










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.13 n.1 Manizales ene./jun. 2009
1 Estudiante de Biología, Universidad de Caldas. Ingeniero Civil, Universidad Nacional de Colombia. E-mail: adrian.salazar@ucaldas.edu.co, adrianzuns@hotmail.com.
Resumen
Este estudio tiene como objetivo examinar la relación entre el nivel de agresividad intraespecífica de la hormiga poligínica Crematogaster longispina, y la distancia que separa los árboles de yarumo blanco (Cecropia telealba) en los que reside cada colonia. Además, se indaga si existen diferencias en el grado de hostilidad que presenta C. longispina frente a Myrmelachista sp. Con este propósito, se observaron encuentros entre forrajeras que fueron calificados según su nivel de agresividad. Se encontraron diferencias en el grado de agresión intraespecífico, pero no en el interespecífico. No se halló correlación entre el nivel de agresión intraespecífico y la distancia entre árboles. Se sugiere la medición de otras variables que puedan explicar las diferencias en los niveles de agresión intraespecífica. También, se destaca que la variación en la hostilidad intraespecífica se puede entender mejor si se cuenta con estudios de estructura genética de las colonias.
Palabras clave: Hormigas, Crematogaster longispina, Myrmelachista, Hymenóptera, Formicidae, poliginia, comportamiento, agresividad, reconocimiento de miembros de colonia, discriminación de parientes.
AbstractThis study examines the relation between the intraspecific aggressiveness level of the polygynous ant Crematogaster longispina, and the distance separating white yarumo trees (Cecropia telealba), this species' habitat. In addition, it investigates whether the degree of interspecific hostility between C. longispina and Myrmelachista sp. has differences. The aggressiveness level was measured through the observation of encounters between worker ants, graded according to an aggressiveness scale. There were differences in the intraspecific aggression degree, but not in the interspecific one. No correlation was found between the intraspecific aggression level and the distance between trees. The measurement of other variables that could better explain the differences in intraspecific aggression levels is suggested. The variation in the intraspecific hostility can be better understood if studies on the genetic structure of colonies are conducted.
Key words: Ants, Crematogaster longispina, Myrmelachista, Hymenoptera, Formicidae, polygyny, behavior, aggressiveness, colony member recognition, kin discrimination.
INTRODUCCIÓNA pesar de que las hormigas presentan conductas altruistas, también manifiestan esquemas agresivos tendientes a la protección de su territorio (HÖLLDOBLER & WILSON, 1990). Los comportamientos territoriales incluyen los costos de tiempo y energía invertidos en marcar una zona y defenderla, así como los riesgos de luchar por ella; pero también generan beneficios dentro de los cuales se encuentran los derechos de exclusividad sobre el alimento y un mayor éxito reproductivo (DAWKINS, 1976/1993; TINBERGEN, 1951/1989). Sin embargo, sólo se empezaron a tener indicios de la manera como las hormigas escogían a los destinatarios de sus ataques, desde que la conducta agresiva en insectos sociales empezó a explorarse en el contexto del reconocimiento de los miembros de nido (nestmate recognition) (KRAVITZ & HUBER, 2003). La capacidad para distinguir a los miembros de la colonia frente a individuos foráneos se hace preponderante para la vida social de las hormigas, ya que constituye una de las formas para favorecer exclusivamente a los parientes con las ventajas adquiridas por toda la comunidad (HÖLLDOBLER & WILSON, 1990).
Los investigadores han encontrado que la respuesta de las hormigas hacia individuos de otra colonia de la misma especie, se ubica en un punto dentro de un amplio gradiente de conductas. En un extremo, el intruso será aceptado pero con un menor ofrecimiento de comida hasta que haya adquirido el olor de la colonia, y en el otro, los residentes atacarán al invasor con violencia, aferrando las mandíbulas a su cuerpo o a sus apéndices, mientras lo aguijonean o lo rocían con ácido fórmico, citronelal u otra sustancia tóxica (HÖLLDOBLER & WILSON, 1990).
Al menos dos factores pueden afectar el reconocimiento de los miembros de la colonia y explicar la hostilidad intraespecífica. En primer lugar, se encuentran las diferencias genéticas entre colonias que darán como resultado disparidades en las feromonas producidas para reflejar las necesidades colectivas, y en segundo lugar están las diferencias en los olores provenientes del ambiente que contribuyen a la conformación del olor específico de la colonia (HÖLLDOBLER & WILSON, 1990; SATOH & HIROTA, 2005). De esta manera, se podría esperar que entre mayor separación haya entre los nidos, mayor será la diferencia entre sus tipos de olor y, en consecuencia, serán mayores las respuestas agresivas entre colonias (HEINZE et al., 1996; SATOH & HIROTA, 2005).
Respecto a las interacciones entre distintas especies de hormigas, se ha encontrado que los miembros de una colonia pueden expresar desde un comportamiento neutral hasta un ataque en masa hacia los adversarios potenciales (CARLIN & JOHNSTON, 1984; FEENER, 1986, 1987; HÖLLDOBLER, 1979, 1983). Este fenómeno se conoce como discriminación del enemigo (enemy specification) y se piensa que es un proceso clave en la organización de comunidades de hormigas, ya que provee las condiciones para que una especie débil pueda vivir junto a una dominante, y dirige los efectos de la exclusión de una especie del territorio de otra, acentuando los patrones del mosaico de hormigas (HÖLLDOBLER & WILSON, 1990).
En esta investigación se buscan relaciones entre el grado de agresividad intraespecífica de la especie poligínica Crematogaster longispina y la distancia que separa los árboles en los que se encuentran sus colonias. Adicionalmente, se examina si existen diferencias en el grado de hostilidad que presenta C. longispina hacia Myrmelachista sp.
MATERIALES Y MÉTODOS Área de Estudio
El trabajo de campo se llevó a cabo en el municipio de Pereira (Risaralda, Colombia), en la finca "El Cedral" (4° 45' N - 75° 38' W), c. 1800 m de altitud, perteneciente a la cuenca media del río Barbas en los Andes centrales colombianos. El área de estudio se caracteriza por presentar fragmentos de bosques secundarios inmersos en una matriz de plantaciones forestales comerciales, compuestas principalmente por cultivos exóticos de eucalipto (Eucalyptus grandis), y de pino (Pinus caribaea y Pinus patula) en menor proporción.
Recolección de muestras
Forrajeras de C. longispina y de Myrmelachista sp. fueron colectadas vivas de seis árboles de yarumo blanco (Cecropia telealba), ubicados en dos diferentes tipos de fragmentos boscosos (Figura 1). Para la colecta se usó el método de captura directa con aspirador, pinzas y pinceles (SARMIENTO, 2003).
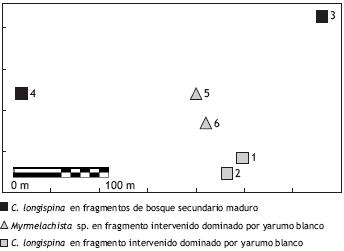
Pruebas de agresividad
Para cada enfrentamiento se ingresaban un par de hormigas dentro de un contenedor transparente de 20x5x3 mm. Para el estudio de agresividad intraespecífica se opusieron entre sí forrajeras de C. longispina provenientes de diferentes árboles. Para el estudio de agresividad interespecífica se enfrentaron forrajeras de C. longispina contra forrajeras de Myrmelachista sp.
Después de ingresar cada pareja en el contenedor, sus interacciones eran observadas durante cinco minutos a través de un estereoscopio (16X), tiempo durante el cual se calificaba el nivel de agresividad a partir de una escala modificada de HÖLLDOBLER & WILSON (1990) (Tabla 1). Para cada combinación se realizaron por lo menos siete encuentros con hormigas diferentes (es decir, renovándolas en cada turno), hasta alcanzar un total de noventa y ocho mediciones. Adicionalmente, se efectuaron pruebas de agresión entre forrajeras de un mismo árbol como un control (cinco parejas por árbol, para un total de treinta ensayos). Todas las pruebas fueron realizadas dentro de la semana de recolección (25-31 de julio de 2007).

Análisis estadístico
Para examinar correlación entre hostilidad y distancia entre árboles, se realizaron pruebas de rangos ordenados de Spearman y de Kendall (SIEGEL & CASTELLAN, 1995; SOKAL & ROHLF, 1995). Para detectar diferencias entre la agresividad, tanto intraespecífica como interespecífica, se utilizó la prueba de Kruskal-Wallis, y para determinar entre qué casos particulares se presentaron las diferencias se aplicó la prueba de Nemenyi (ZAR, 1996).
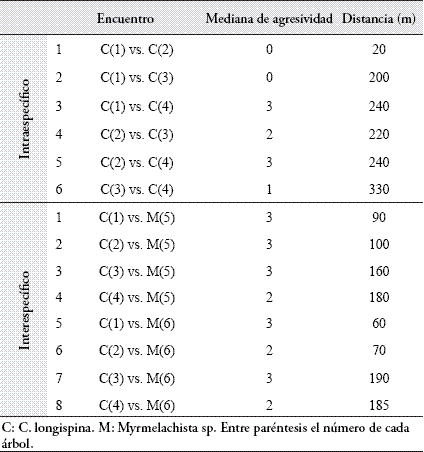
En todas las pruebas de control las forrajeras pertenecientes a un mismo árbol se toleraron entre sí, sin mostrar ningún nivel de hostilidad (Agresividad = 0, tanto para C. longispina como para Myrmelachista sp.). En la Tabla 2 se presentan las medianas de las calificaciones de agresividad para cada encuentro, y las distancias entre árboles.
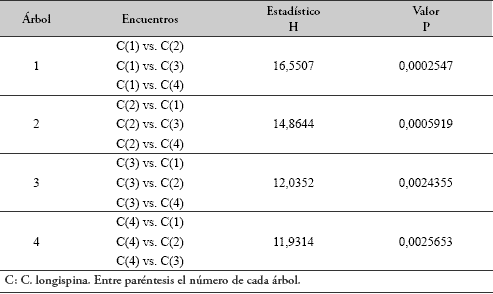
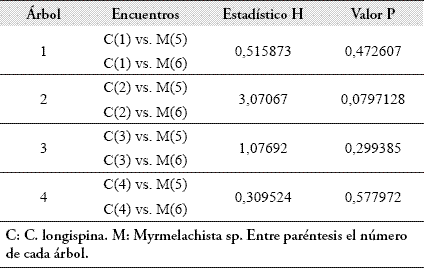
No se encontró correlación entre la distancia que separa los árboles y la agresividad intraespecífica de C. longispina (n=6, rs=0.6269, P=0.1610; τ=0.5930, P=0.1360). Tampoco se encontró correlación entre la distancia que separa a los árboles y la agresividad de C. longispina hacia Myrmelachista sp. (n=8, rs=-0.1690, P=0.6547; τ=-0.1464, P=0.61209). Se hallaron diferencias en las respuestas de agresividad de cada una de las colonias de C. longispina hacia las demás colonias de su misma especie (Tabla 3).
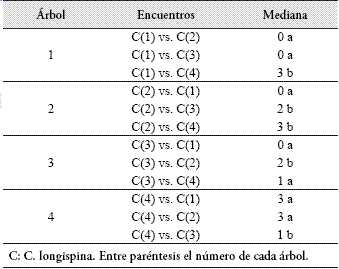
En la Tabla 4 se discriminan los encuentros intraespecíficos que resultaron diferentes. Así, por ejemplo, el nivel de agresividad mostrado por la colonia 1 de C. longispina hacia las colonias 2 y 3 resultó igual, pero diferente al mostrado hacia la colonia 4.
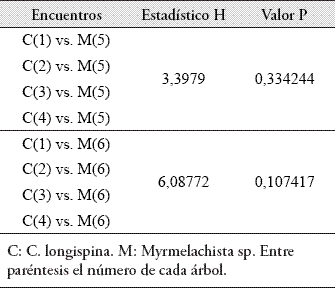
Respecto a los encuentros interespecíficos, no se hallaron diferencias en la agresividad con la que respondía cada colonia individual de C. longispina hacia las dos colonias de Myrmelachista sp. (Tabla 5).
Tampoco se encontraron diferencias entre las respuestas agresivas de las cuatro colonias de C. longispina hacia cada una de las dos colonias de Myrmelachista sp. (Tabla 6).
DISCUSIÓN
Agresividad intraespecífica
En C. longispina no resultó válido el modelo que predice correlación entre la distancia espacial y la agresividad. SATOH & HIROTA (2005) tampoco encontraron relación entre distancia espacial y agresividad en la especie poligínica Camponotus yamaokai. Sin embargo, correlaciones positivas entre el nivel de agresión intraespecífica y la distancia entre nidos han sido reveladas en las especies poligínicas Leptothorax nylanderi (HEINZE et al., 1996) y Formica pratensis (BEYE et al., 1998; PIRK et al., 2001). En F. pratensis, la agresión también fue correlacionada positivamente con la distancia genética entre colonias (BEYE et al., 1998) y con el grado de parentesco intranido de la colonia receptora (PIRK et al., 2001). Estos últimos resultados explican que una colonia poligínica con un alto grado de parentesco intranido podría llegar a funcionar como una colonia monogínica respecto al reconocimiento de los compañeros de colonia, y en ese caso la hostilidad se vería incrementada con la distancia (BEYE et al., 1998; PIRK et al., 2001; SATOH & HIROTA, 2005).
Sin embargo, tanto la ocurrencia de inseminación múltiple como la poliginia pueden generar colonias con diversidad genética intranido que, aún si es moderada, hace necesario un mecanismo de identificación que permita aceptar compañeros de colonia con diferencias genéticas y, asimismo, oponerse a los verdaderos intrusos (HÖLLDOBLER & WILSON, 1990). En muchas especies poligínicas el sistema de reconocimiento de miembros de nido podría estar basado principalmente en el olor ambiental, a diferencia de colonias monogínicas y poligínicas con alto grado de parentesco intranido, donde el componente genético parece ser preponderante (HÖLLDOBLER & WILSON, 1990). Así, por ejemplo, se ha encontrado que colonias de la hormiga poligínica L. curvispinosus separadas entre sí por diferentes distancias, responden entre ellas con distintos grados de agresión, pero después de ser mantenidas en laboratorio bajo condiciones ambientales uniformes se muestran significativamente menos hostiles entre sí (STUART, 1987). Obreras L. nylanderi provenientes de nidos ubicados en diferentes tipos de árbol, exhiben significativamente más agresión entre ellas, comparadas con obreras procedentes de árboles del mismo tipo (HEINZE et al., 1996). También ha sido demostrado un componente principalmente ambiental en los sistemas de reconocimiento de la especie poligínica Solenopsis invicta (OBIN, 1986; OBIN & VANDER MEER, 1988).
Las diferencias entre olores ambientales han sido exploradas en términos del sustrato, materiales para la construcción del nido y diferencias en la dieta (HÖLLDOBLER & WILSON, 1990), condiciones que no se relacionan con la distancia que separa los nidos. En C. longispina, el bajo nivel de agresividad entre las colonias 1 y 2, y entre 3 y 4 (que compartían hábitats parecidos), así como los altos niveles de agresividad de las demás interacciones, parecen sugerir algún tipo de influencia ambiental en el reconocimiento. Sin embargo, de este pronóstico se escapa el resultado del encuentro entre las colonias 1 y 3, que respondieron entre sí con baja agresividad, aunque no compartían hábitats similares.
Por otra parte, el mecanismo de discriminación de parientes (kin discrimination) ha sido planteado como una solución para la convivencia interna de colonias con diversidad genética intracolonial, dado que admite distintas respuestas comportamentales para reaccionar frente a los compañeros de colonia, y una obrera que se encuentra con otra en medio del nido puede, o bien ser indiferente, o escalar por diferentes grados de cooperación en relación con el grado de parentesco que posean (HÖLLDOBLER & WILSON, 1990). Aunque en los enfrentamientos entre forrajeras de C. longispina provenientes de un mismo nido se observaron conductas separadas de tolerancia mutua, acicalamiento y trofalaxia, todas ellas se incluyeron dentro del comportamiento de aceptación y fueron calificadas con 0 en la escala de agresividad. Por eso, esta baja respuesta agresiva no es suficiente para inferir un alto grado de parentesco intranido y, antes bien, la atención diferencial entre obreras parece sugerir cierta diversidad genética intracolonial, que de ser probada, coadyuvaría en la explicación de la independencia entre agresividad y distancia.
Aún cuando se trata de colonias monogínicas o poligínicas con alto grado de parentesco intranido, existen muchos otros factores, además de la distancia espacial, que podrían actuar como fuente de diferenciación genética entre nidos. Estos pueden ser tanto endógenos como exógenos, y entre ellos se cuentan la capacidad de dispersión propia de la especie, la complejidad del hábitat (arquitectura del hábitat), la disponibilidad de hábitats, la estacionalidad y la presencia de obstáculos geográficos, entre otros (BEGON et al., 1995). En el caso particular de especies arbóreas, se ha reportado que los mosaicos de hormigas no son constantes en copas de bosques y plantaciones tropicales, sino que dependen de factores como la competencia con otras especies, la distribución de las especies vegetales hospederas, la presencia de nectarios extraflorales, corpúsculos nutritivos e insectos productores de ligamaza y el mecanismo particular de reproducción de la colonia (DEJEAN et al., 2003).
Agresividad interespecífica
En lo referente a hormigas arbóreas, HÖLLDOBLER (1979) encontró que solamente unas pocas especies de hormigas coexisten en el mismo árbol con hormigas tejedoras del género Oecophylla, y que estas últimas reaccionaban con reclutamiento en masa para defender a sus obreras, cuando individuos de especies competidoras eran puestos en el árbol. HÖLLDOBLER (1983) también halló que obreras de Oecophylla smaragdina que se aventuraron en árboles ocupados por Podomyrma laevifrons, fueron inmediata y fieramente atacadas por obreras de esta última especie. Estos resultados sugieren que especies dominantes en el mosaico arbóreo reclutan grandes fuerzas defensivas, exclusivamente cuando son confrontadas por sus enemigos más peligrosos (HÖLLDOBLER, 1979, 1983; HÖLLDOBLER & WILSON, 1990). La competencia es el tipo más obvio de interacción entre diferentes especies de hormigas, y se la considera la característica primordial de su ecología; de manera que muchas, si no la mayoría de especies, emplean técnicas agresivas que incluyen ataques organizados contra sus rivales (HÖLLDOBLER & WILSON, 1990).
Ahora bien, dado que:
(1) En la zona de estudio C. longispina y Myrmelachista sp. son habitantes de las copas de árboles de yarumo blanco, los cuales son ampliamente conocidos por ser especies mirmecófitas (plantas con estructuras especializadas para alojar hormigas), que producen cuerpos de Müller (DELABIE et al., 2003; HÖLLDOBLER & WILSON, 1990; JOLIVET, 1996).
(2) Los cuerpos de Müller contienen glucógeno, lípidos y proteínas, que se constituyen en una importante fuente de alimento para las hormigas residentes, hasta el punto de que algunas especies dependen enteramente de ellos para su alimentación (HÖLLDOBLER & WILSON, 1990).
(3) Las especies más agresivas defienden estos recursos, extendiendo su territorio muchas veces sobre la planta entera (DEJEAN et al., 2003; DELABIE et al., 2003; HÖLLDOBLER & WILSON, 1990; JOLIVET, 1996).
(4) No se encontró a Myrmelachista sp. en los árboles que ocupaba C. longispina y viceversa.
(5) La mayoría de respuestas agresivas encontradas en los enfrentamientos interespecíficos de un par de forrajeras incluyeron comportamientos de alarma y ataque débil, que en el medio natural constituyen un aviso previo para desencadenar un ataque en masa de mayores proporciones agresivas.
(6) No se encontraron diferencias entre estos niveles relativamente altos de agresividad.
Entonces:
Esto sugiere que la agresividad entre estas dos especies se debe a que son competidoras y cada una de ellas defenderá su territorio de la otra, sin importar la distancia que separe las colonias. Este tipo de interacción de competencia se debe a la superposición de nichos, que coinciden en factores ambientales, sitios de nido y dieta, entre otros parámetros capaces de limitar a cada población (HÖLLDOBLER & WILSON, 1990).
CONCLUSIONES Y RECOMENDACIONES
Este estudio revela que la agresividad intraespecífica en C. longispina es independiente de la distancia que separa los árboles de yarumo blanco que ocupa cada colonia.
Hay razones para sospechar que C. longispina presenta diversidad genética intranido y que basa su sistema de reconocimiento de compañeros de nido en el olor ambiental. Sin embargo, se recomienda la realización de nuevas pruebas para develar el efecto del olor ambiental, como las efectuadas por STUART (1987). Además, sería conveniente incluir las mediciones de los diferentes tipos de comportamiento de aceptación entre obreras provenientes de un mismo nido y contrastarlas con exámenes genéticos, con el fin de confirmar que es posible medir indirectamente el grado de parentesco intranido a partir del grado de aceptación en C. longispina.
Hasta el momento se han analizado otras variables que podrían tener influencia en el fenómeno de agresividad intraespecífica de hormigas poligínicas. Se han reportado correlaciones negativas entre el número de reinas y el grado de agresividad en L. lichtensteini (PROVOST, 1989), S. invicta (MOREL et al., 1990), y Pseudomyrmex pallidus (STARKS et al., 1998). Sin embargo, correlaciones como esta no han sido detectadas en L. ambiguous (STUART, 1991), Rhytidoponera confusa (CROSLAND, 1990), y C. yamaokai (SATOH & HIROTA, 2005). En estos últimos casos, la relación de parentesco entre las obreras del mismo nido podría ser lo suficientemente alta como para que no se produzca una correlación entre el número de reinas y la agresión (STUART, 1991; SATOH & HIROTA, 2005). En especies poligínicas también se han hallado correlaciones positivas entre el tamaño de la colonia y la agresividad (STUART, 1991), y se ha estudiado la relación entre el número de huevos y la agresividad (SATOH & HIROTA, 2005), porque se cree que estos fenómenos son importantes en la competencia territorial y en la defensa contra parásitos sociales (STUART, 1991). También se podría explorar el efecto de estas posibles fuentes de variación en la agresividad intraespecífica de C. longispina, aunque el papel del olor ambiental y la distancia genética intra e internidos parece ser trascendental para el cabal entendimiento de este fenómeno en colonias poligínicas.
En el caso interespecífico, la agresividad encontrada entre C. longispina y Myrmelachista sp. se explica por la existencia de una relación de competencia por recursos entre las dos especies.
Aunque en la literatura especializada en hormigas tanto el género Crematogaster como Myrmelachista se encuentran asociados a plantas del género Cecropia, sorprende la ausencia de estos registros en el ámbito local. Se reconoce sí, la relación de mutualismo obligado entre estas plantas y hormigas del género Azteca.
Queda abierta la posibilidad de investigar las interacciones agresivas de C. longispina y Myrmelachista sp. con otras especies de hormigas que habiten yarumo blanco. Así, por ejemplo, sería muy interesante conocer lo que sucede con las hormigas del género Azteca cuando está presente Crematogaster (un género que ha sido reconocido frecuentemente por ser dominante en mosaicos arbóreos). Igualmente, sería importante develar si C. longispina o Myrmelachista sp. son enteramente dependientes del yarumo blanco para su alimentación, o si emplean complementariamente otro tipo de recursos alimenticios, o estrategias como la depredación.
En las pruebas de agresividad resulta ventajoso el uso de un contenedor pequeño frente a la alternativa tradicional de utilizar cajas de Petri. Una caja de Petri provee un espacio demasiado amplio cuando se trabaja con hormigas pequeñas, lo que dificulta y retrasa el encuentro entre dos obreras. En contraste, un contenedor pequeño reduce el tiempo que tardan las dos obreras en encontrarse, agiliza la observación de sus interacciones y permite la realización de un mayor número de réplicas. Los artrópodos en general presentan un esquema de comportamiento de pautas de acción fija, es decir, liberan una respuesta conductual programada, sólo ante la presencia o ausencia de un estímulo particular. En el caso de las respuestas agresivas en hormigas, estas son liberadas según se reconozcan o no las señales olorosas del contrincante. De modo que se debería utilizar un contenedor nuevo para cada encuentro, o por lo menos, garantizar la eliminación de cualquier rastro de olor que hubiese quedado de encuentros previos.
Los estudios de agresividad en hormigas son de gran importancia, ya que ayudan a comprender la manera como las especies se afectan entre sí: cómo se llevan a cabo las interacciones de competencia, predación, parasitismo, comensalismo y mutualismo, cómo se organizan las comunidades y cómo se forman los patrones de mosaicos de hormigas. Además, si se llega a clarificar la relación entre distancia genética y agresividad, esta última se puede convertir en un rasgo fenotípico importante, de fácil observación, alternativo a complejos y costosos exámenes genéticos, y que permitiría realizar distinciones tanto en el nivel interespecífico (MORI & LE MOLI, 2006a, 2006b) como en el nivel intraespecífico (BEYE et al., 1998; HEINZE et al., 1996; PIRK et al., 2001). Por eso, sorprende la escasez de investigaciones que al respecto se tienen en Colombia. Tal carencia puede explicarse, en parte, por la juventud de la etología como ciencia (MORENO & MUÑOZ, 2007; WRIGHT, 1998/2000), y por el desconocimiento de los aportes que ha entregado a muchas áreas del conocimiento (CASSINI, 1999a, 1999b). Sin embargo, esta situación tiende a revertirse, e internacionalmente cada vez son más abundantes las investigaciones en este campo y se incrementa el número de revistas especializadas en temas comportamentales. La presente investigación constituye, entonces, un gran esfuerzo individual para producir un pequeño aporte al conocimiento del comportamiento de C. longispina y Myrmelachista sp., y se suma al limitado acervo de investigaciones en etología de artrópodos en el país.
AGRADECIMIENTOSAl Dr. John Longino y a mi compañero Hernán Llano Duque, por su ayuda en la identificación de los especímenes. A Efraín Henao y a Juan Alejandro Morales, por las sugerencias y correcciones al borrador de este artículo. A Héctor Chica, por sus recomendaciones acerca del análisis estadístico de los datos. A Jaime Vicente Estévez Varón y Gabriel Jaime Castaño Villa, por gestionar el lugar donde se realizó esta investigación. Finalmente, a la empresa Smurfit Kappa Cartón de Colombia, por permitirme efectuar el estudio en una de las fincas de su propiedad.
BIBLIOGRAFÍA BEGON, M., HARPER, J. & TOWNSEND, C., 1995.- Ecología: Individuos, poblaciones y comunidades. Barcelona: Omega. [ Links ]
BEYE, M., NEUMANN, P., CHAPUISAT, M., PAMILO, P. & MORITZ, R.F.A., 1998.- Nestmate recognition and the genetic relatedness of nests in the ant Formica pratensis. Behavioral Ecology and Sociobiology, 43: 67-72. [ Links ]
CARLIN, N.F. & JOHNSTON, A.B., 1984.- Learned enemy specification in the defense recruitment system of an ant. Naturwissenschaften, 71 (3): 156-157. [ Links ]
CASSINI, M.H., 1999a.- Etología y conservación: un encuentro con futuro. Etología, 7: 1-4. [ Links ]
__________., 1999b.- Importancia de la etología en la Conservación. Etología, 7: 69-75. [ Links ]
CROSLAND, M.W.J., 1990.- Variation in ant aggression and kin discrimination ability within and between colonies. Journal of Insect Behavior, 3: 359– 379. [ Links ]
DAWKINS, R., 1976/1993.- El gen egoísta: Las bases biológicas de nuestra conducta. Barcelona: Salvat. [ Links ]
DEJEAN, A., CORBARA, B., FERNÁNDEZ, F. & DELABIE, J.H.C., 2003.- Mosaicos de hormigas arbóreas en bosques y plantaciones tropicales: 149-158 (en) FERNÁNDEZ, F. (ed.) Introducción a las Hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt. [ Links ]
DELABIE, J.H.C., OSPINA, M. & ZABALA, G., 2003.- Relaciones entre hormigas y plantas: una introducción: 167-180 (en) FERNÁNDEZ, F. (ed.) Introducción a las Hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. [ Links ]
FEENER, D.H., 1986.- Alarm-recruitment behaviour in Pheidole militicida (Hymenoptera: Formicidae). Ecological Entomology, 11 (1): 67-74 [ Links ]
__________., 1987.- Response of Pheidole morrisi to two species of enemy ants, and a general model of defense behavior in Pheidole (Hymenoptera: Formicidae). Journal of the Kansas Entomological society, 60 (4): 569-575. [ Links ]
HEINZE, J., FOITZIK, S., HIPPERT, A. & HÖLLDOBLER, B., 1996.- Apparent dear-enemy phenomenon and environment-based recognition cues in the ant Leptothorax nylanderi. Ethology, 102: 510-520. [ Links ]
HÖLLDOBLER, B., 1979. Territories of the African weaver ant (Oecophylla longinoda [Latreille]): a field study:. Zeitschrift für Tierpsychologie, 51 (2): 201-213. [ Links ]
__________., 1983.- Territorial behavior in the green tree ant (Oecophylla smaragdina). Biotropica, 15 (4): 241-250. [ Links ]
HÖLLDOBLER, B. & WILSON, E.O., 1990.- The Ants. Cambridge: The Belknap Press of Harvard University Press. [ Links ]
JOLIVET, P., 1996.- Ants and plants, an example of coevolution (enlarged edition). Backhuys Publishers, Leiden. [ Links ]
KRAVITZ, E.A. & HUBER, R., 2003.- Aggression in invertebrates. Current Opinion in Neurobiology, 13: 736–743. [ Links ]
MOREL, L., VANDER MEER, R.K. & LOFGREN, C.S., 1990.- Comparison of nestmate recognition between monogyne and polygyne populations of Solenopsis invicta (Hymenoptera: Formicidae). Annals of the Entomological Society of America, 83: 642-647. [ Links ]
MORENO, C.B. & MUÑOZ, J., 2007.- An account on the history of ethology. Suma Psicológica, 14 (2): 213-224. [ Links ]
MORI, A. & LE MOLI, F., 2006a.- The aggression test as a taxonomic tool: Evaluation in sympatric and allopatric populations of wood-ant species. Aggr. Behav., 19: 151-156. [ Links ]
__________., 2006b.- The aggression test as a possible taxonomic tool in the Formica rufa group. Aggr. Behav., 12: 93-102. [ Links ]
OBIN, M.S., 1986. Nestmate recognition cues in laboratory and field colonies of Selenopsis invicta Buren (Hymenoptera: Formicidae): effect of enviroment and role of cuticular hydrocarbons. Journal of Chemical Ecology, 12 (9): 1965-75 [ Links ]
OBIN, M.S. & VANDER MEER, R.K., 1988.- Sources of nestmate recognition cues in the imported fire ant Selenopsis invicta Buren (Hymenoptera: Formicidae). Animal Behaviour, 36 (5): 1361-70. [ Links ]
PIRK, C.W.W., NEUMANN, P., MORITZ, R.F.A. & PAMILO, P., 2001.- Intranest relatedness and nestmate recognition in the meadow ant Formica pratensis (R.). Behavioral Ecology and Sociobiology, 49: 366-374. [ Links ]
PROVOST, E., 1989.- Social environmental factors influencing mutual recognition of individuals in the ant Leptothorax lichtensteini Bondr. (Hymenoptera: Formicidae). Behavioural Processes, 18: 35-39. [ Links ]
SARMIENTO, C.E., 2003.- Metodologías de captura y estudio de las hormigas: 201-210 (en) FERNÁNDEZ, F. (ed.) Introducción a las Hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. [ Links ]
SATOH, T. & HIROTA, T., 2005.- Factors affecting internest variation in the aggressiveness of a polygynous ant, Camponotus yamaokai. Entomological Science, 8 (3): 277–281. [ Links ]
SIEGEL, S. & CASTELLAN, N.J., 1995.- Estadística no paramétrica aplicada a las ciencias de la conducta. 4a ed. Trillas, México. [ Links ]
SOKAL, R.R. & ROHLF, F.J., 1995.- Biometry. 3a ed. Freeman, New York. [ Links ]
STARKS, P.T., WATSON, R.E., DIPAOLA, M.J. & DIPAOLA, C.P., 1998.- The effect of queen number on nestmate discrimination in the facultatively polygynous ant Pseudomyrmex pallidus (Hymenoptera: Formicidae). Ethology, 104: 573–584. [ Links ]
STUART, R.J., 1987.- Transient nestmate recognition cues contribute to a multicolonial population structure in the ant, Leptothorax curvispinosus. Behavioral Ecology and Sociobiology, 21(4):229-235 [ Links ]
__________., 1991.- Nestmate recognition in leptothoracine ants: testing for effects of queen number, colony size and species of intruder. Animal Behaviour, 42: 277-284. [ Links ]
TINBERGEN, N., 1951/1989.- El estudio del instinto. México: Siglo XXI. [ Links ]
WRIGHT, W., 1998/2000.- Así nacemos: genes, conducta, personalidad. Madrid: Taurus. [ Links ]
ZAR, J.H., 1996.- Biostatistical Analysis. 3a ed. Prentice Hall International Editions, Upper Saddle River, NJ. [ Links ]

