










Servicios Personalizados
Revista
Articulo
 Español (pdf)
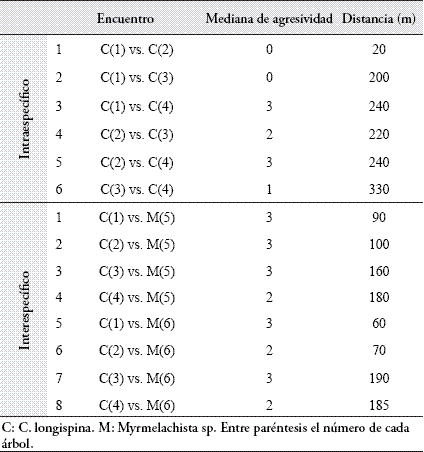
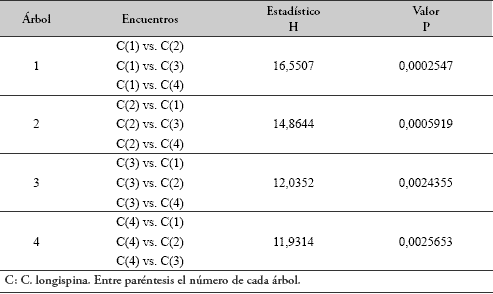
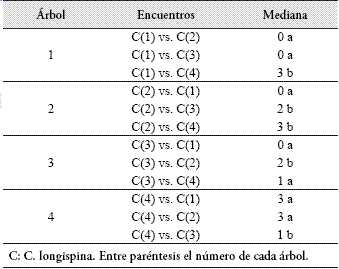
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.13 n.1 Manizales ene./jun. 2009
1 Bióloga. Universidad de Nariño. Ciudad Universitaria Torobajo. Pasto. E-mail: martisaromo@yahoo.com
2 Autor para correspondencia. Biólogo, Msc. Instituto de Ciencias Naturales. Universidad Nacional de Colombia. A.A. 7495, Bogotá. E-mail: aeflorezd@unal.edu.co
Resumen
Este estudio tiene como objetivo analizar la composición, diversidad y abundancia de arañas orbitelares en tres hábitats aledaños a la Laguna Negra, Santuario de Flora y Fauna Galeras, correspondientes a Bosque, Arbustal y Pastizal, identificándose en cada uno de ellos los microhábitats y estratos vegetales ocupados por las arañas orbitelares para la ubicación de sus telas y refugios. La araneofauna orbitelar fue muestreada manualmente mediante la técnica "revelado de telas", que consiste en espolvorear la vegetación con harina para visualizar los hilos; y estuvo conformada por tres familias: Araneidae, Tetragnathidae y Uloboridae, distribuidas en 6 géneros, siendo Chrysometa el dominante. Se detectó además que los hábitats de Arbustal y Pastizal comparten el 77% del total de las especies registradas, indicando más de una comunidad de arañas orbitelares en más de un tipo fisionómico de la vegetación. En cuanto a los estratos, se encontró que el bajo y medio poseen el mayor número de individuos, esto al parecer por la amplia disponibilidad de recursos físicos que les brinda la vegetación para la construcción de las telas. Concluyendo, finalmente, que los principales factores que pueden influir en la distribución y diversidad de la araneofauna orbitelar estudiada, son la diversidad y abundancia de soportes estructurales en la vegetación y la disponibilidad de alimento.
Palabras clave: Arañas, abundancia, estructura vegetal, telas orbiculares.
AbstractThe composition, diversity and abundance of orb-web spiders were studied in three bordering habitats of the Laguna Negra, Galeras Flora and Fauna Sanctuary, corresponding to forest, shrubs, and grassland, identifying in each one the microhabitats and the vegetation stratum occupied by the orb weaver spiders where their webs and shelters are located. The orb weaver spider community was manually sampled by means of the "web revealing" technique, by sprinkling the vegetation with flour in order to visualize the threads. It is composed of three families: Araneidae, Tetragnathidae and Uloboridae, distributed in 6 genera, being Chrysometa the most dominant genus. Furthermore, it was seen that the shrub and grassland habitats share 77% of all the registered species, indicating the presence of more than one orb-web spider community, in more than one physiognomic type of vegetation. Regarding the stratum, both the low and high strata have the majority of individuals, possibly because of the wide availability of physical resources offered by the vegetation for the construction of their webs. Finally, the main factors that can influence the distribution and diversity of the orb-weaver spiders studied are: variety and amount of structural supplies in the vegetation and the availability of food.
Key words: Spiders, abundance, vegetation structure, orb-webs.
INTRODUCCIÓNLas arañas (Araneae) son un grupo faunístico de amplia diversidad y distribución mundial, conquistando la mayoría de los ambientes terrestres (FOELIX, 1996). Dentro del Reino Animal ocupan el séptimo lugar entre los órdenes más diversos con alrededor de 34.000 especies agrupadas en 100 familias y estimando una riqueza del grupo en 170.000 especies (CODDINGTON & LEVI, 1991). Las arañas cumplen un importante papel como reguladoras de una comunidad de artrópodos, pues son carnívoras obligadas donde los insectos son su principal presa, lo que las hace de gran interés económico para los entomólogos. De igual manera, conforman un grupo muy importante para la ecología, ya que las arañas pueden ser agentes indicadores de cambios ambientales (TURNBULL, 1973). Dado que las arañas son predadoras, actúan indirectamente con la vegetación, pues las plantas les suministran escondites y sitios para hacer sus telas (GUNNARSON, 1990). Cabe destacar además que las arañas son de fácil ubicación en el campo gracias a sus telas, y su historia natural, distribución global y taxonomía son las mejor conocidas dentro de los arácnidos (LEVI, 1966).
A pesar de que las arañas son elementos comunes y de gran importancia en el equilibrio ecológico de poblaciones de invertebrados en bosques tropicales, el conocimiento que se tiene de ellas en el neotrópico es aún incipiente (FLÓREZ, 1999). En Colombia, la mayoría de estudios en comunidades de arañas se han realizado en la parte central del país, y en ellos se evaluán aspectos principalmente de tipo ecológico. En arañas orbitelares se conocen investigaciones desarrolladas en bosques andinos, pero son pocas las realizadas en bosques de alta montaña tropical, por lo que se desconocen aspectos como su composición, diversidad y distribución, relacionados con hábitats naturales o de restauración. Teniendo en cuenta estos aspectos, se planteó este estudio, con el fin de investigar la composición de la araneofauna orbitelar en áreas naturales afectadas por cambios estructurales en su vegetación en una zona de alta montaña, y se analizó su abundancia, diversidad, distribución en hábitats y estratos, así como la disponibilidad de alimento.
METODOLOGÍA Materiales empleados
Área de estudio
El trabajo se realizó en la zona suroccidental del departamento de Nariño (1°15´41,16" N y 77° 26´28,73" W), en tres formaciones vegetales aledañas a la Laguna Negra en el Santuario de Flora y Fauna Galeras (S.F.F.G) (Fig. 1), correspondientes a Bosque, Arbustal y Pastizal. El Bosque se encuentra ubicado a 3.470 msnm, dominado por especies vegetales de Weinmannia brachystachya Will. ex Pamp 1905, Clethra fagifolia Kunth 1818 y Miconia spp; en la zona Arbustal (3.320 msnm) se presentan especies de Gaultheria glomerata (Cav.) Sleumer 1935, Gaiadendron punctatum (Ruiz & Pav.) G. Don 1834 y Diplostephium floribundum (Benth.) Wedd 1857; finalmente, en la zona de Pastizal, límite con la Laguna (3.350 msnm), se presentan organismos como Rhynchospora schiedeana Kunth 1837, Cortaderia nitida (Kunth) Pilg 1906 y Tibouchina mollis (Bonpl.) Cogn 1885, principalmente. El área en general presenta un clima húmedo bimodal, con una precipitación promedio anual de 750 mm y una temperatura promedio anual de 13°C (IDEAM, 2004).
Muestreos
El estudio se realizó entre marzo y septiembre del 2003 con salidas mensuales. La intensidad de muestreo fue de dos días por hábitat aplicando un esfuerzo de captura (un colector) de 4h/persona por jornada de muestreo. Se establecieron transectos cuya longitud y dirección varió dependiendo del área de cada zona. La araneofauna se recolectó manualmente y se preservó en alcohol etílico al 70%. Los muestreos se realizaron en jornadas diurnas (08:00-12:00 h) y nocturnas (19:00-23:00 h), aplicando en la jornada diurna la técnica de "revelado de telas" (EBERHARD, 1976), que consiste en espolvorear con harina la vegetación para visualizar los hilos. En la jornada nocturna la captura de los organismos se realizó con ayuda de lámparas y la observación directa de las arañas ubicadas en su tela.
La artropofauna (analizada como disponibilidad de alimento) se recolectó en cada hábitat durante cada salida por los transectos muestreados para la araneofauna. Se realizaron barridos con red entomológica aplicando 250 pases sencillos desde la parte inferior de la vegetación hasta los 2 m de altura.
Se realizó además una caracterización de las estructuras vegetales utilizadas por las arañas para fijar sus telas y ubicar sus refugios, determinándose tres estratos vegetales en cada hábitat: bajo (0-60 cm), medio (61-130 cm) y alto (131-180 cm). La identificación a género de la araneofauna se realizó en el Laboratorio de Entomología de la Universidad de Nariño, con las claves de LEVI (2002) y OPELL (1979). La separación a morfoespecies se realizó a través de la observación de la genitalia y patrones de coloración. Los especímenes fueron depositados en las Colecciones Entomológicas del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia en Bogotá y de la Universidad de Nariño.
Análisis de Datos
Se evaluó la diversidad en cada hábitat, estrato, jornada de muestreo y total de la zona, determinando la abundancia de arañas orbitelares con base en el número de individuos para cada morfoespecie y la riqueza como el número de morfoespecies (MAGURRAN, 1989) para cada uno de los casos. Con el programa Past versión 1.38 (HAMMER et al., 2001) se calcularon los índices de diversidad de Shannon (H´), equitabilidad de Pielou (J´) y disimilitud de especies empleando el índice de Bray Curtis. Con el programa Estimates versión 7.5.0 (COLWELL, 2005) se calcularon las especies esperadas empleando estimadores no paramétricos basados en la incidencia (presencia- ausencia) como Chao 2 y Jacknife de segundo orden (Jack 2) (MORENO, 2001). Se empleó también un análisis de componentes principales (PCA) con el programa NTSYS, versión 2.11 f (ROHLF, 2000), para describir la agrupación de las zonas con base en la composición de la araneofauna orbitelar.
Composición y riqueza de la araneofauna orbitelar
Se recolectaron 453 individuos, agrupados en las familias Tetragnathidae, Araneidae y Uloboridae. Las dos primeras conforman el 99,7% del total de la araneofauna, con 359 y 93 individuos, respectivamente. Se determinaron seis géneros y 17 morfoespecies, y se registró a Chrysometa como el género dominante para la región (288 individuos), mientras que Azilia y Uloborus resultaron ser los géneros menos frecuentes (Tabla 1).


Los hábitats con mayor número de arañas orbitelares fueron Arbustal (149) y Pastizal (197), aportándoles los valores más altos las morfoespecies Araneus sp.2, Chrysometa sp.2, Chrysometa. sp 3, Tetragnatha sp.1, T. sp2 y T. sp3, las tres últimas abundantes en Pastizal; mientras que en Bosque lo hace Chrysometa sp1 (46 individuos) (Fig. 2).

En la tabla 2 se indican los valores de diversidad, el número de especies observadas y los valores de los estimadores (Chao 2 y Jack 2) para cada hábitat, estrato, jornada de muestreo y total de la zona. Los hábitats de Arbustal y Pastizal presentaron la mayor diversidad de arañas orbitelares (H´= 2,17 y 1,94 respectivamente) y la riqueza más alta con 14 y 15 morfoespecies observadas y 17 y 15 esperadas respectivamente (Chao2) (Fig. 3b, 3c); mientras que en Bosque de las 11 morfoespecies esperadas (Chao 2 y Jack 2) se observaron siete (Fig. 3a). La riqueza total observada en la zona fue de 17 morfoespecies, igualando al estimado por Chao 2, pero según el estimador Jack 2 se debió encontrar dos morfoespecies más (Jack 2 = 19) (Fig. 3d).

La similitud de morfoespecies entre los tres hábitats arrojó el valor más alto entre Arbustal y Pastizal (Bray Curtis = 0,77), indicando que la composición de morfoespecies entre estos dos hábitats es muy semejante, ya que su valor es cercano a uno, mientras que los valores más bajos se registraron al comparar Bosque con Arbustal y Pastizal, entre los cuales se comparte aproximadamente la mitad de la araneofauna registrada (Tabla 3).
Distribución vertical
En general, los microhábitats de mayor preferencia por las arañas orbitelares fueron los comprendidos entre la vegetación baja (142) y media (210), cuyo índice de similitud de Bray-Curtis fue de 0,73; es decir, la composición de la araneofauna orbitelar se encuentra distribuida entre estos dos estratos, mientras que entre la parte alta y media su valor es de 0,53 y 0,58 entre el estrato alto y bajo. En la zona de Bosque se observó que la mayor abundancia de arañas se encuentra en el estrato bajo (62) y en las zonas de Arbustal y Pastizal; el estrato medio incluye la mayoría de organismos (80 y 102 respectivamente) (Fig. 4), destacándose a Chrysometa como el género dominante en cada uno de los estratos en todos los hábitats.

En el estrato alto se observaron 14 morfoespecies difiriendo en tres con respecto a Chao 2 (17) y en seis con respecto a Jack 2 (20); en el estrato medio se esperaba encontrar 15 y 17 morfoespecies según Chao 2 y Jack 2 respectivamente, observándose un total de 15; únicamente en el estrato bajo se igualan los valores de morfoespecies observadas con respecto a las esperadas (Chao 2 y Jack 2 = 11).
Patrones de actividad
En la jornada diurna se registraron 233 individuos (Tabla 4) con una diversidad de 1,72 (H´); mientras que en la noche se presentó un valor de diversidad más alto (H´= 2,17) con 220 organismos, representados en 16 morfoespecies observadas, de las cuales se esperaba encontrar 19 y 21 según Chao 2 y Jack 2, respectivamente. En la jornada diurna se esperaba encontrar 16 (Chao 2) y 19 morfoespecies (Jack 2), registrándose 14. Entre estas jornadas, la similitud de especies fue de 0,6 (Bray-Curtis).
Disponibilidad de presas
Se identificó a Diptera con el mayor número de individuos, seguida de Hymenoptera, Araneae, Hemiptera y Coleoptera (Tabla 5). En este grupo de artropofauna se incluyó a las arañas como disponibilidad de alimento, ya que estas pueden presentar depredación intragrupal (COMSTOCK, 1948; GREENSTONE, 1999).
En Bosque se registraron 75 individuos, la abundancia más baja entre las tres zonas de muestreo; mientras que en las zonas correspondientes a Arbustal y Pastizal se recolectaron 250 y 249 ejemplares, respectivamente; así, estos dos hábitats son los más abundantes en cuanto a disponibilidad de presas, a la vez, se presentó una mayor riqueza de artrópodos y se identificaron en Bosque ocho familias, en Arbustal 26 y en Pastizal 23, esto sin contar las posibles otras familias de Diptera que no se lograron identificar.




DISCUSIÓN
La araneofauna orbitelar registrada en este estudio revela a Tetragnathidae como la familia dominante y con mayor riqueza de morfoespecies (nueve), mientras que Araneidae, considerada como una de las más cosmopolitas y de alta riqueza genérica (CODDINGTON & LEVI, 1991) en este estudio, se vio representada únicamente con el género Araneus. Este hecho corrobora lo observado por CEPEDA & FLÓREZ (2007) en un bosque alto andino de la Cordillera Oriental, quienes también reportan a Tetragnathidae como la familia más abundante y sugieren la posibilidad de un reemplazamiento ecológico de Tetragnathidae por Araneidae, esta última dominante en bosques andinos por debajo de los 3.000 msnm.
Con respecto a los géneros, es importante destacar la dominancia de Tetragnatha en la zona límite con la Laguna (Pastizal), confirmando lo descrito por COMSTOCK (1948), como un género común sobre pastos secos y otros objetos cercanos a cuerpos de agua. El género Chrysometa resultó ser el más abundante en los tres hábitats y estratos, considerado por FLÓREZ & SÁNCHEZ (1995) como uno de los que posee el mayor número de especies para Colombia; además, según LEVI (1966), este género es considerado como un miembro neotropical que habita en altas altitudes de los andes y páramos, áreas que concuerdan con los hábitats estudiados que corresponden a formaciones vegetales propias de Bosque alto andino por encima de los 3.300 msnm. Este aspecto quizá incidió en la baja distribución y diversidad de arañas orbitelares para la región, si se considera que estudios como el de BLANCO et al (2003) y FLÓREZ (1998, 1999) realizados en bosques andinos colombianos de menor altitud, registraron la presencia de otras especies y familias del grupo orbicularia.
El 77% de la araneofauna orbitelar registrada en este estudio se distribuye entre las zonas de Pastizal y Arbustal, áreas que por su estado adyacente regenerativo contienen abundantes estructuras vegetales como ramas secas, plántulas, macollas (plantas graminoides), entre otras, de diferentes hierbas y arbustos; al respecto varios autores (RIECHERT & CADY, 1983; RIECHERT & GILLESPIE, 1986) mencionan que la cantidad de estructuras disponibles en el hábitat puede limitar la población de arañas; en este estudio la abundancia y variedad en la forma y dispersión de hojas, tallos y otras formas vegetales permitieron un mayor ofrecimiento de recursos para ser empleados en la fijación de sus telas, lo que se ve reflejado en la alta diversidad y abundancia de la araneofauna orbitelar presente en estas dos zonas.
La estructura del hábitat es muy importante en la selección de un sitio para la tela, tal como lo ha señalado COLEBOURNE (1974), pero también se requiere encontrar cierta demanda espacial (FOELIX, 1996). Como factores que pueden estar incidiendo en la baja riqueza y abundancia de arañas orbitelares en Bosque, se postulan el hecho de que éste ofrece una gran densidad de sustratos vegetales, pero con una dispersión y formas de crecimiento poco adecuados para el anclaje de las telas, aspectos que son considerados determinantes en la ubicación de un sitio para su construcción (JANETOS, 1986; MARSHALL & RYPSTRA, 1999). Una mayor concentración de arañas en el Bosque se registró en las partes bajas de la vegetación, donde se distribuyeron la mayoría de los sustratos como plantas inferiores, tallos, plántulas y troncos caídos ubicados de tal manera que espacialmente fue propicio para la colonización de las arañas orbitelares.
Una situación similar se presentó en Pastizal y Arbustal, los estratos bajos y medios refugiaron a la mayoría de arañas orbitelares (73%), ya que en estos se presentaron espacios entreabiertos con abundancia de soportes físicos de plantas en crecimiento, arbustos y gramíneas, cuyo entrecruzamiento y disposición de hojas y tallos contribuyó a la ubicación de la tela y refugios, reflejando la importancia de la disponibilidad de las estructuras como puntos de fijación. Pero la mayor diversidad de arañas se presentó en las partes altas y medias de la vegetación (H´=2,17 y 2,12 respectivamente), y esto sugiere que existe un desplazamiento vertical en busca de un sitio adecuado para su supervivencia, disminuyendo el grado de competencia interespecífica, equilibrando así la comunidad de arañas orbitelares (J´=0,82).
De otra parte, en los recorridos nocturnos se logró registrar una mayor diversidad y uniformidad de arañas orbitelares que se considera están en estado de actividad, ya que se observaron en su tela; al contrario de lo ocurrido en la jornada diurna, donde por lo general se encontraban en sus refugios o en ocasiones elaborando su telaraña como es el caso de algunas especies de Chrysometa y Araneus; estas últimas consideradas por LEVI (1976) de hábitos nocturnos pero que hacen sus telas durante el día. De esta manera se puede explicar por qué el 60% de las arañas se pueden encontrar en las jornadas diurna y nocturna, teniendo en cuenta además de sus aspectos de comportamiento, la dominancia que estos géneros representan para esta zona.
Finalmente, el suministro de presas constituye un factor importante para la ubicación de la tela (JANETOS, 1986), y se observa que la diversidad poblacional de las arañas tiende a incrementarse en respuesta al incremento de las presas locales (NIFFELER, 1999); razón adicional por la cual puede atribuirse la mayor abundancia y diversidad de la araneofauna orbitelar en las zonas de Arbustal y Pastizal, ya que son sitios donde igualmente se presentó la mayor riqueza y abundancia de la artropofauna disponible como alimento, teniendo en cuenta que esto se puede deber a que estas áreas presentan una gran abundancia vegetal, lo cual puede contribuir a conformar diferentes asociaciones entre artrópodos y plantas, incluyendo así a organismos como fitófagos, perforadores, minadores, masticadores, chupadores, entre otros (MADRIGAL, 2001). Esto favorecería a las arañas porque pueden disponer de un amplio rango de organismos que podrían ser consumidos. Dentro de este grupo se encuentran principalmente Diptera e Hymenoptera, dos órdenes que se presentaron con mayor frecuencia en las telarañas (observación personal) y que a la vez son considerados como presas reales consumidas (BLANCO et al., 2003).
CONCLUSIONESLa distribución de la araneofauna orbitelar para la zona de los alrededores de la Laguna Negra en el Santuario de Flora y Fauna Galeras se concentra en las formaciones sucesionales de la vegetación por la amplia disponibilidad de alimento y la proporción de espacios abiertos y estructuras físicas empleadas como recursos para la fijación de sus telas.
La comunidad de arañas orbitelares estuvo dominada por una alta riqueza de la familia Tetragnathidae, reemplazando posiblemente a Araneidae debido al factor altitudinal.
Los insectos de los ordenes Diptera e Hymenoptera conforman el principal suministro en la dieta de las arañas orbitelares de la región de la Laguna Negra.
AGRADECIMIENTOSEste estudio se desarrolló gracias a la colaboración del personal de la Unidad de Parques Nacionales de Colombia regional Nariño, del Departamento de Biología de la Universidad de Nariño y del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, Bogotá.
BIBLIOGRAFÍA BLANCO, E., AMAT, G. & FLÓREZ, E., 2003.- Araneofauna orbitelar (Araneae: Orbiculariae) de los andes en Colombia: Comunidades en hábitats bajo regeneración. Revista Ibérica de Aracnología, 7: 189-203. [ Links ]
CEPEDA, J. & FLÓREZ, E., 2007.- Arañas tejedoras: uso de diferentes microhábitats en un bosque andino de Colombia. Revista Ibérica de Aracnología, 14: 39-48. [ Links ]
CODDINGTON, J. & LEVI, H., 1991.- Systematics and Evolution of spiders (Araneae). Annual Review of Ecology and Systematics, 22: 565-592. [ Links ]
COLEBOURNE, P.H., 1974.- The influence of habitat structure on the distribution of Araneus diadematus. Journal of Animal Ecology, 43: 401-409. [ Links ]
COLWELL, R., 2005.- Estimates: Statistical estimation of species richness and shared species from samples. Uso de la guía y aplicaciones (en) http://viceroy.eeb.uconn.edu/estimates. [ Links ]
COMSTOCK, J., 1948.- The spider book. Vail Ballou Press. [ Links ]
EBERHARD, W., 1976.- Photography of orb webs in the field. Bulletin of the British Arachnological Society, 3 (7): 200-204. [ Links ]
FLÓREZ, E., 1998.- Estructura de comunidades de arañas (Araneae) en el Departamento del Valle, suroccidente de Colombia. Caldasia., 20 (2): 173-192. [ Links ]
__________., 1999.- Estudio de comunidades de arañas (Araneae) del Parque Nacional Farallones de Cali, Colombia. Cespedesia, 23 (73-74): 99-113. [ Links ]
FLÓREZ, E. & SÁNCHEZ, H., 1995.- La Diversidad de los Arácnidos en Colombia. Aproximación inicial: 327-372 (en) RANGEL, O. Colombia Diversidad Biótica I. Bogotá: Guadalupe Ltda. [ Links ]
FOELIX, R., 1996.- Biology of spiders. Oxford University Press. [ Links ]
GREENSTONE, M.H., 1999.- Spider predation: how and why we study it. The Journal of Arachnology, 27: 227-235. [ Links ]
GUNNARSON, B., 1990.- Vegetation structure and the abundance and size distribution of spruce-living spiders. Journal of Animal Ecology, 59: 743-752. [ Links ]
HAMMER, O., HARPER, D.A.T. & RYAN, P.D., 2001.- Past: Paleontological statistics software package for education and data analysis. Paleontología electrónica, 4 (1): 9. [ Links ]
INSTITUTO DE HIDROLOGÍA, METEREOLOGÍA Y ESTUDIOS AMBIENTALES DE COLOMBIA (IDEAM)., 2004.- Regional Nariño-Cauca. Estación Obonuco, Pasto-Nariño. [ Links ]
JANETOS, A., 1986.- Web- site selection: are we asking the right questions?: 9-22 (en) SHEAR, W., Spiders webs, behavior, and evolution. Stanford University Press, California. [ Links ]
LEVI, H., 1966.- The neotropical orb-weaver genera Chrysometa and Homalometa (Araneae: Tetragnathidae). Bulletin of Museum of Comparative Zoology, 151 (3): 91-211. [ Links ]
__________., 1976.- The orb-weaver genera Verrucosa, Acanthepeira, Wagneriana, Acacesia, Wixia, Scoloderus and Alpaida North of Mexico (Araneae: Araneidae). Bulletin of Museum of Comparative Zoology, 147 (8): 351-391. [ Links ]
__________., 2002.- Keys to the genera of Araneid orbweavers (Araneae, Araneidae) of the Americas. The Journal of Arachnology, 30: 527-562. [ Links ]
MADRIGAL, A., 2001.- Fundamentos de control biológico de plagas. Centro de Publicaciones Universidad Nacional. [ Links ]
MAGURRAN, A., 1989.- Ecological diversity and its measurement. Ediciones Vedrá [ Links ].
MORENO, C., 2001.- Métodos para medir la biodiversidad. Editorial Manuales y Tesis (M&T) SEA. Vol. I. [ Links ]
MARSHALL, S. & RYPSTRA, A., 1999.- Spider competition in structurally simple ecosystems. The journal of Arachnology, 27: 343- 350. [ Links ]
NYFFELER, M., 1999.- Prey selection of spiders in the field. The Journal of Arachnology, 27: 317-324. [ Links ]
OPELL, B.D., 1979.- A key to neotropical genera of Uloboridae. Bulletin of Museum of Comparative Zoology, 148 (10): 443-549. [ Links ]
RIECHERT, S. & CADY, A., 1983.- Patterns of resource use and tests for competitive release in a spider community. Ecology, 64 (4): 899-913. [ Links ]
RIECHERT, S. & GILLESPIE, R., 1986.- Habitat choice and utilization in web-building spiders: 23-48 (en) SHEAR, W. Spiders webs, behavior, and evolution. Stanford University press, California. [ Links ]
ROHLF, F. J., 2000.- Numerical taxonomy and multivariate analysis system. Exeter publishing Ltd. [ Links ]
TURNBULL, A., 1973.- Ecology of the true spiders (Araneomorphae). Annual Review of Entomology, 18: 305-347. [ Links ]

