Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.13 n.1 Manizales ene./jun. 2009
1 Universidad Javeriana. Dirección actual: University of Washington. E-mail: cgomez@u.washington.edu, carogomezposada@hotmail.com
Resumen
El presente estudio tiene como finalidad estudiar el patrón de actividad y alimentación de un grupo silvestre de micos maiceros (MC-1) del Parque Nacional Natural Tinigua (Meta, Colombia). Este grupo fue aprovisionado (es decir, que recibe diariamente una cantidad limitada de alimento –principalmente plátanos–, con fines científicos) durante más de 10 años, por tanto disfrutó de una fuente de alimento que no varía espacio-temporalmente. Por medio del método de barrido se completaron 971 horas de observación del grupo, durante seis meses en 1995-96. MC-1 invirtió 55,2% del tiempo forrajeando, seguido por movimiento (13,8%), interacciones sociales (13%) y descanso (12,1%). La actividad de espera del alimento ofrecido, denominada alerta, ocupó el 5,9% del tiempo. El grupo presentó una dieta omnívora, ingiriendo principalmente artrópodos (60,8%) y frutos (27,2%), 10,4% partes vegetales y 1,6% otros ítems. MC-1 utilizó 60 especies de plantas y consumió pequeños vertebrados, huevos y tierra de termiteros, entre otros. Tanto el patrón de actividad como el de alimentación variaron cuando el grupo fue aprovisionado, principalmente porque durante las mañanas MC-1 se dedicó a la búsqueda de artrópodos y presentó bajos porcentajes de forrajeo, contrario a lo reportado para la especie. Se concluye que el alimento ofrecido suple parcialmente los requerimientos de fruto de esta tropa, por lo cual, el grupo puede incrementar el consumo de artrópodos e invirtir menor tiempo al forrajeo y mayor a otras actividades como el descanso e interacciones sociales.
Palabras clave: Maicero cachón o cornudo, dieta, patrón de actividad, patrón de alimentación, aprovisionamiento, Parque Nacional Natural Tinigua.
AbstractThis article presents the study of the activity and feeding pattern of a wild group of brown capuchin monkeys (MC-1) in a forest sector of the Tinigua National Natural Park (Meta, Colombia). This group was provisioned daily with a limited amount of food, usually bananas, for scientific purposes for more than 10 years; therefore, enjoying a food resource that did not vary spatially or temporarily. Using scan sampling, 971 hours of troop observation were completed during six months in 1995-96. MC-1 invested 55.2% of their time foraging, followed by movement (13.8%), socially interaction (13%), and rest (12.1%). Waiting for provisioned food was named "alert," and the monkeys spent 5.9% of their time engaged in this activity. The group showed an omnivorous diet, consuming principally arthropods (60.8%) and fruits (27.2%), but also vegetables parts (10.4%) and other items (1.6%). MC-1 fed on 60 plants species, and consumed small vertebrates, eggs and termites' nests, among other items. Activity and feeding pattern varied when the group was provisioned, principally because during the mornings, MC-1 invested more time searching for arthropods, and presented low percentages of time foraging, contradicting the behavior usually reported for the species. The food offered to MC-1 partially replaces the fruit requirements for this troop, letting them increase their consumption of arthropods, decrease their foraging time, and enabling more time for other activities including rest and social interactions.
Key words: Brown or tufted capuchin, diet, activity pattern, feeding pattern, provisioning, Tinigua National Natural Park.
INTRODUCCIÓNLa forma como los individuos distribuyen el tiempo entre las actividades permite establecer cómo utilizan las horas y cómo organizan un patrón, es decir, qué hacen (actividades) y cuándo lo hacen (tiempo) (LEHNER, 1979). Los patrones de actividad de una especie son respuestas adaptativas determinadas por las características intrínsecas del animal como fisiología y comportamiento, entre otros, y por factores externos como por ejemplo la distribución y abundancia de los recursos (FRAGASZY et al., 2004).
Habituar una tropa silvestre de primates para que periódicamente visite un lugar determinado donde se le ofrece alimento, se denomina aprovisionamiento (del inglés: provisioning, A. Nishimura com. pers., 1995). Este método es empleado por algunos primatólogos, ya que permite un contacto más estrecho con la tropa y facilita la identificación de los individuos. En general, el aprovisionamiento permite realizar diferentes estudios con el grupo que está muy acostumbrado a la presencia humana y que asiste regularmente al sitio de alimentación, disminuyendo la dificultad para encontrarlo y seguirlo. El aprovisionamiento ofrece a los primates condiciones diferentes en la consecución del alimento, especialmente, porque es una fuente de recursos en un espacio limitado, generalmente muy nutritiva y que no varía en distribución y abundancia (BRENNAN & ANDERSON, 1988; FORTHMAN, 1986; LEE et al., 1986). Estas condiciones pueden afectar no solo directamente las relaciones intragrupales e indirectamente los parámetros poblacionales de las tropas, sino posiblemente también sus patrones de actividad (DUMOND, 1968; FORTHMAN, 1986; LEE et al., 1986).
En el Parque Nacional Natural Tinigua, Meta, Colombia, una tropa de Cebus apella, mico maicero, denominada MC-1, fue aprovisionado por más de 10 años consecutivos (IZAWA, 1988, 1999). Esta tropa recibió alimento en una plataforma de madera en la cual fueron observadas las relaciones jerárquicas entre los individuos, cuando estaban siendo aprovisionados. Por lo tanto, MC-1 fue base de numerosos estudios de carácter sociológico (IZAWA, 1988, 1999). El propósito del presente trabajo fue determinar el patrón de actividad y alimentación del grupo MC-1, que posee una fuente de recursos que no varía espacio-temporalmente.
Los micos maiceros C. apella (familia Cebidae) son primates omnívoros de tamaño mediano (promedio 3,7 kg), que consumen principalmente artrópodos y frutos. Esta es una especie muy activa, con elevadas tasas metabólicas (TERBORGH, 1983), por lo cual para suplir sus requerimientos debe forrajear intensivamente durante el día, siendo la alimentación la principal actividad (FREESE & OPPENHEIMER, 1981; ROBINSON, 1986; OPPENHEIMER, 1982; STEVENSON et al., 1991; TERBORGH, 1983; ZHANG, 1995). Esta especie tiene el área de distribución más extensa de todos los primates neotropicales y se encuentra en diferentes tipos de bosque, desplegando una amplia variación ecológica y conductual (DEFLER, 2003). Los maiceros son bien conocidos por sus habilidades cognitivas y capacidad de manipular objetos. Debido a estas características, esta especie es comúnmente estudiada en cautiverio con propósitos etológicos y biomédicos (FRAGASZY et al., 2004; ULYAN et al., 2006).
ÁREA DE ESTUDIOEl trabajo se desarrolló en el Parque Nacional Natural Tinigua, municipio La Macarena, Meta, Colombia (2°40' N y 74°10' W), a una altitud de 350 m, en el margen occidental del río Duda (Figura 1). El lugar de estudio consta de aproximadamente 430 ha caracterizadas por la ausencia de intervención antrópica (IZAWA & TOKUDA, 1988).
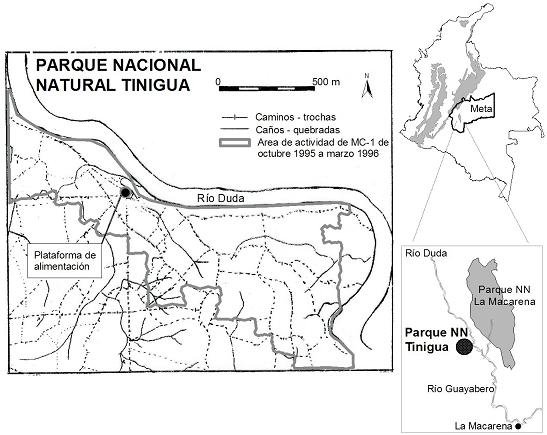
La región presenta un régimen de lluvias unimodal, con fuerte época seca de diciembre a marzo (menos de 60 mm de precipitación mensual). El promedio anual es de 2400 mm de precipitación y una temperatura promedio de 25°C (STEVENSON et al., 1991). Según la clasificación de zonas de vida de Holdridge, el área corresponde a bosque húmedo tropical estacional (KIMURA et al., 1994). El patrón de producción de frutos presenta un pico al final de la época seca, con máximos a comienzos del período de lluvias y un declive gradual hasta alcanzar los menores valores al final de la temporada lluviosa (STEVENSON et al., 1991, 2000).
La zona conocida como el corredor del Duda es uno de los centros de biodiversidad más altos del país, debido a su peculiar localización entre la Serranía de la Macarena y la Cordillera Oriental de los Andes (STEVENSON et al., 1991) (Figura 1). La gran riqueza faunística en primates se ve representada especialmente en las siete especies que habitan simpátricamente (Lagothrix lagotricha, Ateles belzebuth, Alouatta seniculus, Callicebus cupreus ornatus, Aotus cf. brumbacki, Saimiri sciureus y Cebus apella), las cuales constituyen aproximadamente el 58% de la biomasa total de mamíferos del lugar (STEVENSON et al., 1991).
MÉTODOSEl grupo MC-1 fue estudiado desde octubre de 1995 hasta marzo de 1996. Durante este período, MC-1 estuvo conformado por 18 individuos: 4 machos adultos, 2 machos sub-adultos, 5 hembras adultas, 4 machos jóvenes y 3 machos infantes. El área de actividad de esta tropa durante este período fue de 158 ha, con recorridos diarios de 1683 + o - 587 m (GÓMEZ-POSADA, 2003) (Figura 1).
Patrón de actividad. Para establecer el patrón de actividad se empleó el método de barrido (LEHNER, 1979), que permite obtener información sobre el porcentaje de tiempo invertido en diferentes actividades. Cada seis minutos se registraron las actividades de cada uno de los individuos que fue posible observar. Se definieron las siguientes actividades: Forrajeo: búsqueda e ingestión de diferentes tipos de alimentos. Movimiento: desplazamiento de un lugar a otro con una dirección definida y sin realizar forrajeo. Interacciones Sociales: incluyeron juego, cópula, acicalamiento, interacciones agonísticas intra e intergrupales. Descanso: quietud o reposo del individuo. Alerta: por la naturaleza del trabajo fue necesario definir esta categoría, que incluyó el comportamiento en la plataforma en espera de recibir o robar el alimento ofrecido.
Patrón de alimentación. Durante los barridos, fue registrado también el tipo de alimento consumido. Estos se clasificaron de la siguiente manera: artrópodos, frutos, partes vegetales y varios (el ítem varios incluyó tierra de termiteros, agua, vertebrados, huevos, entre otros).
Aprovisionamiento. Se diferenciaron dos casos:
- Cuando el grupo no fue aprovisionado: Comprendió los días en los cuales la tropa no visitó la plataforma y por lo tanto no recibió alimento ofrecido por el hombre.
- Cuando el grupo fue aprovisionado: Comprendió los días en que el grupo visitó la plataforma y consumió el alimento ofrecido allí. En ocasiones el grupo asistió a la plataforma más de una vez al día, pero solamente se les ofreció comida durante la primera visita. El alimento se les proporcionó aproximadamente una hora después de su llegada a la plataforma, generalmente trozos de 3-4 plátanos o bananos maduros, y en algunas ocasiones se complementó con galletas, por lo cual no fue posible cuantificar el alimento proporcionado.
Análisis de datos. Los datos de actividad y alimentación se organizaron por hora y día durante los seis meses, y se analizaron diferenciando los casos de aprovisionamiento. A partir de las observaciones, se obtuvieron las frecuencias relativas y los porcentajes para cada actividad y tipo de alimento. Fueron empleadas pruebas de bondad de ajuste Chi cuadrado para comparar las actividades y los tipos de alimento entre casos de aprovisionamiento. Los patrones diarios de activad y los patrones diarios de alimentación fueron comparados entre casos de aprovisionamiento, con una prueba de Fisher-Snedecor "F": ley de probabilidad del cociente. Para realizar esta prueba se tomaron los valores de X2 del segundo factor (entre horas del día) arrojados por una prueba de Friedman, con la que se analizó la variación a lo largo del día. Las correlaciones necesarias se hicieron con una prueba de Spearman rs.
RESULTADOSDurante el estudio se completaron 971 horas de observación durante 121 días de seguimiento. Del total de días en que el grupo fue seguido, un 87% visitó la mesa de alimentación y fue aprovisionado.
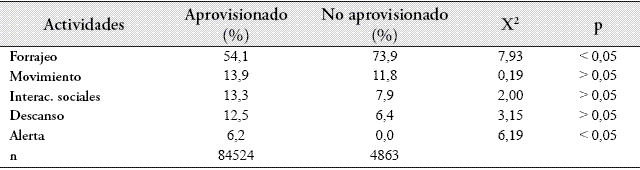
Patrón de actividad cuando el grupo fue aprovisionado. La principal actividad desarrollada por el grupo MC-1 durante el período de estudio fue el forrajeo (55,2%), seguido por el movimiento (13,8%), interacciones sociales (13%), descanso (12,1%) y alerta (5,9%). Al comparar entre casos de aprovisionamiento, forrajeo y alerta mostraron diferencias significativas, puesto que cuando el grupo fue aprovisionado, el forrajeo disminuyó y alerta incrementó (Tabla 1).
Patrón diario de actividades. La actividad de MC-1 se inició muy temprano, hacia las 05:30 h, cuando comenzó a aclarar el día y terminó hacia las 18:30 h, al oscurecer, con un promedio de actividad de 12:57 h por día. Los patrones diarios variaron entre casos de aprovisionamiento (F=2,92, g.l.=11,11, p < 0,05). Las principales diferencias se presentaron durante la mañana, especialmente para los valores de forrajeo (Figura 2).
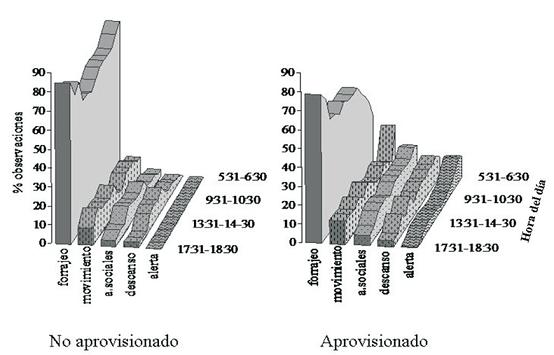
Patrón diario de actividades cuando el grupo no fue aprovisionado. En general, luego de abandonar el dormitorio, MC-1 forrajeó alrededor del mismo y se desplazó hacia una fuente cercana de alimento, la mayoría de las ocasiones a un árbol o grupo de árboles en fruto, de los cuales se alimentó una o varias veces en el día, alternándolo con excursiones en busca de artrópodos y otros tipos de alimento. Hacia el medio día, la mayoría de individuos descansaron, solos o en familias. En la tarde, el grupo realizó un forrajeo menos intensivo y hacia el final del día, empezó a desplazarse hacia el dormitorio, presentando otro pico forrajeo. Normalmente los dormitorios fueron guaduales (Guadua sp.), en donde los individuos se ubicaron por familias.
En términos de porcentajes, el forrajeo presentó los valores más altos durante todo el día (Figura 2), la mayoría superiores al 70%, con valores cercanos al 90% en las primeras horas de la mañana (05:30 – 06:30 h) y hacia el final de la tarde (17:30 – 18:30 h). Para las demás actividades se encontraron porcentajes inferiores al 20%. El descanso y las interacciones sociales mostraron una tendencia a aumentar hacia el medio día (11:30 - 14:30), cuando el forrajeo presentó los valores más bajos (menores del 70%). En el transcurso del día, el forrajeo se correlacionó de forma negativa con el descanso y con las interacciones sociales (descanso rs= 0,88, i. sociales rs= 0,91, n= 13, p<0,01).
Patrón diario de actividades cuando el grupo fue aprovisionado. En general el grupo, luego de abandonar el dormitorio, se dirigió a la plataforma de alimentación en donde recibió el alimento, descansó, se acicaló y permaneció alerta. Posteriormente, se desplazó hacia el bosque, utilizando el recurso alimenticio disponible del momento. Hacia el final de la tarde, el grupo presentó un segundo pico de forrajeo, aunque menor, desplazándose en dirección al dormitorio, usualmente ubicado cerca de la plataforma de alimentación.
En términos de porcentaje, el forrajeo fue la principal actividad desarrollada por MC-1 durante el día (Figura 2). Sin embargo, a diferencia del patrón descrito anteriormente, los porcentajes más bajos de forrajeo se presentaron entre las 05:30 y las 08:30 h, con valores menores al 50%. El resto del día, MC-1 forrajeó alrededor del 60%, con un pico cerca del 80% hacia el final de la tarde (16:30 – 18:30 h). El movimiento presentó un pico alrededor del 30% inmediatamente después de abandonar el dormitorio en la madrugada, puesto que el grupo se desplazaba rápidamente hacia la plataforma de alimentación. Los porcentajes de descanso e interacciones sociales se mantuvieron mayores al 10% a lo largo del día, disminuyendo un poco hacia el final de la tarde. Estas dos actividades no se correlacionaron con el forrajeo a lo largo del día (p>0,05). La actividad alerta presentó un pico de cerca del 10% en las primeras horas de la mañana (05:30 - 08:30), durante las visitas a la plataforma (Figura 2).
Diariamente, MC-1 permaneció cerca de 2 h en los alrededores de la plataforma de alimentación. La mitad de los días en que el grupo fue aprovisionado, regresó una segunda vez a la plataforma, generalmente hacia el medio día, permaneciendo un promedio de 1:48 minutos en los alrededores. Esta segunda visita incluyó descanso individual o por familias, sesiones de acicalamiento, además de juego por individuos jóvenes e infantes. Ocasionalmente se presentaron más visitas a la plataforma, hasta un máximo de cuatro en un día, pero estas fueron muy cortas, menores a 30 minutos. La hora de la primera visita a la plataforma de alimentación varió desde las 6:15 h, cuando no hubo oferta natural de frutos en el bosque (fin de época de lluvias), hasta las 10:00 h, cuando abundaron los frutos (inicio época de lluvias). En este último caso, el grupo primero visitó árboles en el bosque y hasta la media mañana fue en busca del alimento a la plataforma.
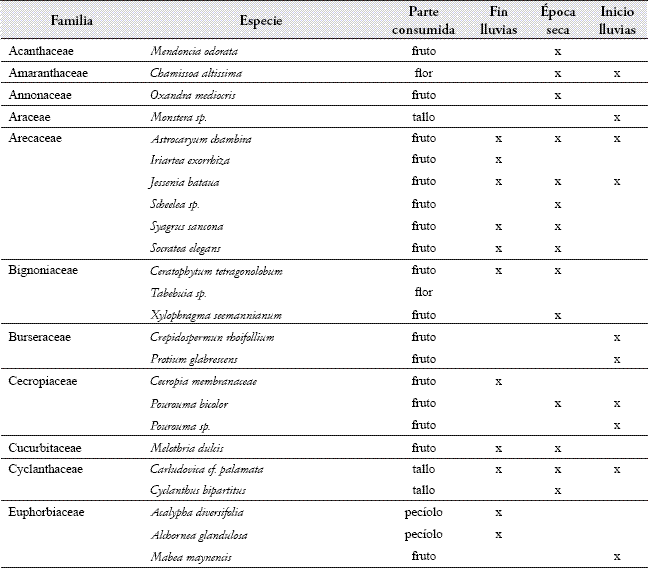
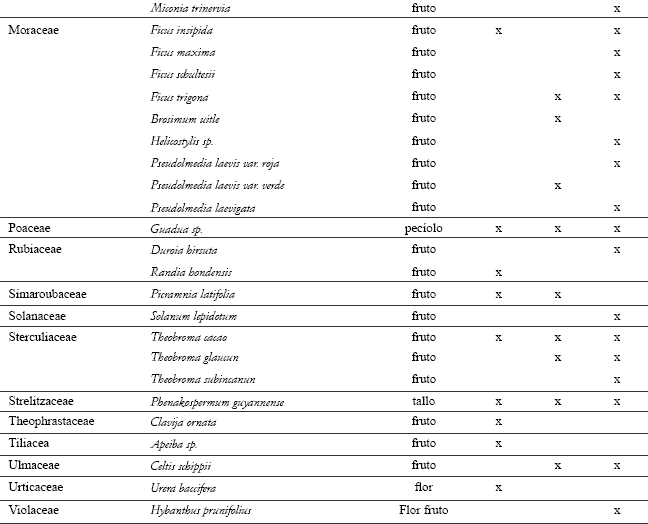
Patrón de alimentación. Durante el período de estudio, en el 60,8% de las observaciones, el grupo consumió artrópodos, 27,2% frutos, 10,4% partes vegetales y 1,6% el ítem varios. El alimento ofrecido en la plataforma correspondió a un 9% del total de las observaciones sobre alimentación.
Al comparar los tipos de alimento entre los casos de aprovisionamiento, se encontró que los consumos de frutos y de artrópodos presentaron diferencias significativas (Tabla 2). El consumo de artrópodos se incrementó y el de frutos disminuyó, cuando el grupo recibió el alimento en la plataforma (Tabla 2).

MC-1 utilizó 60 especies de frutos pertenecientes a 27 familias durante el período de estudio (ver listado de especies consumidas en el anexo). Las principales especies en la dieta fueron: Pseudolmedia laevis var. roja y var. verde, Protium glabrecens, Celtis schipii, tres especies de Inga y cuatro especies de Ficus. Durante todo el período de estudio, diferentes especies de cacao Theobroma spp. fueron recursos importantes, al igual que el tallo de platanillo (Phenakospermun guyannense) y el pecíolo de guadua (Guadua sp.), los cuales abundan en la zona. El coco del cumare Astrocaryum chambira fue de importancia en la dieta de MC-1, así como los frutos de las palmas "milpé" (Jessenia bataua) y "patona" (Socratea elegans). Se registró el consumo de diferentes huevos y polluelos de aves que fueron robados de sus nidos, ranas que viven dentro de los tallos de las guaduas (los maiceros removieron la piel de las ranas antes de consumirlas, frotándolas contra los tallos de los árboles), roedores, ardillas (Sciurus granatensis) y tierra de termiteros (incluyendo huevos y larvas de las mismas).
Patrón diario de alimentación. El patrón diario de alimentación varió entre los casos de aprovisionamiento (F=3,36, g.l.=11,11, p<0,05). Las diferencias se debieron a los bajos valores para el consumo de frutos y altos para el de artrópodos, cuando el grupo recibió el alimento en la plataforma (Figura 3). En los dos casos, el consumo del ítem varios y de partes vegetales mostraron porcentajes muy bajos a lo largo del día. Sin embargo, el consumo de partes vegetales presentó una tendencia a aumentar hacia el final de la tarde.
Patrón diario de alimentación cuando el grupo no fue aprovisionado. El mayor consumo de frutos (Figura 3) se presentó en las primeras horas de la mañana (05:31 – 08:30 h) con valores cercanos al 60% y un segundo pico al comenzar la tarde (13:30 – 15:30 h). A lo largo del día, se observó un comportamiento oscilatorio entre el consumo de artrópodos y de frutos (rs= -0,82 n=13 p<0,01), con porcentajes siempre mayores al consumo de partes vegetales y el ítem otros.
En general, luego de abandonar el dormitorio, el grupo se dirigió a uno o varios árboles en fruto de los cuales se alimentó durante las primeras horas de la mañana. El consumo de frutos se alternó con períodos de forrajeo de artrópodos, ya fuera en los mismos árboles donde estaban alimentándose, o haciendo excursiones en su búsqueda. Este comportamiento se repitió a lo largo del día. En ocasiones, MC-1 se alimentó de un solo árbol hasta que los frutos se terminaron, o visitó varios consecutivamente. El consumo del ítem varios fue esporádico y oportunista. Hacia el final de la tarde, el grupo forrajeó principalmente artrópodos y partes vegetales, incrementando el consumo de los pecíolos de guadua (Guadua sp.).
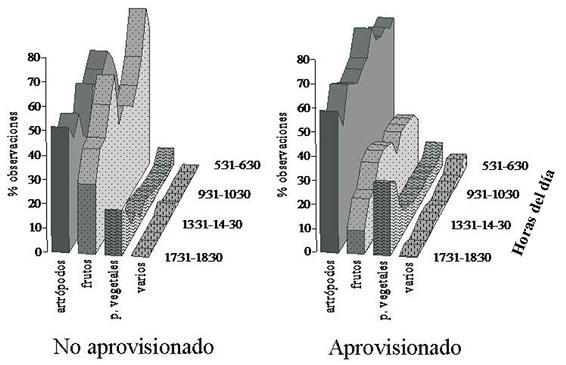
Patrón diario de alimentación cuando el grupo fue aprovisionado. Durante el transcurso del día, el porcentaje de frutos consumido fue menor que el de artrópodos y no superó en todo el día un 30% (Figura 3). El consumo de estos dos ítems no se correlacionó a lo largo del día (rs=0,13, n=13, p>0,05). Cuando MC-1 fue aprovisionado, generalmente se levantó y se desplazó primero a la plataforma y luego hacia un parche de alimentación en el bosque, donde forrajeó artrópodos y frutos. Durante los meses de inicio de las lluvias, en los cuales varios árboles de Ficus spp. e Inga spp. se encontraron en fruto, el grupo prefirió alimentarse de éstos en las primeras horas del día, antes de dirigirse a la plataforma. Hacia el final de la tarde, aumentó el consumo de partes vegetales, particularmente de pecíolos de guadua en los alrededores del dormitorio.
DISCUSIÓNPatrón de actividad. Entre las principales variables que determinan el tiempo dedicado al forrajeo están la abundancia y distribución de los recursos, la calidad nutritiva de éstos y los requerimientos energéticos del animal (CANT & TEMERIN, 1984; MILTON, 1980; ROBINSON, 1986; TERBORGH, 1983). El alimento ofrecido al grupo MC-1en la plataforma tiene un alto valor nutritivo (el plátano es en general una fruta muy rica en carbohidratos, NOVAK, 1987), está disponible para el grupo en un lugar fijo y es un recurso estable y seguro durante todo el año, es decir, no varía espacio-temporalmente. Estas características van a permitir que el grupo dedique al forrajeo un menor porcentaje de tiempo. Cuando MC-1 llega a la plataforma y espera el alimento, está "economizando" tiempo de forrajeo, el cual destina a otras actividades como descanso, alerta e interacciones sociales. Este tiempo de espera es justificado, puesto que con seguridad recibirá una cantidad apropiada de alimento, con la cual posiblemente obtendrá las mismas ganancias energéticas que si estuviera forrajeando en el bosque. Comparado con grupos silvestres no aprovisionados, el tiempo que MC-1 invirtió en el forrajeo (55%) fue un poco menor al reportado en diferentes estudios: GÓMEZ-POSADA (en imprenta) = 64%, STEVENSON et al., (2000) = 62%, TERBORGH (1983) = 66%. Una reducción similar del porcentaje de forrajeo fue encontrada por FORTHMAN (1986), al comparar dos grupos de babuinos aprovisionados en Kenya. El grupo que recibió mayor cantidad de alimento presentó bajos porcentajes de forrajeo y altos para descanso, debidos principalmente a la riqueza calórica del alimento ofrecido, la facilidad de digestión del mismo, la oferta durante todo el año y la concentración de este recurso en un sitio determinado.
El tiempo dedicado por MC-1 al descanso y las interacciones sociales fue mayor al reportado por estudios para grupos silvestres no aprovisionados del género Cebus (FREESE & OPPENHEIMER, 1981; GÓMEZ-POSADA, en imprenta; OPPENHEIMER, 1982; ROBINSON, 1986; STEVENSON et al., 1991; TERBORGH, 1983). A consecuencia del menor tiempo invertido en el forrajeo, MC-1 pudo dedicar más tiempo a otras actividades consideradas secundarias, como el descanso y las interacciones sociales. El tiempo dedicado a éstas, depende de la satisfacción de las necesidades primarias como la alimentación (TERBORGH, 1983). ULYAN et al. (2006) estudiaron el comportamiento y concentración de corticoides (como indicativo de estrés) en un grupo de C. apella en cautiverio. Los animales invirtieron mayor cantidad de tiempo en actividades sociales, y menor en el forrajeo, cuando recibieron alimento en un horario determinado y predecible. Dichos autores sugieren que rutinas de alimento predecibles pueden ser menos estresantes para los maiceros y favorecer patrones de comportamiento positivo, como las interacciones sociales y el descanso.
En general, los primates de hábitos diurnos presentan dos picos principales de alimentación: al final del día, preparándose para pasar la noche; y en la mañana, luego del prolongado descanso nocturno (CHIVERS & RAEMARKERS, 1986; TERBORGH, 1983). En las primeras horas de la mañana, deben conseguir el alimento que sea más eficiente en términos de tiempo y energía, pues es necesario satisfacer los requerimientos metabólicos y restablecer los niveles de azúcar de la sangre, después del largo período nocturno de privación de alimento (CHIVERS & RAEMARKERS, 1986). Así, diferentes estudios realizados en el genero Cebus (FREESE & OPPENHEIMER, 1981; OPPENHEIMER, 1982; ROBINSON, 1986; STEVENSON et al., 1991; TERBORGH, 1983) muestran picos de forrajeo y movimiento hacia el inicio y final de la jornada diaria; y una disminución de las actividades hacia la mitad del día (esta disminución se ha relacionado con el incremento de la temperatura ambiental, FRAGASZY et al., 2004). Cuando MC-1 no visitó la plataforma de alimentación, presentó un patrón diario de actividades similar al descrito anteriormente y que ha sido reportado para grupos no aprovisionados del género Cebus. En dichos días, el grupo presentó los más altos porcentajes de actividad durante la mañana, ya que debe forrajear para conseguir su alimento (más del 70% del tiempo). Hacia el medio día, al estar satisfechos, pudieron descansar e incrementar las interacciones sociales. Así, los porcentajes de forrajeo se complementaron de forma negativa con dichas actividades durante el día. Al final de la tarde, antes de llegar al dormitorio, el grupo presentó una fuerte actividad de alimentación, con el fin de obtener las energías necesarias para pasar la noche.
Por el contrario, cuando es aprovisionado, MC-1 durante la jornada de la mañana presentó los porcentajes más bajos de forrajeo (menores al 50%), que corresponden a las horas en las cuales visitó la plataforma. El alimento ofrecido en la plataforma suple un buen porcentaje de las necesidades alimenticias durante este período, de tal forma que el grupo necesita forrajear menos para quedar satisfecho. El resto del día los porcentajes de forrajeo se mantuvieron altos (pero no superiores al 70%), similares a los encontrados en otros estudios en grupos no aprovisionados (STEVENSON et al., 1991; TERBORGH, 1983). A diferencia de lo reportado para el género, el descanso y las interacciones sociales de MC-1 no se incrementaron hacia la mitad del día, sino que mantuvieron porcentajes cercanos al 10% a lo largo de la jornada; y no se complementaron de forma negativa con el forrajeo. Las alteraciones en el patrón de actividad de MC-1, cuando fue aprovisionado, son consecuencia de la fuente fija y nutritiva de alimento, que obtienen sin esfuerzo de forrajeo.
Patrón de alimentación. El consumo de frutos de MC-1 (27%) fue menor al reportado para esta especie en diferentes estudios de grupos silvestres no aprovisonados, en los cuales los frutos siempre ocuparon más del 40% de la dieta (FREESE & OPPENHEIMER, 1981; GÓMEZ-POSADA, en imprenta; OPPENHEIMER, 1982; ROBINSON, 1986; STEVENSON et al., 2000; ZHANG, 1995). Sin embargo, en los días que MC-1 no visitó la plataforma de alimentación, el tiempo invertido en el consumo de frutos alcanzó el 45%. El alimento ofrecido en la plataforma está supliendo una porción de los requerimientos de frutos de MC-1 y le permitió dedicar más tiempo a la búsqueda de otros tipos de alimento como artrópodos. Similar disminución en el consumo de frutos silvestres fue encontrado por NORRIS (1988), para un grupo aprovisionado de mandriles (Mandrillus sphinx) en un ambiente limitado de Gabón, África.
Esta misma situación se observó en el patrón diario de alimentación. En general, los patrones diarios de alimentación para el género Cebus muestran los mayores porcentajes de ingestión de frutos durante las primeras horas de la mañana. Estos son fácilmente digeribles y aportan los nutrientes necesarios para elevar rápidamente el azúcar en la sangre (FREESE et al., 1981; OPPENHEIMER, 1982; STEVENSON et al., 19991; TERBORGH, 1983). Cuando MC-1 fue aprovisionado, el consumo de frutos presentó bajos valores durante la mañana y en general a lo largo del día. Por el contrario, en ausencia del alimento ofrecido, el consumo de frutos fue mayor durante las primeras horas de la mañana. Es decir, cuando el grupo no fue aprovisionado, su patrón de alimentación fue similar al reportado para el género en diferentes trabajos (CHIVERS & RAEMARKERS., 1986; OPPENHEIMER, 1982; ROBINSON, 1986; STEVENSON et al., 1991; TERBORGH, 1983).
Los animales necesitan, de acuerdo con sus requerimientos metabólicos, una proporción de proteínas y energía en la dieta. Para el caso de Cebus, la proteína la obtienen en general de fuentes animales, principalmente artrópodos, y la energía, de frutos ricos en carbohidratos (JANSON & BOINSKI, 1992). Las hojas contienen proteínas y azúcares (MILTON, 1980). En el patrón diario de alimentación, cuando MC-1 no fue aprovisionado, claramente se ve la necesidad de un balance energético entre proteínas y carbohidratos en la dieta, representado en la alternancia entre el consumo de artrópodos y frutos a lo largo del día. Este patrón se alteró al recibir calorías extras sin necesidad de forrajeo, cuando fue aprovisionado; esto implicó, como ya se ha dicho, que el grupo pudiera alimentarse en mayor porcentaje de artrópodos.
A lo largo del día, los porcentajes de consumo de partes vegetales de MC-1 fueron muy bajos, y el mayor valor se presentó hacia el final de la tarde, principalmente por el consumo de hojas de guadua. Este patrón ha sido reportado para otros estudios del género Cebus, ya que las hojas luego de un largo período de fotosíntesis contienen altas cantidades de azúcar (BROWN & ZUNINO, 1990; CHIVERS & RAEMARKERS, 1986; ROBINSON, 1986; STEVENSON et al., 1991; TERBORGH, 1983; ZHANG, 1995). Adicionalmente, los Cebus no presentan adaptaciones morfológicas para la digestión de las hojas (JANSON & BOINSKI, 1992). Por tanto, las consumen principalmente al final del día, pues el reposo nocturno facilita la digestión de alimento fibroso como lo son las hojas (MILTON, 1980).
El grupo MC-1, dependiendo de la oferta natural de frutos en el bosque, cambia sus actividades y requerimientos de espacio (GÓMEZ-POSADA, 2003). Así, cuando los frutos escasearon, MC-1 disminuyó sus movimientos, área de actividad y tiempo invertido en el forrajeo, e incrementó el descanso y las actividades sociales (GÓMEZ-POSADA, 2003). En dichos períodos de escasez, el alimento ofrecido en la plataforma representó una fuente de energía importante y los animales incrementaron el número y la duración de las visitas a la plataforma (en comparación con los períodos de abundancia de frutos en el bosque), siendo la primera actividad realizada en la mañana.
Diariamente el alimento ofrecido al grupo MC-1, que obtiene sin esfuerzo de forrajeo, suple un porcentaje de sus requerimientos nutritivos. Por tanto, el grupo puede invertir menos tiempo en la búsqueda de alimentos en el bosque y puede dedicar mayor tiempo a otras actividades como descanso e interacciones sociales. A pesar de que no es mucho el alimento ofrecido, pues son solo trozos de unos pocos plátanos maduros y en ocasiones pedazos de galletas, este alimento rico en azúcares y carbohidratos suple parte de dichos requerimientos, disminuyendo el consumo de frutos en el bosque y permitiendo mayor tiempo a la búsqueda de artrópodos. Esta situación altera los patrones diarios de alimentación y actividad, reduciendo el tiempo de forrajeo, particularmente el de frutos en las primeras horas de la mañana, contrario a lo reportado para esta especie.
AGRADECIMIENTOSA los Doctores Kosei Izawa de la Universidad de Miyagi, Akisato Nishimura de la Universidad de Doshisha y Koshin Kimura de la Universidad de Nagoya, por el apoyo financiero y logístico. Al Doctor Akisato Nishimura por sus enseñanzas, apoyo y valiosas sugerencias, así como por sus amistad y por ser fuente de inspiración para estudiar primates. A la Universidad de los Andes, y especialmente al profesor Carlos Mejía, por el apoyo al proyecto. Al Doctor Jeffrey Jeorgenson, por sus comentarios sobre el trabajo escrito. A Nancy Vargas, por la asesoría y apoyo prestado para escribir el trabajo. Al Doctor Pablo Stevenson, por la identificación del material vegetal.
BIBLIOGRAFÍA BRENNAN, J. & ANDERSON, J., 1988.- Varying responses to feeding competition in a group of rhesus monkeys (Macaca mulatta). Primates, 29 (3): 353-360. [ Links ]
BROWN, A. & ZUNINO, G., 1990.- Dietary variability in Cebus apella in extreme habitats: evidence for adaptability. Folia Primatol., 54: 187-195. [ Links ]
CANT, J. & TEMERIN, A., 1984.- A conceptual approach to foraging adaptation in primates: 316-339 (en) RODMAN, P. & CANT, J. (eds.) Adaptation for foraging in nohuman primates. Columbia University Pess, New York. [ Links ]
CHIVERS, D. & RAEMAERKERS, J., 1986.- Natural and synthetic diets of malayan gibbons: 39-56 (en) ELSE, J. & LEE, P. (eds.) Primate ecology and conservation, vol. 2. Cambridge University Press, Cambridge. [ Links ]
DEFLER, T., 2003.- Primates de Colombia. Serie de Guías Tropicales 4. Conservación Internacional, Bogotá, D.C. [ Links ]
DUMOND, F., 1968.- The squirrel monkey in a seminatural environment: 88-146 (en) ROSENBLUM, L. & COOPER, R. (eds.) The squirrel monkey. Academic Press, New York. [ Links ]
FORTHMAN, Q., 1986.- Activity budgets and the consumption of human food in two troops of baboons, Papio anubis, at Gilgil, Kenya: 221-228 (en) ELSE, J. & LEE, P. (eds.) Primate ecology and conservation, vol. 2. Cambridge University Press, Cambridge. [ Links ]
FRAGASZY, D., VISALBERGHI, E. & FEDIGAN, L., 2004.- The complete capuchin, the biology of the genus Cebus. Cambridge University Press, Cambridge. [ Links ]
FREESE, C. & OPPENHEIMER, J., 1981.- The capuchin monkey, Genus Cebus: 331-390 (en) COIMBRA-FILHO, A. & MITTERMEIER, R.A. (eds.) Ecology and behaviour of neotropical primates, vol. 1. Academia Brasileira de Ciencias, Rio de Janeiro. [ Links ]
GÓMEZ-POSADA, C., 2003.- Estrategias de forrajeo de un grupo provisionado de mico maicero (Cebus apella) en el Parque Nacional Tinigua: 36-146 (en) PEREIRA, V., NASSAR, F. & SAVAGE, A. (eds.). Primatología del Nuevo Mundo: Biología, medicina, manejo y conservación. Centro de Primatología Araguatos, Bogotá [ Links ].
__________., en imprenta.- Cebus apella: variación en el patrón de actividad de acuerdo a la disponibilidad de frutos y artrópodos (en) ALARCÓN-NIETO, G. & PALACIOS, E. (eds.). Estación Biológica Mosiro Itajura-Caparú: biodiversidad en el territorio del Yaigojé-Apaporis. Conservación Internacional Colombia, Bogotá [ Links ].
IZAWA, K., 1988.- Preliminary report on social changes of black-capped capuchin (Cebus apella). Field Stud. New World Monkeys, La Macarena, Colombia, 1: 13-18. [ Links ]
__________., 1999.- Social changes within a group of wild black-capped capuchins, VI. Field Stud .Fauna Flora, La Macarena, Colombia, 13: 1-6 [ Links ]
IZAWA, K. & TOKUDA, K., 1988.- General aspects of study site. Field Stud. New World Monkeys, La Macarena, Colombia, 1: 1-3. [ Links ]
JANSON, C. & BOINSKI, S., 1992.- Morphological and behavioral adaptations for foraging in generalist primates: the case of the Cebinae. Am. J Anthropol., 88: 483-498. [ Links ]
KIMURA, K., NISHIMURA, A., IZAWA, K., & MEJÍA, C., 1994.- Annual changes of rainfall and temperature in the tropical seasonal forest at La Macarena Field Station, Colombia. Field Stud. New World Monkeys, La Macarena, Colombia, 9: 1-3. [ Links ]
LEE, P., BRENNAN, J., ELSE, J. & ALTMANN, J., 1986.- Ecology and behaviour of vervet monkeys in a tourist lodge habitat: 229-235 (en) ELSE, J. & LEE, P. (eds.) Primate ecology and conservation, vol. 2. Cambridge University Press, Cambridge. [ Links ]
LEHNER, P., 1979.- Handbook of Ethological Methods. Garland STMP Press, New York. [ Links ]
MILTON, K., 1980.- The foraging strategy of howler monkeys. Columbia University Press, New York. [ Links ]
NORRIS, J., 1988.- Diet and feeding behaviour of semi-ranging mandrills in an enclosed Gabonais forest. Primates, 29 (4): 449-464. [ Links ]
NOVAK, J., 1987.- Musa (bananas and platains): 449-488 (en) HAEMMERSCHLAG, F. & LITZ, R. (eds.) Biothechnology of perennial fruit crops. C.A.B. International, Cambridge. [ Links ]
OPPENHEIMER, J., 1982.- Cebus capucinus: Ámbito doméstico, dinámica de población y relaciones interespecíficas: 337-356 (en) LEIGH, E., STANLEY, A. & WINDSOR, D. (eds.) Ecología de un bosque tropical: ciclos estacionales y cambios a largo plazo. Smithsonian Tropical Research Institute, Panamá [ Links ].
ROBINSON, J., 1986.- Seasonal variation in use of time and space by the wedge-capped capuchin monkey, Cebus olivaceus: Implications for foraging theory. Smithsonian Institution Press, Washington. [ Links ]
STEVENSON, P., QUIÑONES, M. & AHUMADA, J., 1991-. Relación entre la abundancia de frutos y las estrategias alimenticias de 4 especies de primates en La Macarena, Colombia. Informe final presentado al Fondo para la Promoción de la Investigación y la Tecnología. Banco de la Republica, Santafé de Bogotá [ Links ].
__________., 2000.- Influence of fruit availability on ecological overlap among four neotropical primates at Tinigua National Park, Colombia. Biotropica, 32 (3): 533-544. [ Links ]
TERBORGH, J., 1983.- Five new world Primates. A study in comparative ecology. Princeton University Press, New Jersey. [ Links ]
ULYAN, M., BURROWS, A., BUZZELL, C., RAGHANTI, M.A., MARCINKIEWICZ, J.L. & PHILLIPS, K., 2006.- The effects of predictable and unpredictable feeding schedules on the behavior and physiology of captive brown capuchins (Cebus apella). Applied Animal Behaviour Science, 101: 154–160. [ Links ]
ZHANG, S., 1995.- Activity and ranging patterns in relation to fruit utilization by brown capuchins (Cebus apella) in French Guiana. Inter J Primatol., 16 (3): 489-507. [ Links ]