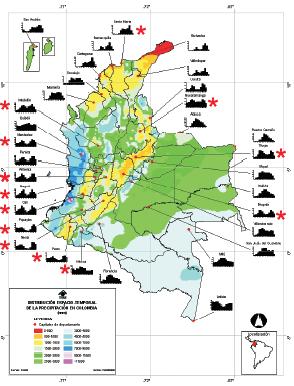
Según la información de la estación meteorológica "Las Palomas", a 2630 m de altitud la zona media de la Reserva presenta los siguientes promedios anuales de temperatura: Máxima (19°C), Media (10°C) y Mínima (6.9°C), donde los períodos más cálidos del año se encuentran en los meses de menor precipitación (diciembre - febrero y julio - agosto). El promedio anual de precipitación entre los años 1997 y 2001 fue de 2500 mm, y una humedad relativa del 80 al 90%. Los meses de mayor precipitación son abril y mayo, octubre y noviembre. Para la época en la cual se realizó este estudio, los meses con mayor precipitación fueron abril y mayo, en el primer semestre. Octubre y noviembre registraron los mayores valores de precipitación (Figura 2).
El suelo es de origen volcánico, con piroclastos Andesíticos del Cuaternario Tardío (Andisoles), según el sistema FAO/UNESCO (WRIGHT, 1964, citado por VIS, 1995). Y rico en materia orgánica (Altos contenidos de alófanas) y humíferos, con regímenes de humedad desde Údico hasta Ácuico.
La reserva se ubica sobre las siguientes formaciones vegetales: Selva Andina, Selva Subandina y Páramo (CUATRECASAS, 1958). Y según las zonas de vida, se localiza dentro del Bosque muy húmedo montano hasta el Sub-Páramo (HOLDRIDGE, 1985). La flora de la zona ha sido estudiada por SANÍN et al. (2006) y (SANÍN & DUQUE, 2006).
Aspectos metodológicos
En una localidad ubicada a los 5° 4' 3" N y 75° 25' 4" W, con estado sucesional avanzado dentro de la Reserva, se marcaron y monitorearon nueve individuos con alturas comprendidas entre los 1,8 hasta los 4,8 m, y diámetros entre los 0,09 y 0,168 m, donde a cada planta se la marcó con un número consecutivo, ubicado siempre hacia el Sur de cada localidad, registrando mensualmente para cada sitio la circunferencia a la altura del pecho (CAP) (medida siempre en el punto donde se encontraba el número de registro), la altura total, el número de hojas y su fertilidad. Los especímenes recolectados fueron enviados a los Herbarios Nacional Colombiano (COL), Universidad de Antioquia (HUA) y Universidad de Caldas (FAUC), bajo la serie SANÍN et al. (2006). Se realizaron tres mediciones durante los años 2006 y 2007, cada una con un lapso de tiempo de cinco meses (marzo, agosto y febrero).
Esta información se comparó con la precipitación, y se determinó un patrón de crecimiento. Complementariamente, se revisaron 51 ejemplares de Dicksonia sellowiana Hook., depositados en los herbarios COL, HUA y FAUC (Anexo 1), donde se procesó información acerca de su estado reproductivo, la localidad de registro, el recolector y la fecha.
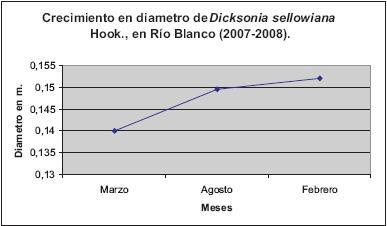
RESULTADOS Y DISCUSIÓNEl crecimiento en el diámetro de la población evaluada fue de 1,2 cm en un periodo de 11 meses, el mayor incremento se presentó en la primera temporada del año (Figura 3). No se encontraron estudios que registren el incremento en la circunferencia del diámetro de helechos arbóreos y su relación con los factores ambientales (SEILER, 1981, 1995; TANNER, 1983; BITTNER & BRECKLE, 1995; ARENS & SÁNCHEZ-BARACALDO, 1998; ARENS & SÁNCHEZ-BARACALDO, 2000). Esto limita el diseño de estrategias de manejo y aprovechamiento sostenible de las poblaciones de Dicksonia sellowiana. Así, este estudio presenta información que en el futuro puede ser comparable para tales fines. Para Angiospermas, se registran datos similares en plantaciones forestales, determinando un volumen que se calcula a partir de la altura fustal y el área basal (LEMA, 1995). Esta información y sus métodos de obtención pueden consolidar metodologías para la generación de registros en helechos arborescentes.
El crecimiento en altura de Dicksonia sellowiana fue de 6,7 cm en el periodo evaluado (11 meses). El primer periodo mostró el mayor incremento (Figura 4). Al comparar este registro con otros estudios realizados en Cyatheaceae, específicamente en Alsophila (A. bryophyla, A. erinacea, A. polystichoides y A. setosa), se registran valores similares (Tabla 1). En este sentido, A. setosa presentó dos promedios diferentes de crecimiento en el tallo, determinados por el estado sucesional en el cual crecían las poblaciones evaluadas, donde los individuos localizados en un ecosistema secundario muestran valores promedio de 14,51 cm, y los ecosistemas primarios de 6,32 cm en un periodo de un año (SCHMITT & WINDISCH, 2006).

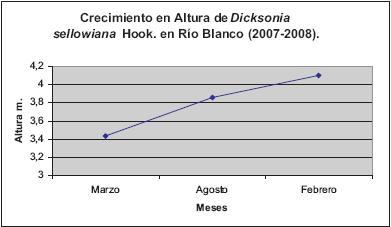
Por lo anterior, es necesario registrar la posible variación fenotípica de Dicksonia sellowiana, ya que ésta presenta considerable abundancia y densidad en ecosistemas abiertos, mientras que en bosques secundarios y primarios de la Planada-Nariño no registró niveles significativos para estas características ecológicas (ARENS & SÁNCHEZ-BARRACALDO, 1998), indicando que no solo la estrategia de distribución se puede alterar, sino también sus tasas de crecimiento y su morfología (SEILER, 1981; ARENS & SÁNCHEZ-BARRACALDO, 1998).
La emisión de hojas (Figura 5) se mantuvo estable, permaneciendo el mismo número de frondas iniciales y finales (9). Sin embargo, la segunda medición registró un promedio de siete hojas (Figura 6). En el estudio realizado en Alsophila setosa, se observó un promedio de 8,66 hojas al año en la localidad de Morro Reuter "ecosistema alterado", mientras que en el ecosistema comparado, donde el estadío sucesional era avanzado, denominado Sapiranga, el promedio fue de 6,92 hojas al año (SCHMITT & WINDISCH, 2006).
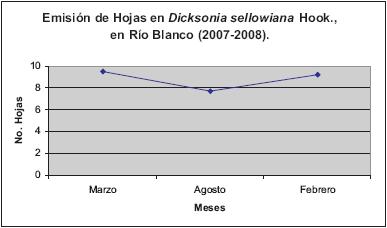

Nuestros datos indican que el tiempo de evaluación pudo ser menor que la duración de la hoja. Debe tenerse en cuenta que esta medición es parcial, puesto que en este lapso de tiempo no es posible determinar la duración de la hoja en Dicksonia. Según MEHLTRETER & PALACIOS-RÍOS (2003), el desarrollo foliar en helechos arborescentes está estimado entre uno y tres años, por lo que se recomienda aumentar el muestreo a tres años.
Los datos de fertilidad en la población evaluada no permiten registrar un patrón de esporulación. Sin embargo, únicamente las plantas que presentaron alturas mayores de 4 m se encontraron fértiles (Tabla 2). En relación con lo anterior, se han registrado en especies arbóreas pioneras de bosques mexicanos, que exhiben floración a lo largo del año, mientras que las especies de fases sucesionales tardías pueden presentarse en periodos marcados anualmente o incluso cada dos o tres años, tardando lapsos de varios años para la primera actividad reproductiva (RICKER & DALY, 1997). Esto es importante si se tiene en cuenta la esporulación de individuos pertenecientes a dicho rango de altura, la cual podría señalar que son adultos y que los otros aún no han alcanzado la etapa reproductiva.

Asimismo, es necesario incrementar el tiempo de muestreo, debido a que las hojas de estas plantas son monomórficas (MURILLO, 1988). Por esto, una vez la lámina termina de esporular, continúa en la planta con una función fotosintética y posteriormente entra en senescencia, pero, al no presentarse dehiscencia en lo pecíolos, se dificulta la observación y la determinación de su estado reproductivo.
La emisión de las hojas en D. sellowiana, se registra en módulos continuos "duplos foliares" (Figura 7), los cuales propician la acumulación de hojarasca en las hojas, como lo ha registrado CALDERÓN-SÁENZ (2000) en la Planada Nariño. Para ecosistemas altoandinos, la acumulación de materia orgánica y su lenta descomposición representan una estrategia adaptativa a las condiciones ambientales, que asegura la continua protección contra el frío, los vientos y la distribución de nutrientes en forma iónica al sistema radical (MORA-OSEJO, 1994).
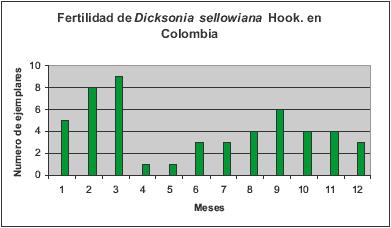
Dicksonia sellowiana responde a un patrón fenológico estacional determinado por la precipitación. Los meses de marzo y septiembre presentan los más altos registros de plantas fértiles en los ejemplares de herbario, y claramente se observan dos picos de esporulación en el año, donde el primero se localizó entre enero, febrero y marzo, incrementándose en los primeros dos meses y logrando la máxima acumulación de registros en el último mes, para posteriormente descender. El otro pico registrado se presentó en los meses de agosto, septiembre y octubre, localizándose el mayor número de registros en el segundo mes (Figura 2). Al comparar los promedios de precipitación en las regiones donde se registraron los ejemplares colectados, se determina que los picos máximos de precipitación coinciden con los picos máximos de esporulación (Figura 3).
El patrón fenológico que determina la fertilidad, la producción de semillas, la forma de los frutos, la longevidad floral y la sincronía de los frutos en las comunidades de Angiospermas, puede ser registrado y explicado por su forma de crecimiento (SARMIENTO & MONASTERIO, 1983), por los periodos de temporalidad ambiental (CROAT, 1975), por los mecanismos de aislamiento reproductivo y la presencia de polinizadores y dispersores específicos fluctuantes en el tiempo (GENTRY, 1974), y por patrones aún no explicados, que se atribuyen principalmente a errores en la toma de las variables y el desconocimiento de la filogenia de los taxones (WRIGTH & CALDERON, 2006). En helechos, aparentemente los únicos patrones registrados en fenología, son la forma de crecimiento y la variación climática, donde la precipitación influye de forma considerable (MEHLTRETER & RÍOS-PALACIOS, 2003).
Teniendo en cuenta lo anterior, en estudios realizados en la Isla de Barro Colorado (Panamá), se registró que las especies de hábitos arbustivos presentan un patrón fenológico estacional, determinado por las temporadas húmedas (CROAT, 1975). Esto puede reflejarse en la fenología de Dicksonia sellowiana, según los registros de precipitación en Colombia (Figura 8).
Cabe resaltar que un problema crítico en la estimación de la edad de helechos arborescentes, es el crecimiento del gametofito y el esporofito juvenil (BITTER & BRECKLE, 1995). Por lo que la realización de estos estudios dirigidos a la germinación de esporas en helechos arbóreos, complementaría el registro real del crecimiento de estas plantas.
La planeación del territorio y la conservación de los recursos biológicos de una área rural o silvestre, deben considerar la fauna y la flora, y el uso que las poblaciones humanas hacen de estas (TOWNSEND, 1996). En muchas ocasiones, estas comunidades dependen de dichos recursos para suplir sus necesidades básicas, y una alternativa para la conservación de los mismos está representada por su conocimiento, la generación de sentimientos de apropiación y uso y manejo sustentados en argumentos ecológicos, económicos y ambientales.
AGRADECIMIENTOSExpresamos nuestra sincera gratitud a la Universidad de Caldas, por financiar esta investigación; a la Empresa Aguas de Manizales, especialmente a Carlos Enrique Ruiz, por su apoyo logístico, y a William Cardona, por la información metereológica. A Johana Salazar, por el apoyo en campo y en diversas fases de la investigación; a Benjamín Gonzáles por su acompañamiento, y a los participantes del Semillero de Investigación en Biodiversidad y Recursos Fitogenéticos de la Universidad de Caldas, por sus comentarios y aportes en la continua construcción colectiva.
BIBLIOGRAFÍA ÁLVAREZ-MEJÍA, L.M., SANÍN, D., ALZATE-Q., N.F., CASTAÑO-R., N., MANCERA-SANTA, J.C. & GONZÁLEZ-O, G., 2007.- Plantas de la Región Centro-Sur de Caldas. Manizales: Universidad de Caldas, Cuadernos de Investigación No. 28. [ Links ] 363 pp.
ARENS N.C. & SÁNCHEZ-BARACALDO, P., 1998.- Distribution of tree fern (Cyatheaceae) across the sussecional mosaic in an Andean cloud forest, Nariño, Colombia. Amer. Fern J., 88: 60-71. [ Links ]
__________., 2000.- Variation in Tree Fern Stipe Length with Canopy Height: Tracking Preferred Habitat through Morphological Change. A. Fern J., 90 (1): 1-15. [ Links ]
ASH, J., 1987.- Demography of Cyathea hornei (Cyatheaceae), a tropical tree-fern in Fiji. Austral. J. Bot., 35: 331–342. [ Links ]
BITTNER J. & BRECKLE, S. W., 1995.- The growth rate and age of tree fern trunks in relation to habitats. Amer. Fern. J., 85 (2): 37-42. [ Links ]
CALDERÓN, E., 1997.- Lista selecta de plantas de Colombia, extintas o en peligro de extinción. Anexo 1.17. pp. 448-462. (en) CHÁVEZ-S, M.E. & ARANGO, N. (eds.), Diversidad Biológica. Tomo I. Informe Nacional sobre el estado de la biodiversidad, Colombia. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt, Ministerio del Medio Ambiente, Programa de las Naciones Unidas para el medio ambiente. [ Links ]
CONANT, D.S., 1976.- Ecogeographyc and systematic studies in American Cyatheaceae. Ph.D. dissertation, Harvard University, Cambridge, MASS. [ Links ]
CROAT, T., 1975.- Phenological behavior of habit and habitat of Barro Colorado Island, Panama Canal Zone. Biotropica, 7: 270-277. [ Links ]
CONSTANTINO, S., SANTAMARÍA-R., L.M., GÓNGORA-F., G.A., & HODSON, E., 1999.- Cultivo de tejidos vegetales como una alternativa para la preservación ex situ de Dicksonia sellowiana, un helecho arborescente amenazado: 303-308 (en) I Congreso Nacional Sobre Biodiversidad. Minambiente - Proyecto Biopacífico, Universidad del Valle, Santiago de Cali. [ Links ]
CUATRECASAS, J., 1958.- Aspectos de la vegetación natural de Colombia. Perez-Arbelaezia, 2 (8): 155-283. [ Links ]
ELEÚTERIO A.A., 2006.- Management of Tree Ferns (Cyathea spp.) for Handicraft Production in Cuetzalan, Mexico. Economic Botany, 60 (2): 182–191. [ Links ]
FONT-QUER, P., 1982.- Diccionario de Botánica. Barcelona: Labor. [ Links ]
GENTRY A.H. 1974.- Flowering phenology and divesity in tropical Bignoniceae. Biotropica, 6: 64-68. [ Links ]
HOLDRIDGE L.R., 1985.- Ecología basada en las zonas de vida. Santiago, Costarica. 199 pp. [ Links ]
HOLMGREN, P.K, KEUKEN, W. & SCHOFIELD, E.K., 1981.- Index Herbariorum. Regnum Veg. 106 (1) 7 ed. [ Links ]
KORALL, P., PRYER, K.M., METZGAR, J.S., SCHNEIDER, H. & CONANT, D.S., 2006.- Tree ferns: monophyletic groups and their relationships as revealed by four protein-coding plastid loci. Molec. Phylog. Evol., 39: 830–845. [ Links ]
KRAMER, K.U., 1976.- The american paradox in the distribution of fern taxa above the rank of species. Ann. Mo. Bot. Gdn., 77 (2): 330-333. [ Links ]
LEMA, A., 1995.- Dasometría. Algunas aproximaciones a la medición forestal. Medellín: Universidad Nacional de Colombia. 478 pp. [ Links ]
MEHLTRETER, K. & RIOS- PALACIOS, M., 2003.- Phenological studies of Acrostichum danaeifolium (Pteridaceae, Pteridophyta) at a mangrove site on the Gulf of Mexico. J. Trop. Ecol., 19: 155-162. [ Links ]
MORA-OSEJO, L.E., 1994.- Estudios ecológicos del páramo y del bosque altoandino cordillera oriental de Colombia. Santafé de Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales. [ Links ]
MORAN R.C. & RIBA, R. (vol. eds.)., 1995.- Psilotaceae a Salviniaceae. (en) DAVIDSE G., SOUSA, M. & KNAPP, S. (gen. eds.). Flora Mesoamericana. Vol. 1. Universidad Nacional Autónoma de México, Ciudad Universitaria, D. F. [ Links ]
MURILLO, M.T., 1988.- Pteridophyta-I. Flora de Colombia 9. Bogotá: Instituto de Ciencias Naturales-Museo de Historia Natural, Universidad Nacional. [ Links ]
ORTEGA, M.F.J., 1984.- Notas sobre la autoecología de Sphaeropteris senilis (Kl.) Tryon (Cyatheaceae ). En el Parque Nacional El Avila. Pittieria, 12: 31-53. [ Links ]
RICKER, M. & DALY, D.C., 1997.- Botánica económica en Bosques Tropicales. Mexico: Editorial Diana. pp. 293. [ Links ]
SANÍN, D. & DUQUE-CASTRILLÓN, C., 2006.- Estructura y composición florística de un bosque en la reserva de Río Blanco, Manizales, Caldas, Colombia. Bol. Cient. Mus. Hist. Nat. U. Caldas, 10: 44-75. [ Links ]
SANÍN, D., MANCERA-SANTA, J.C., CASTAÑO-R., N., ALZATE-Q., N.F., GONZÁLEZ-O., G. & ÁLVAREZ-MEJÍA, L.M., 2006.- Catálogo preliminar de las plantas vasculares de la Reserva Forestal Protectora "Río Blanco" (Manizales, Caldas, Colombia). Bol. Cient. Mus. Hist. Nat. U. Caldas, 10: 19-44. [ Links ]
SARMIENTO, G. & MONASTERIO, M., 1983.- Life forms and phenology. (en) BOULIER, F. (ed.). Ecosystem of the world: tropical savannas. Vol. 13: 79-108. [ Links ]
SEILER, R.L., 1981.- Leaf turnover rates and natural history of the Central America tree fern Alsophila salvinii. Amer. Fern. J., 71: 75-81. [ Links ]
__________., 1995.- Verification of estimated growth rates in the tree ferns Alsophila salvinii. Amer. Fern. J., 85: 96-97. [ Links ]
SCHMITT, J.L. & WINDISCH, P.G., 2006.- Growth Rates and Age Estimates of Alsophila setosa Kaulf. in Southern Brazil. Amer. Fern J., 96 (4): 103–111. [ Links ]
SMITH, A.R., PRYER, K.M., SCHUETTPELZ, E., KORALL, P., SCHNEIDER, H. & WOLF, P., 2006.- A classification for extant ferns. Taxon, 55 (3): 705-731. [ Links ]
TANNER, E.V.J., 1983.- Leaf demography and growth of the tree fern Cyathea pubescens in Jamaica. Bot. J. Linn. Soc., 87: 213-227. [ Links ]
TOWNSEND, W., 1996.- La utilidad del monitoreo de la cacería para la defensa del territorio. (en) CAMPOS-ROSO, C., ULLOA, A. & RUBIO TORGLER, H. (eds.). Manejo de Fauna con comunidades rurales. [ Links ]
VIS, M., 1995.- Processes and patterns of erosion in natural and disturbed andean forest ecosystems: 17-183 (en) VAN DER HAMMEN, T. & DOS SANTOS, A. (eds.), La Cordillera Central Colombiana, Transecto Parque Los Nevados. Vol. 4 (Tercera parte). Berlín, Alemania: Estudios de Ecosistemas Tropoandinos. J. Cramer. [ Links ]
WITTE, H.J.L., 1995.- Seasonal and altitudinal distribution of precipitation, temperature and humidity in the Parque Los Nevados Transect (Central Cordillera, Colombia): 279-328 (en) VAN DER HAMMEN, T. & DOS SANTOS, A. (eds.), La Cordillera Central Colombiana, Transecto Parque Los Nevados. Vol. 4. (Tercera parte). Berlín, Alemania: Estudios de Ecosistemas Tropoandinos. J. Cramer. [ Links ]
WRIGTH, S.P. & CALDERON, O., 1995.- Phylogenetic patterns among Tropical flowering phonologies. The Journal of ecology, 83 (6): 937-948. [ Links ]
Anexo 1.: Antioquia: San José de la Montaña, W. Rodríguez et al. 3555 (COL); W. Rodríguez et al. 4041 (COL); F. Giraldo et al. 1899, 1983, 2573 (COL); M. T. Murillo 5996 (COL); Nutibara, D. Sánchez et al. 452 (COL). Boyacá: Arcabuco, R. Jaramillo-Mejía et al. 3027 (COL); M.T. Murillo & M.A. Harper 2091 (COL); S. Espinal-T. et al. 1432 (COL); M. T. Murillo et al. 2091 (COL); S. Espinal-T. et al. 1573 (COL); H. Bischler 1836 (COL); F. Mejía-P. 308 (COL); Huertas y Camargo 6310 (COL); W. Hagemann 231 (COL); M.T. Murillo 1520 (COL); J. Betancur et al. 4220 (COL). Caldas: Manizales-Reserva Torre IV, D. Sanín et al. 870 (FAUC); Manizales-Reserva Río Blanco, D. Sanín et al. 2700. Cauca: Popayán, R.M. Tryon et al. 5967 (COL). Cundinamarca: W. Hagemann 303 (COL); S. Salamanca et al. 537 (COL); M.T. Murillo 467, 1087 (COL). Huila: L. Elbert et al. 8037, 9015 (COL). Magdalena: J. Cuatrecasas 25252 (COL); M.L. Grant 10765, 10992, 10975 (COL); R. Romero-Castañeda 7445 (COL). Nariño: Pasto, B.R. Ramírez et al. 4487 (COL); W. Hagemann et al. 1378, 1824 (COL); R. Bernal et al. 1422 (COL). Putumayo: J. Cuatrecasas 11673, 11829 (COL). Quindío: Salento, J. Betancur et al. 3638 (COL); P. Franco-Roselli et al. 3939 (COL). Santander: C.I. Orozco et al. 1575 (COL); G. Lozano et al. 5503 (COL); J.H. Torres et al. 524 (COL); K. P. Killip 18949 (COL); K.P. Killip et al. 19887 (COL). Tolima: M.T. Murillo 7682 (COL). Valle del Cauca: E. Calderón-Sáenz 103 (COL).

