










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.13 no.1 Manizales Jan./june 2009
1 Departamento de Producción Animal. Decanato de Docencia, Universidad Nacional. Experimental del Táchira. Paramillo 5001, San Cristóbal, estado Táchira, Venezuela. E-mail: aorell@unet.edu.ve
Abstract
This paper aims to describe Papilio polyxenes costarum, a new subspecies, whose populations come from the Cordillera de la Costa Central of northern Venezuela. The populations of this country have traditionally been identified as belonging to the subspecies americus Kollar, 1850, which flies in Colombia and integrates to the new subspecies in the Andes of the Cordillera de Merida in Venezuela, near Colombian territories. Illustrations of adults of this new subspecies were done in order to demonstrate their new status regarding other similar subspecies. Its final-instar larva on host plants of the Apiaceae family is shown.
Key words: Papilio polyxenes costarum, neotropical, Venezuela, polymorphism, Apiaceae, immature states.
ResumenEste trabajo tiene el propósito de describir a Papilio polyxenes costarum, subespecie nueva, cuyas poblaciones provienen de la Cordillera de la Costa Central del norte de Venezuela. Las poblaciones de este país tradicionalmente se han identificado como pertenecientes a la subespecie americus Kollar, 1850, la cual vuela en Colombia y se intergrada hacia la nueva subespecie en los Andes de la Cordillera de Mérida en Venezuela, próximos a las localidades de Colombia. Ilustraron adultos de esta nueva subespecie, para demostrar su nuevo estatus con respecto a otras subespecies similares, e incluso se muestra la larva del último instar sobre una de sus plantas hospedadoras de la familia Apiaceae.
Palabras clave: Papilio polyxenes costarum, neotropical, Venezuela, polimorfismo, Apiaceae, estados inmaduros.
INTRODUCTIONUpon having the opportunity to sort and curate swallowtail butterfly specimens at the Museo del Instituto de Zoología Agrícola at the Universidad Central de Venezuela in Maracay (MIZA) an appropriate representation of the American species of the genus Papilio Linnaeus, 1758, sensu stricto (TYLER et al., 1994) was available for study. Most specimens belonged to subspecies of the black swallowtail (Papilio polyxenes Fabricius, 1775) collected from several localities of North, Central and South America. From the latter, Venezuelan material coming from the north-central coastal range, or Cordillera de la Costa, and the Andes of Mérida, was compared with neighboring countries, particularly with the tropical subspecies. It became evident that populations from the Cordillera de la Costa belong to an undescribed subspecies, despite the fact that the species has been known from this range since at least the early 20th century (RAYMOND, 1934).
Papilio polyxenes costarum, new subspecies (Figs. 1a-d)
Papilio polyxenes f. americus, RAYMOND, 1934, Bol. Soc. venez. Cienc. Nat., 2 (16): 307; Papilio polyxenes f. americus var. melasina, p. 308; Papilio machaon americus, FORSTER, 1948, Bol. Ent. venez., 7: 92; Papilio polyxenes americus, RAYMOND, 1982, Mar. Venez., pl. 32, figs. 2 & 3 (identified by F. Fernández-Y.)}


Holotype: male, red handwritten label: "Papilio polyxenes costarum Orellana/Holotipo/A. Orellana det., 2004", white handwritten and printed label: "Venezuela, Miranda/La Peñita, 1270 m / xii 1974, J. Sanz leg.", "en Eneldo". REFERVEN collection (Main- MIZA).
Paratypes: Melanic form: 8 males, 2 females Distrito Federal, El Valle, vii.1950, F. Fernández Y. leg., ex-larva en umbelifera. Cría 36; 1 male Distrito Federal, El Manantial, El Junquito, 1200 m, vi.1982, S. Vera leg., [pupal exuvium preserved]; 1 female Distrito Federal, Bajo Seco, vii. 1982, O. Aponte leg., En anicillo n° 73 1982, [pupal exuvium preserved]; 1 male Miranda, San Antonio de los Altos, 1350 m, i. 1956, F. Fernández Y. leg., ex-larva (8.i.1956) en anís silvestre, crisálida (12.i.1956), imago (22.i.1956), [pupal exuvium preserved]; 1 female Miranda, Mariches, carretera de Petare, viii.1953, C. J. Rosales leg; 1 female Yaracuy, Nirgua, vi.1962; 1 female Carabobo, Cumbre Aguirre, xi.1951, F. Fernández Y. leg; 1 female Carabobo, Palmichal, cr. Canoabo, viii. 1984, F. Fernández Y. y C. J. Rosales leg., ex larva en apio, [pupal exuvium preserved]; Yellow form: 1 male Same as holotype. 3 males Distrito Federal, El Manantial, El Junquito, 1200 m, vi. 1982, S. Vera leg., [pupal exuvium preserved]; 1 male Distrito Federal, est. exp. Bajo Seco, El Junquito, 2000 m, iii.1978, C. J. Rosales leg; 2 male Miranda, San Antonio de los Altos, 1350 m, i.1956, F. Fernández Y. leg., ex-larva (8-i.1956) en anís silvestre, crisálida (12.i.1956), imago (22.i.1956), [pupal exuvium preserved]; 1 male y 1 female Miranda, Núcleo El Laurel, 1200/1300 m, ix.1972, A. Montagne y J. Sanz leg., ex larva en apio; 1 male idem. x.1972; 4 males Distrito Federal, El Manantial, km 12 El Junquito, 1200-1300 m, vi.1982, A. Vera leg.; 1 female Distrito Federal, km 14 carretera El Junquito, vi. 1981, D. Vargas leg; 1 female Distrito Federal, Bajo Seco, vi. 1982, D. Grance leg., en anís de monte n° 53 1982, [larval and pupal exuviae y preserved]; 1 female Aragua, Col. Tovar, 2000 m, v. 1978, C. Michelangelli, ex-pupa en rama de clavel; 1 female Miranda, Bajo Seco, 1650 m, i. 1972, E. Ollarves, ex-larva en apio; 1 female Carabobo, Palmichal, cr. Canoabo, viii. 1984, F. Fernández Y. y C. J. Rosales, ex larva en apio, pupa verde amarillo, [pupal exuvium preserved] (all in MIZA); 1 female Miranda, Sn. Antonio de los Altos, 1300 m, i.1972, A. Montagne, ex-larva en apio (ex-MIZA. Bollino collection, Lecce-Italy); 1 female Distrito Federal, Caracas, Las Acacias, 900m, 2. xii. 1966 P. Bermúdez leg., ex-larva (Romero family collection, Maracay-Venezuela); 1 male Miranda, Sabaneta, near Turgua, S.E. of Caracas, 1000 m, R. DeLaFuente leg., coll. Neild 9.ii. 1980 (Neild collection, London-UK).
Diagnosis: Yellow hindwing bar is far less wide than in P. polyxenes americus Kollar, 1850, being similar to P. polyxenes stabilis Rothschild & Jordan, 1906 and P. polyxenes sadalus Lucas, 1852 (see Fig. 2). The bar occupies the distal quarter of discal cell and never touches the base of CuA2. On the ventral surface of the HW, the basal quarter is less yellow, being clothed in dark gray scaling. Both surfaces of the forewing are similar to americus, blotches slightly smaller. A melanic form is also known, but it is indistinguishable from that of P. p. americus described as f. melasina Rothschild & Jordan, 1906.

Description: Yellow form. Body and wings black with pale yellow markings. Thoracic and cephalic appendages black. Head with two yellow spots behind the eyes and antennae. Genae yellow. Eyes dark walnut brown. Patagia and tegulae with a yellow dot/streak each (respectively). Abdomen with two dorsolateral dots on the sides of each segment. Dorsal surface: Forewing with two rows of yellow blotches. That in the median area with larger blotches, forming a straight bar interrupted at veins, starting at the distal quarter of the basal margin to the proximities of the apex. The blotch below R3 is shifted towards the discal cell. Blotch on space R4 is divided into a large distal one and a smaller proximal. The external row lacks a blotch on 1A. The end of discal cell has a yellow streak. Terminal edge alternates yellow streaks between veins. Hindwing with a median yellow bar, barely twice as wide as the thorax on its broadest portion, extending across the center of the wing from the inner edge to the anal edge passing through the distal end of the discal cell. The bar widens at its middle portion and abruptly acuminates at the tornal area. There, the edge is bordered by yellow from the end of the bar towards the base of the wing. Veins at end of cell that enter the bar are broadly black. A row of yellow crescents and an orange, distally graded to yellow, anal black-pupilled eyespot lies near the terminal border. Between this row and the central bar is a dusting of blue scales, much reduced towards the apex where it is totally absent. Depth of teethed terminal edge streaked with yellow. No-spatulate straight short tail at end of M3. Ventral surface: Background dark gray, with vestigial veins in cell (M) and basal area (CuP) yellow. Basal area slightly dusted in yellow. Same elements as on dorsal surface, only that yellow blotches are slightly larger, thus enabling the dorsal blotches see-through due to excess black. Yellow blotches with an additional smaller orange dot shifted distally on the median bar and almost completely on submarginal row. Blue scaling more produced. Distal edge of median bar lined in black. Anal and inner edge bordered i yellow.
Variations: Females differs from the male holotype in the rounder wing shape, larger size, and to some extant, in having few more submarginal blue scales; otherwise similar. A melanic morph is also known to occur on both sexes, not distinguishable from the dark morph of P. polyxenes americus (=melasina Rothschild & Jordan, 1906). Altogether, the subspecies is fairly constant both on the yellow and dark morphs. Cordillera de Mérida populations occasionally produces individuals with broader yellow bands, having the proximal edge irregularly dusted in yellow.
Etymology: This butterfly subspecies is dedicated to my friends Mauro Costa and his recently deceased wife Clara, both having engaged in exciting studies of Venezuelan butterflies, encouraging many of us to keep along the trail. This name is appropiate considering the range of distribution, and is composed following ICZN 1999.
Biology: Larvae have been raised from several umbelliferous hostplants (Apiaceae), mainly common dill (eneldo, apio Anethum graveolens L.) and fennel (hinojo, Foeniculum vulgare L.). RAYMOND (1934) describes the larvae and notes the apparent emergence of greater number of melanic individuals during drier years. Figure 3 shows a final (fifth) instar larvae. Apparently, melanic individuals occur at lower elevation on a much higher frequency than at higher elevations, but this needs to be tested. It seems also that several populations have decreased in size, particularly at lower elevations and certain valleys (ORELLANA & ERAZO, 2002) which leads to propose global warming as causing a shift to higher elevations and subsequent extinction at lower sites. The species has not been seen for several years near Canoabo in Carabobo state around 600 m.a.s.l. where it was common (N. Flauger, com pers).

DISCUSSION
In a restricted sense, the genus Papilio clusters those species in ROTHSCHILD & JORDAN'S (1906) group V Machaon (TYLER et al., 1994). Under this consideration, the immense size of the genus is greatly reduced, as it has historically embraced taxa today referred to the genera Pterourus Scopoli, 1777; Heraclides Hubner, [1819]; Sinoprinceps Hancock, 1983; Princeps Hubner, [1807]; Druryia Aurivillius, 1881; Achillides Hubner, [1819]; and Menelaides Hubner, [1819] (HÄUSER, 2003). Moreover, several authors (including ROTHSCHILD & JORDAN, 1906) have placed species of the tribes Troidini and Leptocircini (=Graphiini, =Luehdorfiini) in Papilio, hence, becoming equivalent to the whole subfamily Papilioninae. Others recommend that it is not advisable to split Papilio (EHRLICH & MURPHY, 1982; MILLER, 1987), even though they do recognize a few subgenera (HÄUSER, 2003).
As classified in this work, the greatest diversity of Papilio occurs in the boreal hemisphere, comprising at least four species if vicariant theory is taken in consideration. Many accepted taxa regularly taken as species must then be considered but names of subspecific nature. The composition would be as follows: P. alexanor Esper, 1799 (Europe and Asia), P. indra Reakirt, 1866 (western North America), P. machaon Linnaeus, 1758 (circumboreal), and P. polyxenes Fabricius, 1775 (North America south to northern South America). Despite this scheme, many insist that at least P. hospiton Guenée, 1839 (Corsica and Sardinia) and P. saharae Oberthur, 1879 (northern Africa) should be treated as full species very closely related to P. machaon, based on breeding experiments and other evidence. What becomes clear is that all species are highly polymorphic, with more than 60 subspecies recognized as valid for the entire genus. In the words of CLARKE & LARSEN (1986) it seems that the complex is still evolving and the relationships within the various forms are not adequately expressed in terms of the traditional specific or subspecific categories. This last may explain why there is no clear consensus on the number of species actually occurring particularly in the genus and in Papilionidae in general (SPERLING, 1987; HAGEN et al., 1992; PITTAWAY et al., 1994).
Of the above species, only Papilio polyxenes enters into equatorial latitudes, prevailing there in cool montane environments. A large array of combination and synonyms have been collected for this species (see ROTHSCHILD & JORDAN, 1906; D'ALMEIDA, 1965; TYLER et al., 1994) owing to its great variability and polymorphism (Fig. 2) as well as to a lack of any unifying criteria and stable species concept until only a couple of decades ago. Southern populations of subspecies asterius Stoll, 1782 in Mexico seems to be of greatest variability.
The following is a brief account on the nomenclatural subject for populations under our concearn. According to GODMAN & SALVIN (1893), polyxenes and americus KOLLAR (1850) were considered to be separate species during the latter half of the 19th century the former ranging from North America south to Nicaragua and Costa Rica, whereas the latter, from this last country to Ecuador, Colombia and Venezuela. ROTHSCHILD & JORDAN (1906) assumed both are conspecific and questioned the capture of americus from "Angostura, Orinoco" undoubtedly referring to the Ciudad Bolívar of today (Fig. 4), on the south shore of the lower Orinoco river in Venezuelan Guayana. This error was continuosly extended later on by the authors of the "Biologia Centrali Americana" based on an understandable misinterpretation of the original description. KOLLAR (1850) mistook this Venezuelan locality for the true type locality of Angostura, in the Colombian department of Antioquia (LAMAS, 1994). ROTHSCHILD & JORDAN (1906) also described subspecies stabilis from Central America, recognized asterius as a subspecies of P. polyxenes and introduced sadalus Lucas, 1852 under the synonymy of P. p. americus. However, both P. zelicaon Lucas, 1852 and P. nitra Edwards, 1883, remained on the specific status for the reviewers of the "American Papilios". D'ALMEIDA (1965) used P. ajax Linnaeus, 1758 to cluster americus, brevicauda Saunders, 1868 and polyxenes and left asterius as a junior synonym. It is warranted that ajax was earlier "suppressed" by ROTHSCHILD & JORDAN (1906), but it was formally done so by the commission (ICZN: Papilio ajax Linnaeus, 1758 rejected, Opinion 286). The most recent synonymic checklist for P. polyxenes is given by TYLER et al. (1994) based upon a myriad of experimental works (REMINGTON, 1959; AE, 1960; EMMEL & EMMEL, 1964; FISHER, 1977; EMMEL & SHIELDS, 1978; FERRIS & EMMEL, 1982; SPERLING, 1987, 1993) and followed by LAMAS (2004). The last addition is the recent description of a distinctive north Peruvian subspecies (BOLLINO & VITALE, 2002).
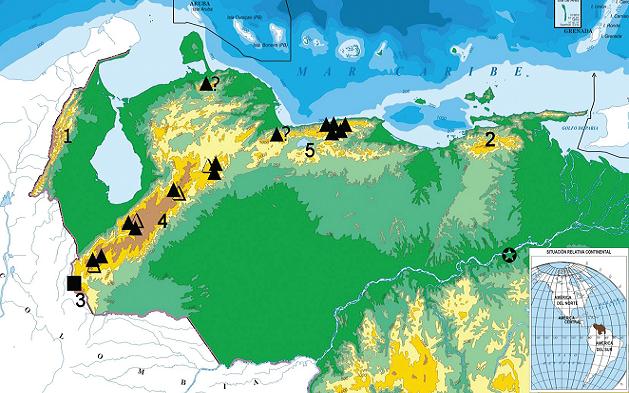
It is clearly evident that P. polyxenens americus is a subspecies described from Colombian material as shown by its original description (KOLLAR, 1850), subsequent literature (ROTHSCHILD & JORDAN, 1906), the material available for this study from the critical analysis of LAMAS (1994) and an examination of the lectotype specimen (Fig. 1-e). Concerning Venezuela, as it is mentioned in ROTHSCHILD & JORDAN (1906), the material from this country was taken in the vicinities of Mérida in the Andes, but the authors do not mention any locality from the central coast range, suggesting that few or none were then hosted in European collections. In this study, it became evident that some specimens from Mérida range show a close resemblance to americus and since authors frequently took other similar subspecies (sadalus and stabilis) as synonymous, a less broad banded polyxenes from Venezuela must also had been americus for these authors. RAYMOND (1934) is then the first to mention the species from Caracas and its surrounding (having identified it as subespecies americus) and notices also the melanic form melasina. Two specimens appeared in the facsimile edition of the watercolor paintings portrayed posthumously in his book (RAYMOND, 1982). Later, FORSTER (1948) lists the species as being collected from Caracas cited under the incorrect nomination P. machaon americus. Since then, all references have used the trinomial P. polyxenes americus (RAYMOND, 1982; ÁLVAREZ & ÁLVAREZ, 1984; ORELLANA & ERAZO, 2002; ORELLANA & GONZÁLEZ, 2003). It seems then that no one had ever before verified the status of Venezuelan black swallowtail, thus it seemed easier to assume that the name americus was granted for all Venezuelan populations.
CONCLUSIONSWith the large series available some conclusions can be drawn relating to the distribution of Venezuelan populations of the black swallowtail, P. polyxenes (please consult Fig. 4). 1) Populations from the north central coastal range (Cordillera de la Costa) with individuals showing great constancy in the phenotype, also appearing in two genetically balanced morphs, one melanic, the other yellow merit an infraespecific nomination separate from subespecies P. polyxenes americus, this being the purpose of this paper. 2) Andean populations in the Cordillera de Mérida, also dimorphic belong to the same subspecies, but many specimens throughout showing some gradation to P. polyxenes americus as evidenced by the heavy yellow dusting and widening of the yellow middle bar on the basal area of the hindwing on both surfaces (Fig. 1d). This suggests some genetic introgression from P. p. americus. 3) Populations from the El Tamá range, a continuation of the Colombian Eastern Cordillera: despite the few individuals available (n=8), all yellow individuals show an even wider middle yellow bar and none like P. polyxenes costarum, these being assumed to belong nearer to P. polyxenes americus, though this may be the case. 4) The discontinuity of the mountains at geographical depressions may not represent genuine genetic barriers, although this has nt been tested (Táchira depression, separating the El Tamá and Mérida ranges; and the Turbio-Yaracuy depression separating the Mérida range from the Cordillera de la Costa). 5) A single specimen from the Falcón range (south of Paraguaná peninsula) is melanic, and thus cannot be classified as either andigena or americus. No specimens are known from the Perijá range. The existence of the black swallowtail in the north-eastern coastal range (Turimiquire massif) and extreme north Andes at norhtwestern Venezuela in Perijá cannot be ruled out.
ACKNOWLEDGEMENTSThe encouragement and assessment of José Clavijo and Jürg Demarmels are greatly acknowledged. Quintín Arias kindly provided his photographic expertise and time for specimens depicted in figure 1. Gerardo Lamas contributed with valuable corrections and comments to a Spanish version as well as with a photograph of the lectotype of Papilio americus (Fig. 1-e). Andrew Neild made important corrections to the English manuscript and suggestions. Thanks are given to Maurizio Bollino for sharing ideas and information. To Francisco Romero for access to his family's collection and to Eustaquio Arnal for providing an excellent photograph of the larva (Fig. 3). This work is part of the results of the short survey and special issue Catalogación y Base de Datos de la colección de Papilionidae (Insecta: Lepidoptera) del Museo del Instituto de Zoología Agrícola "Francisco Fernández Yépez"' credited to graduate studies in Entomology at the Facultad de Agronomía of the Universidad Central de Venezuela.
BIBLIOGRAPHY AE, S.A., 1960.- A study of hybrids between Papilio xuthus and the P. polyxenes/machaon group. Journal of the Lepidopterists' Society, 14, 15-18. [ Links ]
ÁLVAREZ-S, J.R. & ÁLVAREZ-C, J.R., 1984.- Mariposas Diurnas de Venezuela. Introducción a su conocimiento. Caracas, Editorial Arte. 200 pp. [ Links ]
BOLLINO, M. & VITALE, F., 2002.- Una nuova sottospecie di Papilio polyxenes Fabricius, 1775 (Lepidoptera: Papilionidae) del Perú (Sud America). Lambillionea, 52, 152-155. [ Links ]
CLARKE, C. & LARSEN, T.B., 1986.- Speciation problems in the Papilio machaon group of butterflies (Lepidoptera: Papilionidae). Systematic Entomology, 11, 175-181. [ Links ]
D'ALMEIDA, R.F., 1965.- Catálogo dos Papilionidae americanos. São Paulo, Sociedade Brasileira de Entomologia. 366 pp. [ Links ]
EHRLICH, P. & MURPHY, D.D., 1982.- Butterflies and biospecies. Journal of Research on the Lepidoptera, 21, 219-225. [ Links ]
EMMEL, J.F. & EMMEL, T.C., 1964.- Genetic relationships of Papilio indra and Papilio polyxenes. Journal of Research on the Lepidoptera, 3, 157-158. [ Links ]
EMMEL, J.F. & SHIELDS, O. 1978.- Larval foodplant records for Papilio zelicaon in the western United States and further evidence of the conspecificity of P. zelicaon and P. gothica. Journal of Research on the Lepidoptera, 17, 56-67. [ Links ]
FERRIS, C.D. & EMMEL J.F., 1982-. Discussion of Papilio coloro W.G. Wright (=Papilio rudkini F. & R. Chermock) and Papilio polyxenes Fabricius (Papilionidae). Bulletin of the Allyn Museum, 76, 1-13. [ Links ]
FISHER, M.S., 1977.- The taxonomy and identity of Papilio nitra W.H. Edwards in Colorado (Papilionidae). Bulletin of the Allyn Museum, 47, 1-7. [ Links ]
FORSTER, W., 1948.- Liste der von Pater Cornelius Vögl in Maracay und Caracas gesammelten Schemetterlinge. I. Rhopalocera. Boletín de entomología Venezolana, 7, 91-120. [ Links ]
GODMAN, F.D. & SALVIN, O., 1893.- Biologia Centrali Americana. London, Dulau & Co., Bernard Quaritch. vol 2, pp. 241-256. [ Links ]
HAGEN, R.H., LEDERHOUSE R.C., BOSSERT J.L. & SCRIBER J.M., 1992.- Papilio canadensis and Papilio glaucus are distinct species. Journal of the Lepidopterists' Society, 45, 255-258. [ Links ]
HÄUSER, C.L., 2003.- Papilionidae - revised GloBIS/GART species checklist (2nd draft). Prepared by Christoph L. Häuser, in cooperation with Rienk de Jong, Gerardo Lamas, Robert K. Robbins, Campbell Smith & Richard I. Vane-Wright. <http://www.insects-online.de/gartfron.htm> Last changes: 10th July 2003. [ Links ]
KOLLAR, V., 1850.- Beiträge zur Insekten-Fauna von Neu-Granada und Venezuela. Denkschr K Akademie Wiss Wien, 1, 351-364. [ Links ]
LAMAS, G., 1994.- Las mariposas sudamericanas descritas por Vincenz Kollar (Lepidoptera). Revista peruana de Entomología, 37, 55-58. [ Links ]
__________., 2004.- Papilionidae. 96. pp 87-98. In: Lamas G (Ed.) Checklist: Part 4A. Hesperioidea – Papilionoidea. In: HEPPNER, J. (ed.) Atlas of Neotropical Lepidoptera. Gainesville. Association for Tropical Lepidoptera/Scientific Publishers. [ Links ]
MILLER, J.S., 1987.- Phylogenetic studies in the Papilioninae (Lepidoptera: Papilionidae). Bulletin of the American Museum of Natural History, 186, 365-512. [ Links ]
ORELLANA, A.M. & ERAZO, M.C., 2002.- Posible declinación poblacional de la mariposa Papilio polyxenes americus Kollar (Lepidoptera: Papilionidae) en el valle intra-andino de Mérida, Venezuela. Entomotropica, 17, 189-190. [ Links ]
ORELLANA, A.M. & GONZÁLEZ, J.M., 2003.- Adiciones, rectificaciones y actualizaciones a "Mariposas de Venezuela" por Théophile Raymond. III. Castniidae y Papilionidae (Lepidoptera). Entomotropica, 18, 73-76. [ Links ]
PITTAWAY, A.R., LARSEN, T.B., CLARKE, C.A., SMITH, C.R., CRNJAR, R. & CLARKE, F.M.M., 1994.- Papilio saharae Oberthür, 1879 is specifically disitinct from Papilio machaon (L., 1758) (Lepidoptera: Papilionidae). Entomological Gazette, 45, 223-249. [ Links ]
RAYMOND, T., 1934.- Lepidópteros de Venezuela. Familia Papilionidae. Boletín de la Sociedad venezolana de Ciencias Naturales, 2(16/17), 284-352. [ Links ]
__________., 1982.- Mariposas de Venezuela. Caracas, Ediciones Corpoven. 227 pp. [ Links ]
REMINGTON, C.L., 1959.- Wide experimental crosses between Papilio xuthus and other species. Journal of the Lepidopterists' Society, 13, 151-164. [ Links ]
ROTHSCHILD, W.L. & JORDAN, K., 1906.- A revision of the american Papilios. Novitates Zoologicae, 13, 412-752. [ Links ]
SPERLING, F.A.H. 1987.- Evolution of the Papilio machaon species group in western Canada (Lepidoptera: Papilionidae). Quastiones entomologicae, 23, 198-315. [ Links ]
__________., 1993.- Mitochondrial DNA phylogeny of the Papilio machaonspecies group (Lepidoptera: Papilionidae). In: G.E. BALL & H.V. BANKS (eds.) Systematics in support of Entomology. Memoirs of the Entomological Society of Canada, 165, 233-242. [ Links ]
TYLER, H.A., BROWN, JR K.S. & WILSON, K.H., 1994.- Swallowtail Butterflies of the Americas. Scientific Publishers, Gainesville, Florida. 376 pp. [ Links ]

