










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.13 no.1 Manizales Jan./june 2009
CONCISA MIRADA A LA SUBTRIBU MESOSEMIINA STICHEL, 1910, EN COLOMBIA (LEPIDOPTERA: RIODINIDAE)
1 MVZ Centro de Museos, Historia Natural, Universidad de Caldas, A. A. 275, Manizales, Colombia. E-mail: Julianadolfo@hotmail.com
Abstract
This article briefly considers some historical and taxonomic aspects of the members of the Mesosemiina (Lepidoptera: Riodinidae: Riodininae) subtribe in Colombia. The following genera are recorded for Colombia with the probable number of species found: Eunogyra Westwood, 1851 (2); Leucochimona Stichel, 1909 (5); Mesophthalma Westwood, 1851 (1); Mesosemia Hübner, 1819 (59), Perophthalma Westwood, 1851 (2), Semomesia Westwood, 1851 (4) and Teratophthalma Stichel, 1909 (5). Eunogyra satyrus infernalis Salazar & Constantino, a new subspecies originating from Santa Rosa – Rio Villalobos, Cauca, 1350 m is described.
Key words: Colombia, composition, Mesosemiina, Riodinidae, records, kinship, taxonomy.
ResumenEn este artículo se consideran brevemente algunos aspectos históricos y taxonómicos de los miembros de la subtribu Mesosemiina en Colombia. Se registran para el país los siguientes géneros con su probable especiación: Eunogyra Westwood, 1851 (2); Leucochimona Stichel, 1909 (5); Mesophthalma Westewood, 1851 (1); Mesosemia Hübner, 1819 (59), Perophthalma Westwod, 1851 (2), Semomesia Westwood, 1851 (4) y Teratophthalma Stichel, 1909 (5). Se describe a Eunogyra satyrus infernalis Salazar & Constantino, nueva subespecie, procedente de Cauca, Santa Rosa- Río Villalobos, piedemonte este de la Cordillera Oriental de Colombia.
Palabras clave: Colombia, composición, Mesosemiina, Riodinidae, registros, parentesco, taxonomía.
INTRODUCTIONThe family Riodinidae Grote, 1895, comprises small to medium sized butterflies, all brightly coloured, with a generalized pattern which is hiperdiverse in the group. There are a large amount of described species, representing some 140 genera, distributed worldwide but with the greatest diversity in the neotropics (ACKERY et al., 1999). In Colombia there are 1324 riodinid species represented by 126 genera placed in seven tribes and an incertae sedis group of twenty genera waiting for a proper taxonomic and systematic review (CALLAGHAN & LAMAS, 2004). Some of the taxa considered incertae sedis from a systematic point of view have been placed in the subtribe Napaeina by HALL (2003).
Amongst the diverse riodinid groups the subtribe Mesosemiina is one of the most distinctive ones harboring the type genus Mesosemia Hübner, 1819, a conspicuous cluster of riodinids with 122 species known (LAMAS, 2003). This subtribe is represented by seven genera in Colombia, inhabiting both sides of the Andes and also lowland forest habitats. The present paper has the aim to provide some data to the knowledge of this subtribe (sensu stricto) focusing on taxonomy, diversity, relationships of the genera and species recorded in Colombia.
MATERIAL AND METHODS
Material containing voucher specimens was examined in various private and public collections listed in alphabetical order (for the public institutes I follow HEPPNER & LAMAS, 1982, in using acronyms they proposed):
ERH-C: Efraín R. Henao Collection, Villamaria, Caldas; GR-C: Gabriel Rodríguez collection, Envigado, Antioquia; JIV-C: José Ignacio Vargas collection, Villamaría, Caldas; JS-C: Julián Salazar collection, Manizales, Caldas; LMC-C: Luis Miguel Constantino collection, Cali, Valle; MHN-UC: Natural History Museum, Caldas University, Manizales, Caldas; ICN-MHN: Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
Genital dissections were prepared by L. Constantino after macerating and cleaning abdomens in hot 10% KOH solution. The organs were mounted in euparal on slides and examined under optical microscope.
Digital images of the adult specimens were taken by J. Salazar using cameras Nikon D-300, 12 megapixels and Canon power shot A470 of 7.1 megapixels.
The following literature were examined and information gathered concerning riodinids occurring in Colombia: GODMAN & SALVIN (1885), SEITZ (1916, 1924), STICHEL (1910, 1930) and the recent papers by HALL & WILLMOTT (1996), BREVIGNON & GALLARD (1997), DE VRIES (1997), HALL (1999) and HALL & HARVEY (2002).
HISTORICAL APPROACHThe nomenclatural history and taxonomy of the subtribe Mesosemiina was summarized in detail by HALL (2003, 2005) and HALL & HARVEY (2002) on the basis of the classification proposed by D. Harvey in his classic but unpublished PhD dissertation.
The taxon Mesosemiina was erected by H. Stichel in 1910 under the name "Semomesiina", belonging to the subfamily Riodininae (now as family Riodinidae, see LAMAS, 2008).
According to HALL (2005), a small cluster of genera possessing five forewing radial veins was recognized first as Eurybiini by REUTER (1896: 153-154) with Eurybia Illiger, 1850, as type genus. Reuter placed the genera Mesosemia Hübner and Eunogyra Westwood in the tribe Mesosemiidi (= Mesosemiini Bates, 1859). Reuter's classification was based on the examinations of palps and the sensory patches in Eunogyra satyrus (Westwood) and two species of Mesosemia (M. mevania Hewitson and another species hid did not identify).
STICHEL (1930) transferred Eunogyra and Mesosemia with genera Perophthalma Westwood, Mesophthalma Westwood, Leucochimona and Semomesia Westwood to Eurybiini.
On the other hand SEITZ (1916) placed this cluster of genera in the "unterfamilia Erycinidae": Eurybia, Mesosemia, Eunogyra, Diophthalma H.-Schff., Mesophthalma and Perophthalma together with the Napaeina group + the remaining riodinid genera. Seitz in its treatment of the genus Mesosemia lumped the genus Terathopthalma Stichel in the IV group of species and characterized by the following combination of traits: central ocelli large and encircled by red, often angular, males without blue wing scalings. The same author transferred the species of Leucochimona into of the invalid genus Diophthalma Boisduval, 1836 (KIRBY, 1871: 288).
Recently Harvey (quoted by HALL, 2005) redefined Eurybiini to comprise only Eurybia and Alesa Doubleday, chiefly on the basis of Reuter's original palpal character which does no occur elsewhere in the Riodinidae and the tentacle nectary organ present on larval abdominal segments, which otherwise common in the tribe Nymphidiini. Two papers of 1997 and authored by DE VRIES and BREVIGNON & GALLARD on the riodinid faunas of Costa Rica and French Guiana respectively also discussed Mesosemiini. De Vries comments that the tribe is distinguished by the presence of a silk girdle on the pupa that crosses the segment A2; male genitalia has a split base at the pedicel in the abdomen, the eyes hairy and tibial spurs absent in the legs. De Vries cited 130 species and placed them within five genera: the wide ranging Perophthalma, Leucochimona and Mesosemia plus the South American Semomesia and Eunogyra, but Teratophthalma was not cited by him because it does not reach Central America in distribution. In contrast Brevignon and Gallard recorded all of these genera for French Guiana and classified them in the incertae sedis group of Harvey (resolved subsequently by HALL 2003, 2005).
Several works of general interests also illustrated members of the subtribe using specimens deposited in renowned natural history museums around world (eg. LEWIS, 1975; SMART, 1976; VÉLEZ & SALAZAR, 1991; D'ABRERA, 1994, 2001; PIÑAS & MANZANO, 1997). Some of these books placed Mesosemia and Semomesia in the family Nemeobiidae, a younger subjective synonym of Riodiniidae, but the subfamily Nemeobiinae was also in usage (WATSON & WHALLEY, 1975) as well as the tribal name Nemeobiini (REUTER, 1896: 551). Leucochimona species were considered to be representing the genus Mesosemia by GODMAN & SALVIN (1885), but they treated separately Mesosemia and Perophthalma on the basis of characters provided by wing venation, forelegs, palpi and genital organs.
Today the modern treatments of Mesosemiina proposed by HALL & HARVEY (2002) and HALL (2003, 2005) remain the most important contributions so far offering a sound taxonomy of the subtribe. These papers together provide the most firm basis for future studies focused on these butterflies. Evidences were presented by the latter mentioned author for that the Mesosemiina cluster (sensu stricto) has characters that support the monophyly of the subtribal taxon (HALL, 2003).
COMPOSITION OF MESOSEMIINAThe subtribe Mesosemiina has the following synapomorphies representing characters present in adults (nos. 1-4), early stages (no. 6.) and larval hostplant references (no. 5.) as given by HALL (2003, 2005), based on Harvey's analysis:
1. - Eyes setosae (absent in Eurybia).
2. - Eyespot immediately before forewing discal cell end absent in Eunogyra and Perophthalma?).
3. - Multiple narrow bands on wings (absent in Eunogyra and Teratophthalma).
4. - Genital valvae in males triangular with 2 well separated and typically parallel posterior projections and narrow intervening section (absent in Eunogyra and Teratophthalma) (Fig. 8).
5. - Hostplants Rubiaceae, Solanaceae, Acanthaceae (DE VRIES et al., 1992), except Eunogyra in Araceae (BECCALONI et al., 2008).
6. - Silk girdle pupae in abdominal segment.
The following genera are placed in Mesosemiina on the basis of characters given by HALL (2003). The genera are listed in alphabetical order, nomenclatural details are taken from HEMMING (1967):
EUNOGYRA Westwood (1851), in Doubleday, Gen. Diurn. Lep., (2): pl. 72, fig 11. Type species by monotypy: Eunogyra satyrus Westwood (1851). Gen. Diurn. Lep., (2): pl 72, Fig. 11.
LEUCOCHIMONA Stichel, 1909, Berl. Ent. Zeits., 54 (3/ 4) : 273. Type species by original designation: Papilio philemon Cramer (1775). Uitl. Pap. 1(2): 35, pl. 22, figs. 6-7.
MESOPHTHALMA Westwood (1851), in Doubleday, Gen. Diurn. Lep., (2): 455. Type species by monotypy: Mesosemia (Mesophthalma) idotea Westwood (1851), in Dbl., Gen. Diurn Lep., (2): 355, nota.
MESOSEMIA Hubner (1819). Ver. Bekkant Schmett. (2): 21. Type species by selection of Scudder (1875). Proc. Am. Ac. Arts Sci. Boston, 10: 216, Mesosemia philocles Linnaeus (= philoclessa, name invalid).
PEROPHTHALMA Westwood (1851), in Doubleday, Gen Diurn. Lep., (2): 455. Type species by monotypy: Mesosemia (Perophthalma) tenera Westwood (1851), in Dbl. ibid., (2): 455 (as nominal species Papilio tullius Fabricius, 1787), Mantissa Insect., 2: 34.
SEMOMESIA Westwood (1851) in Doubleday. Gen. Diurn. Lep. (2): 455. Type species by selection of Scudder (1875). Proc. Am Ac. Arts. Sci. Boston, 10: 267, Papilio croessus Fabricius, 1777. Gen. Insect: 259.
TERATOPHTHALMA Stichel, 1909, Berl. Ent. Zeits. 54 (1/2): Type species by original designation: Diophthalma phelina C. & R. Felder, 1862. Wien. Ent. Monats., 6: 411.
(Some members of the genera of colombian Mesosemiina are showing in the Fig. 1)

A NOTE ON GENERIC RELATIONSHIPS
In relation with Mesosemiina genera indicated above HALL (2003) suggested that Perophthalma, Mesophthalma, Leucochimona and Semomesia raised from a common Mesosemia ancestor (Table I.) on the basis of the foreleg condition of STICHEL (1910), who proposed that Mesosemiina has a short and swollen femur on the male foreleg, a character, which is not so convincing in relation to the other genera. The Mesosemia adults have four branches in the subcostal venation of the primaries, two before and two beyond the end of the cell. The upper radial and the middle discocellular veins leave the subcostal vein at the same point, so that there is no upper discocellular. In genus Perophthalma the subcostal vein emits only three branches, two before and one beyond the end of the cell. The middle discocellular joins the upper radial vein some way beyond the junction of the latter with the subcostal so that there is a well defined upper discocellular. These differences were remarked by GODMAN & SALVIN (1885) for separate these two genera.
The most impressing evidences to support the monophyly of Mesosemiina are the medially sclerotized and ventrally split male genital pedicel and the setosae eyes present in all genera except in Eunogyra. GODMAN & SALVIN (1885: 451) considered to Perophthalma species an aberrant form of Mesosemia but the distinctive venation of the two genera were also recorded by these authors. In Perophthalma the male secondary organs are very like those of Mesosemia but inside the cavity of aedeagus there is an arrow of serrate papillae. Male Mesosemiina has an approximately triangular shaped genital valva with two well separated posterior projections running parallel, separated by the tegumen "in windows" in all genera. In the genera Teratophthalma and Eunogyra, which are considered as basal taxa, the saccus and the pedicel are ventrally split or medially desclerotized. This last mentioned genus is particular because according to HALL & HARVEY (2002: 185) it possesses specialized androconial organ: the abdominal coremata, which is a pair of eversible tubes of membranous type only present in certain nymphalid subfamilies (Danainae, Satyrinae and Morphinae). Alar androconial organs are frequent in Riodinidae, and in Mesosemiini appear in some Mesosemia species, but in all male Semomesia members. This organ provides important characters and has systematic significance, as it helps the butterfly systematic for elucidating the relationships between the different tribes (HALL & WILLMOTT, 1996; PENZ & DE VRIES, 1999; HALL & HARVEY, 2002).
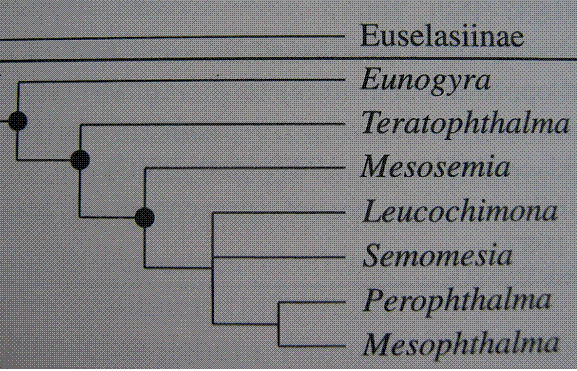
I reproduce in the Table I the generic-level relationships between Mesosemiina genera in the hypothetical phylogeny for the five radial veins tribes of the Riodinidae sensu HALL (2003, 2005). This is based on adult ecology and morphology (wing venation, male genitalia) as well as on early stages. Regarding wing pattern the genera Mesosemia, Semomesia, Mesophthalma and Terathophtalma are similar having an eyespot placed immediately in the forewing discal cell apex with several narrow bands on the dorsal wings surface common in Mesosemia and Semomesia, but in the enigmatic Eunogyra all of these are absent. The adult morphology and wing pattern in Eunogyra and Teratophthalma are atypical in comparison with Mesosemia and their behavior is also somewhat different (BROWN, 1993).
SPECIATIONIn this subtribe six genera display low speciation what probably indicates a primitive lineage, Eunogyra (two species) and Mesophthalma (two species), Perophthalma (three species), Teratophthalma (five species), Leucochimona (nine species) and Semomesia (eight species).
In the contrary the high speciation of Mesosemia with 122 recorded species (LAMAS, 2004) suggests an Amazonian origin with rapid evolutionary development, reflecting in successful radiations to Central America everywhere, including the isolated biogeographic region Chocó in Colombia and the Mata Atlantica in Brazil. The genus Mesosemia is the far well known group of the subtribe, and very well represented in entomological collections. It is widespread from sea level to middle elevations in the Andes (2600 m) and many regions of our country are very rich in species such as Napo (bordering Colombia, Ecuador and Peru) and Tambopata in Peru with 26 species; Jatun-Sacha, Ecuador or Putumayo zone in Colombia with 21 species (SALAZAR, 1995; LAMAS et al., 1996a, 1996b, MURRAY, 1996) and 39 species for French Guiana (BREVIGNON & GALLARD, 1997). The genus is poorly represented in Southeasern Brazil (four-five species) (BROWN & FREITAS, 2000) or in western Ecuador (four species) (RAGUSO & GLOSTER, 1993).
Teratophthalma is restricted to the cloud forest habitats in the Andean region. The species have been recorded in the east slopes of the Oriental Cordillera but the Occidental Cordillera are poorly sampled, and there is evidence to suggest the existence of some undescribed species in montane or remote areas there.
MESOSEMIINA TAXA RECORDED IN COLOMBIA Genus EUNOGYRA Westwood
Hitherto two species were known as representing the genus that inhabits the Amazonian lowland forest: E. satyrus Westwood, 1851, is common in the Amazonas river basin and E. curupira Bates, 1858, has been recorded in the Guianas. There is a newly discovered species that occurs in cloud forests of the eastern foothills of the Oriental Cordillera in Colombia described as follows:
Eunogyra satyrus infernalis Salazar & Constantino ssp. n. (Figs. 2-3)
Male: Forewing length: 18 mm (holotype), upperside entirely black with postmedial row of six black spots very little and insinuated in dark brown background; underside clear brown except with a tin black stripe restricted to medial area from M2 to 1A + 2a space, postmedial area only with one or two black translucent spots. Hindwing upperside entirely black without marks or spots; underside brown, basal area with curved thin black stripe from the costal to anal margin without postmedial row of little black spots (appears in satyrus). Body entirely black

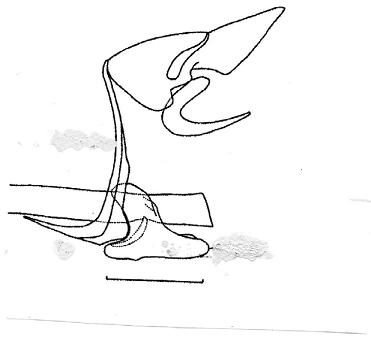
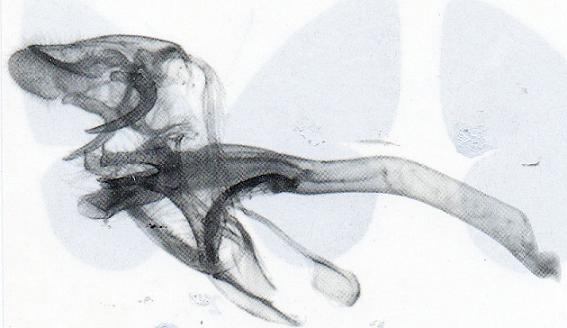
Genitalia: (Fig. 4) Uncus no slender and triangular as satyrus (Fig. 5), more heavy with terminal specula or nail (absent in satyrus); tegumen longer. Gnathos, vinculum and aedeagus similar to nominotypical species, valvae more excavate with terminal part slender.

Female: Unknown.
Etymology: The name is derived from the Latin word "infernalis" in reference to the avernus and the dark coloration of the wings.
Type material, Holotype: male, COLOMBIA, CAUCA: Santa Rosa-Río Villalobos (East slope), 1350 m., 3-III-1993, J. Salazar leg., deposited in the author collection. Paratype: male, (to be deposited in MHN-UC, Manizales) same data and location, J. Salazar leg. (CJS).
Distribution: The subspecies is very local in the premontane rain forest habitats and restricted to the eastern slopes of the Oriental Cordillera flying in deep forest after morning time activity, which is the typical behavior of Eunogyra species (BROWN, 1993: 48). The only congeneric species recorded in Colombia is E. satyrus, a common butterfly that inhabits the entire Amazonian region (PINZÓN, 2008).
Diagnosis and Discussion: E. s. infernalis differs from the most similar congener E. satyrus Westwood, 1851 and E. curupira Bates, 1868 in general by having dorsal pattern entirely black and the postmedial spots of row is far less conspicuous. The infernalis upperside wingsurface is entirely black with postmedial row of six black spots very little and insinuated. These in E. s. satyrus are conspicuous each having yellow rings. Therefore the postmedial area in the underside surface has only one or two black translucent spots, while in E. satyrus there are all black spots evident and remarkable and this is the same for the medial stripe. The hindwing upperside is entirely black without markings or spots in E. s. infernalis, while in E. satyrus the postmedial row of black spots are manifested. The E. s. infernalis hindwing underside anal margin is without postmedial row of little black spots, but these appear in E. satyrus. The new subspecies was previously recorded as E. satyrus by SALAZAR (1995).
Genus TERATOPHTHALMA Stichel
In this genus five large species are known. Three of them are polytypic displaying many subspecies: T. maenades (Hewitson, 1858), T. axilla (Druce, 1904) and T. phelina (C. & R. Felder, 1862) (CALLAGHAN & LAMAS, 2004). According to BROWN (1993) the genus is a typical member of the Amazon Andean forest faunas being active in late morning time and having very low individual. There is a conspicuous sexual dimorphism amongst the known species.
Relatively high Teratophthalma diversity has been recorded in the Andean foothills of eastern and western slopes of the Colombian cordilleras. According to our observations no species of Teratophthalma has been caught in the Central Cordillera. The larval host plants or immature stages are unrecorded (see DÍAS, 1980).
Remarks on some previously described forms
The following species of Teratophthalma were recorded for Colombia by SEITZ (1916) and STICHEL (1930):
1. - The description of the taxon phelina (C. & R. Felder, 1862) was based on a single male specimen from "cordillerae bogotanae" (= eastern cordillera), caught by Lindig (FELDER, 1862: 411). The name minima was proposed subsequently by SEITZ (1916) on the basis of a small male individual from Villavicencio, Meta, which was in the material of Fassl. Is it an individual form, therefore not available, and seems to be close to T. phelina analoga of the same author. This taxon analoga is the Colombian subspecies (SEITZ, 1916) having minute pupilation in the forewing discal cell eyespot but the hindwing similar to that of phelina. The type locality is the upper Rio Negro, 800 m above sea level.
Records: CAUCA: Santa Rosa- Río Villalobos, 1350 m. META: Villavicencio-Bavaria, 500 m.
2. - The taxon monochroma Stichel, 1910 (Fig. 1) was recorded from "Brazil"? by CALLAGHAN & LAMAS (2004). Nevertheless SEITZ (1924, pl. 125a) and D'ABRERA (1994: 931) recorded another stil unnamed taxon taken in Peru under this name. It is characterized by a dark melanised wing surface with a row of seven white spots placed in the marginal area of the forewing and two or three lack marginal white spots of the hindwing. In the both wing ventral surfaces there are bright orange triangular patches or stripes.
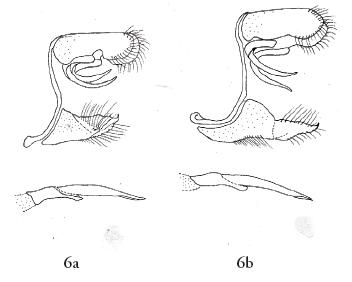
Moreover the name of nigrita (Fig. 6a genitalia) was proposed by SALAZAR (1993) for a similar phenotype based on the holotype male taken in the slopes of the Western Cordillera, in cloud forest habitat. The dark melanised dorsal wing surface shape is entirely black without marginal white spots, and it seems to be identical with the type of monochroma STICHEL (1910). Consequently the two taxa are synonyms, and the name of Stichel has to be applied because it has priority (CALLAGHAN & LAMAS, op. cit.). The hitherto unknown female is illustrated in the plate of this paper (Fig. 1).
Records: RISARALDA: San Antonio del Chami, Quebrada Sutu, 1800 m; Pueblo Rico - Reserva Karagabi, 2100 m.
3. - The taxon maenades Hewitson, 1858 (Figs. 1 male and female, 6b genitalia) was described from "Novelle Grenade" (= Colombia) (KIRBY, 1871). The collecting site is most probably was in the eastern slopes of the Cordilleras, because the species is known only from Cundinamarca and Meta (SALAZAR, 2004: as marsena Hewitson). However, FASSL (1918) recorded maenades to occur also in the western slopes of the same mountain chain. The species is rarely observed. Males perch 2-3 m above the ground in forest trails or occasionally came to feed sand containing urine (J. Vargas, pers. comm.). The typically patterned male has a wide medial white band on the upper side of the forewing but the female has phelina resembling wing pattern with pale white dorsal and ventral markings (Figs. 1 & 14 females).
Records: CAUCA: San Juan de Villalobos, 1350 m. CUNDINAMARCA: Chirajara-Guayabetal, 1200 m. HUILA: Palestina-Jerico, 2200 m. META: Villavicencio-Bavaria, 500 m.
4. - The enigmatical taxon bacche was described by SEITZ (1916: 639) as a subspecific form of maenades. The type "male" (Fig. 7) originates from the Western Cordillera of Colombia (Rio Aguacatal), and represents a distinct species. It differs from similar and allied maenades by the yellow-red illuminated markings and by the forewing upperside white stripe, which is short and reaches only the space between veins M3 and Cu1. The type specimen corresponds with Teratophthalma female phenotype, this it is most probably a female. For this species there is no recent record from Colombia.

5. - The taxon marsidia Hewitson, 1869 has been known from Ecuador (KIRBY, 1871; CAMPOS, 1921), but it also inhabits the western slopes of the Occidental Cordillera, in cloud forest habitats and also occurs locally in the Chocó region. The species is high variable in wing pattern. The taxonomic status and relationships with other taxa representing the maenades group remains to be studied. I illustrate both sexes (Fig 1. male and female).
Records: CHOCÓ: San José del Palmar, 1700 m. NARIÑO: Reserva Natural La Planada, 1850 m. RISARALDA: Pueblo Rico and Santa Cecilia, 800-1900 m.
Genus MESOSEMIA Hübner
This is a "mega diverse" group of small riodinids with pronouncedly sexually dimorphic species. Today there are 122 recognized species which are placed in ten groups by SEITZ (1916) based on Stichel's previous works. The species of the groups IV and VIII have been transferred to the genera Teratophthalma and Semomesia respectively. The group is distributed from Mexico to Northern Argentina (LAMAS, 2003) and inhabits tropical rain forest from the sea level to the 2300-2600 m in the mountains. However, the largest diversity appears to be in the Amazonian region. D'ABRERA (1994: 903) gives general information on adult behavior writing that "they come forth only for a short time for the sake of copula and feeding. Then they perform jerky movements on the leaves, stretching the antennae straight forward placing the hindwings flat on the leaf and slightly raising the forewings. In this manner they hasten from one leaf to another, from one branch to another more jumping that flying" (Fig. 8-9).

In Colombia there are more then 59 recorded species (SALAZAR, RODRÍGUEZ & CONSTANTINO, in prep.).
Genus SEMOMESIA Westwood
Semomesia, a genus allied to Mesosemia contains eight species. All the Semosemia males have androconial patches on the hindwings costa. The sexes strongly dimorphic, the wing upper sides of all males are blue and the females are brown with bluish-white color. The genus has an Amazonian origin and the adult's behavior is similar to that of Mesosemia. According to BREVIGNON & GALLARD (1997), in French Guiana there are four species inhabiting humid hills at low altitude, and they are active between 8 am–3 pm. Some of the species perch 2-3 m above the ground in forest trails.
In Colombia the genus Semomesia is poorly known, there are very few specimens in collections. CALLAGHAN & LAMAS (2004: 146) cited S. croesus lacrimosa (Stichel, 1915) and S. croesus trilineata (Butler, 1874) as members of the Colombian fauna. Some additional records of Semomesia species were provided by FAGUA et al., (1999), SALAZAR et al., (2003) and PINZÓN (2008) listing the following species: S. croesus (Fabricius, 1777), S. macaris (Hewitson, 1859), S. tenella (Stichel, 1910) and Semomesia sp. All these taxa were recorded in the Amazonian region (Amazonas, Caquetá, Meta). Few colour illustrations are based on Colombian examples (VÉLEZ & SALAZAR, 1991; PINZÓN, 2008; and the Humboldt Gallery (H.N.), Villa de Leyva) (Fig. 10).

Genus LEUCOCHIMONA Stichel
This is a small genus of nine species distributed from Mexico, Central America to Nothern Argentina (LAMAS, 2003). The white wings are patterned by brown or grey stripes with characteristic ocelli in the ventral surfaces. In Colombia Leucochimona butterflies are seen in lowland humid forests of all regions up to 2200 m above sea level, although some species reach only 1400-1800 m in the coffee belt (Fig. 11).
Preliminary list of five species occurring in Colombia was provided by CALLAGHAN (1985): L. lagora (H-Schaff) occurs in six Colombian regions; L. aequatorialis (Seitz, 1913), L. icare polita (Stichel, 1910), L. mathisca (Hewitson, 1860) and Leucochimona sp., were recorded from the biogeographic region Chocó; L. icare mathata (Hewitson, 1873) was recorded for the eastern slopes of the Oriental Cordillera, but also from the Western Cordillera (ANDRADE, 2002). Of particular interest is the taxon aequatorialis considered by A. Seitz as a good species apparently from Ecuador. This species was figured in 1924 (pl. 126 e), but no descriptive text appeared in the fifth volume of the "Die Gross-Schmetterlinge der Erde, Amerikanischen Tagfalter" (SEITZ, 1913; LAMAS, 1995: 316). One male L. aequatorialis specimen taken in Colombia (Cachabe?) was illustrated by D'ABRERA (1994: 900). PINZÓN (2008) illustrated L. icare and L. mathisca from the lower Caquetá River and Apaporis-Amazonas.
For L. philemon (= icare) CONSTANTINO (1997) recorded Palicourea sp. and Diodia sp. (Rubiaceae) as hostplants in Colombia.

Genus PEROPHTHALMA Westwood
Three small and distinctive butterfly species are placed in Perophthalma: P. tullius (Fabricius, 1787), P. lasus (Westwood, 1851) and P. lasciva (Stichel, 1929). The type locality of the last mentioned species is in Colombia. Several regional listings contain Perophthalma species records for our country (ARIAS & HUERTAS, 2001; SALAZAR & VAAMONDE, 2002; SALAZAR et al., 2003). ANDRADE (2002: 172) recorded "P. lasus??" in the eastern slopes of the western Cordillera. CALLAGHAN (1985: 63) listed P. tullius for all natural regions of Colombia and indicated the occurrence of P. tullius lasciva only for the Chocó region.
Related with adult behavior GODMAN & SALVIN (1885) comments on P. tullius (= as tenera Ww.) that the species frequents sunny openings in the forest and sometimes to be seen in numbers in one little spot. The individuals rest on the uppersides of the leaves and have much the habit of Mesosemia. (Fig. 12).
DE VRIES (1997) recorded Palicourea guianensis (Rubiaceae) as a hostplants of P. tullius In Costa Rica.

Genus MESOPHTHALMA Westwood
The genus has only one or two tiny species, restricted in distribution to tropical rain forests of the Amazonian drainage and the Guiana shield. The genus is close to Mesosemia in every respects but the small adult size and multistriped wings are diagnostic. CALLAGHAN & LAMAS (2004: 147) cited the taxon mirita Herrich-Schaeffer as an additional species from Surinam together with M. idotea Westwood, 1851. These taxa were considered to be synonymous by SEITZ (1916: 649).
According to BREVIGNON & GALLARD (1997) individuals of Mesophthalma are rare in nature. They observed M. idotea in deep forest between 3-4 p.m. flying low and close to the ground. In Colombia M. idotea is confined to the Amazonas region and several authors recorded the species to the Putumayo zone, Amazonas and the east slopes of Eastern Cordillera (SALAZAR, 1995; CALLAGHAN, 1985). More recently PINZÓN (2008) figured both sexes from the lower Caquetá and Apaporis River (Fig. 13).


ACKNOWLEDGEMENTS
The Natural History Museum of Caldas University (Center of Museum, Manizales) and the following personnel helped in the realization of the present paper by many ways, but particularly with comments, photographs, bibliography, specimens and collections, biological observations and several records: Jason Piers Wilton Hall (Smithsonian Institution, Washington, USA), Gill Carter (USA), Olaf Hermann Hendrix Mielke (Universidade Federal do Parana, Brazil), Dr. Zsolt Bálint (Hungarian Natural History Museum, Budapest), Luis Miguel Constantino (Cenicafe, Chinchina), late Ernesto Wolfgang Schmidt-Mumm (+), Miguel Gonzalo Andrade-C (ICN., Universidad Nacional de Colombia, Bogota), Jorge Eduardo arango, Gabriel Rodriguez, Juan Jaramillo (Envigado, Antioquia) and Efrain Reinel Henao. Jose Ignacio Vargas, Jeisson Cristobal Rios-Malaver and John Curtis Callaghan were of great assistance in field works. Dr. Gerardo Lamas Muller of the Natural History Museum (Universidad Nacional Mayor de San Marcos, Peru) sent generously some photos of type-species and a copy of their insurmountable Checklist of Neotropical butterflies. I express my sincere thanks to all of them.
BIBLIOGRAPHY ACKERY, P.R., JONG, R. & VANE-WRIGTH, R., 1999.- 16. The Butterflies. Hedyloidea, Hesperioidea & Papilionoidea, I: 263-300. In: KRISTENSEN (ed.). Lepidoptera, Moths and Butterflies. Walter de Gruyter, Berlin, NY. [ Links ]
ARIAS, J. & HUERTAS, B., 2001.- Mariposas diurnas de la Serranía de los Churumbelos, Cauca. Distribución altitudinal y diversidad de especies (Lep. Rhop.). Rev. Colomb. Ent., 27 (3-4): 169-176. [ Links ]
ANDRADE, M. G., 2002.- Biodiversidad de las mariposas (Lep. Rhop.) de Colombia: 171-172 (en) COSTA & MELIC (eds.) , PrIBES-SEA, Zaragoza, España. [ Links ]
BECCALONI, G., VILORIA, A., HALL, S.K. & ROBINSON, G.S., 2008.- Catalogue of the hostplants of the Neotropical Butterflies: 536 pp., SEA., NHM, CYTED, IVIC-RIBES, London, UK. [ Links ]
BREVIGNON, C. & GALLARD, J.Y., 1997.- Inventaire des Riodinidae de Guyane Française, II- Riodinidae: Mesosemiini, Eurybiini, incertae sedis. Description de nouveaux taxa (Lep.). Lambillionea, 97: 322-342 + figs. [ Links ]
BROWN, K.S., 1993.- Neotropical Lycaenidae: an Overview. In: NEW (ed.). Conservation Biology of Lycaenidae. Occasional papers IUCN., 8: 45-61, 146-149. [ Links ]
BROWN, K.S. & FREITAS, A.V., 2000.- Diversidade de Lepidoptera em Santa Teresa, Espirito Santo, Brasil. Boll. Mus. Biol. Mello Leitao (NS), 11-12: 7-118. [ Links ]
CALLAGHAN, J.C., 1985.- Notes on the zoogeographic distribution of butterflies of the subfamily Riodininae in Colombia. J. Res. Lepid. Supp. 2: 51-68. [ Links ]
CALLAGHAN, J.C. & LAMAS, G., 2004.- 99 Riodinidae: 141-170. In: LAMAS (ed.). Atlas of the Neotropical Lepidoptera. Checklist, part 4A, Hesperioidea-Papilionoidea. Assoc. Trop. Lep. Gainesville, USA. [ Links ]
CAMPOS, F., 1921.- Estudios sobre la fauna Entomológica del Ecuador 1. Lepidopteros. Rev. Col. Nac. Vicente Rocafuerte, 4: 34. [ Links ]
CONSTANTINO, L.M., 1997.- Conocimiento de los ciclos de vida y plantas hospederas de Lepidopteros diurnos de Colombia como estrategia para el manejo, uso y conservación de poblaciones silvestres. Aconteceres Entomológicos, memorias del I seminario: 57-89. [ Links ]
DE VRIES, P.J. 1997.- The Butterflies of Costa Rica and their Natural History, II. Riodinidae: 282 pp + 25 pls. Princeton University press, NY. [ Links ]
___________., CHACÓN, I. MURRAY, D. 1992.- Toward a better understanding of hostuse and biodiversity in riodinid butterflies (Lep.). J. Res, Lepid., 31(1-2): 103-126. [ Links ]
DÍAS, M.M., 1980.- Morfologia da pupa de alguns Riodinidae brasileiros (Lep.). Rev. Bras. Ent., 24 (314): 181-191. [ Links ]
D´ABRERA, B., 1994.- Butterflies of the Neotropical Region, Riodinidae, 6: vii + 1096 pp. Hill House, Victoria. [ Links ]
__________., 2001.- The Concise Atlas ob Butterflies of the World: 353 pp + figs. Hill House, Victoria. [ Links ]
FASSL, A.H., 1918.- Die vertikale verbreitung der Lepidopteren in der Columbischen Ost-Cordillere. Ent. Rundschau, 35 (1): 1-4, 30-31, 48-50. [ Links ]
FAGUA, G., AMARILLO, A. & ANDRADE, M. G., 1999.- Las mariposas (Lep. Rhop.) como indicadores del grado de intervención en la cuenca del Río Pato (Caquetá): 285-315 (en) Amat, ANDRADE & FERNANDEZ (eds) Insectos de Colombia, II. Ac. Col. Cienc. Ex. Fis. & Nat. U. Nacional, Bogotá [ Links ].
GODMAN, F.D. & SALVIN, O., 1885.- Mesosemia-Perophthalma (in) Biologia Centrali-Americana, I: 378-452, pls. 38-43. London. [ Links ]
HALL, J.P.W., 1999.- A revision of the genus Theope. Its systematic and biology (Lep. Riod. Nymph.): 127 pp + 9 pls. Scient. Publ., Gainesville, USA. [ Links ]
__________., 2003.- Phylogenetic reassessment of the five forewing radial-veined tribes of Riodininae (Lep. Riod.), Syst. Ent., 28: 23-37. [ Links ]
__________., 2005.- A Phylogenetic revision of the Napaeina (Lep. Riod. Mesosem.). Ent. Soc. Washington: 233 pp + figs. [ Links ]
HALL, J.P.W. & WILLMOTT, K.R., 1996.- Systematics of the Riodinid tribe Symmachiini with the description of a new genus and five new species from Ecuador, Venezuela and Brazil (Lep. Riod.). Lambillionea, 96: 637-660. [ Links ]
HALL, J.P.W. & HARVEY, D.J., 2002.- A Survey of androconial organs in the Riodinidae (Lep.). Zool. J. Linn. Soc., 136: 171-197 + figs. [ Links ]
HEMMING, F., 1967.- The Generic names of the butterflies and their type-species (Lep. Rhop.). Bull. Brit. Mus. (N.H.). suppl., 9: 509 pp. [ Links ]
HEPPNER, J. & LAMAS, G., 1982.- Acronyms for world museums collection of insects with emphasis on Neotropical Lepidoptera. Bull. Ent. Soc. Am., 28 (3): 305-315. [ Links ]
KIRBY, W.F., 1871.- A Synonymic Catalogue of diurnal Lepidoptera: 289 pp. John Van Voorst, Patternoster row. [ Links ]
LAMAS, G., 2003.- Las Mariposas de Machu Picchu: 221 pp + pls. 7-34. PROFONANPE, Lima, Peru. [ Links ]
__________., 2008.- La sistemática sobre mariposas (Lep. Hesp. Pap.) en el mundo: estado actual y perspectivas futuras. En: LLORENTE & LANTERI (eds.). Contribuciones Taxonómicas en órdenes hiperdiversos: 57-70. La Prensa UNAM, México. [ Links ]
LAMAS, G., ROBBINS, R.K. & HARVEY, D.J., 1996a.- Mariposas del alto Río Napo, Loreto, Perú (Lep. Pap. - Hesp.). Rev. Per. Ent., 39: 63-74. [ Links ]
LAMAS, G., ROBBINS, R.K., HARVEY, D., MIELKE, O. & CASAGRANDE, M., 1996b.- Taxonomic composition and ecological structure of the rich-butterfly of Pakitza, Manu, Peru: 217-252. In: WILSON & SANDOVAL (eds.). Manu. The Biodiversity of Southern Peru. Nat. Mus. Smith. Contr. Horizonte ed. [ Links ]
LAMAS, G., CAMPOS, L. & RAMÍREZ, J.J., 2003.- Mariposas de la zona reservada Allpahuayo-Mishana, Iquitos, Perú (Lep. Hesp-Pap.): 1-29 + figs. PBIO-IIAP, Iquitos, Loreto, Perú [ Links ].
__________., ROBBINS, R. & FIELD, W., 1995.- Bibliography of Butterflies (in) HEPPNER (eds.). Atlas of Neotropical Lepidoptera, 124: 316. Ass. Trop. Ep. Scient. Publ. Gainesville, USA. [ Links ]
LEWIS, H.L., 1975.- Las Mariposas del Mundo: 312 pp. + figs. Omega, Barcelona. [ Links ]
MURRAY, D.L., 1996.- A Surrey of the Butterfly fauna of Katun- Sacha, Ecuador (Lep. Hesp.- Pap.). J. Res. Lepid., 35: 42-60. [ Links ]
PENZ, C.M. & DE VRIES, P.J., 1999.- Preliminary assessment of the tribe Lemoniini (Lep. Riod.) based on adult morphology. Am. Mus. Novit., 3284: 32 pp. + figs. [ Links ]
PINZÓN, J., 2008.- Mariposas del bajo Río Caquetá y Apaporis (amazonía colombiana): Riodinidae. Watcher Foster & Kaplan prod. & BM. Found. Conservación Internacional Colombia: 5, 17. Bogotá, D. E. [ Links ]
PIÑAS, F. & MANZANO, I., 1997.- Mariposas del Ecuador, Vol. I. Géneros: 115 pp. + figs. Pont Un. Católica de Quito, Ecuador. [ Links ]
RAGUSO, R.A.& GLOSTER, O., 1993.- Preliminary checklist and observations of the butterflies from Maquipucuna field station (Pichincha, Ecuador). J. Res. Lepid; 32:135-161 [ Links ]
REBILLARD, P., 1958.- Contribution a la connaissance des Riodinidae Sud-américains (Ins. Lep.). Mem. Mus. Nat. D´Histoire Nat. (Serie Zool.). 15 (2): 135-266 + 4 pl. [ Links ]
REUTER, E., 1896.- Uber die Palpen der Rhopaloceren: 577 pp + 6 taf. Helsingfors. [ Links ]
SALAZAR, J.A., 1993.- Noticias sobre seis raras especies de Licénidos colombianos. Descripción de una nueva especie de Riodininae para Colombia (Lep. Lyc.). SHILAP, Revta Lepid., 21 (81): 45-53. [ Links ]
__________., 1995.- Lista preliminar de las mariposas diurnas que habitan en el departamento del Putumayo. Notas sobre la distribución en la zona Andina. Colombia Amazónica, 8 (1): 11-69 + figs. [ Links ]
_________., 2004.- Chirajara
.128 años después. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 8:189-200 [ Links ]
SALAZAR, J.A. & CONSTANTINO, L.M., 1993.- Descripción de cuatro nuevas especies de Riodininae para Colombia (Lep. Lyc.). SHILAP, Revta Lepid., 21 (81): 13-18 + figs. [ Links ]
SALAZAR, J.A. & VAAMONDE, C., 2002.- Predicting the overall Butterfly species richness in a tropical montane rain forest in Colombian Choco. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 6: 111-145 + figs. [ Links ]
SALAZAR, J.A., GIRALDO, M. & VARGAS, J.I., 2003. Más observaciones sobre la concentración de mariposas territoriales en cumbres de cerros colombianos y especies residentes en el bosque de Bavaria, Villavicencio- Meta, su distribución y trofismo. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 7: 255-317 + figs. [ Links ]
SALAZAR, J. A., RODRIGUEZ, G., & CONSTANTINO, L. M., 2009.- Contribución al conocimiento del género Mesosemia Hbn., en Colombia y descripción de nuevos taxa. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 13 (2): en prep. [ Links ]
SEITZ, A., 1916.- 1924 (ed.), 8. Familie Erycinidae (in) Die Gross Schemetterlinge der Erde. Amerikanischen Tagfalter, 5: 617-728. Alfred kernen, Stuttgart. [ Links ]
SMART, P., 1976.- Encyclopedie des Papillons: 275 pp + figs. Elsevier Sequoia, Bruxelles. [ Links ]
STICHEL, H., 1910.- Lepidoptera Rhopalocera, fam. Riodinidae Lep. (in) WYTSMAN (ed.) Genera Insectorum, 112A: 1-238., pls. 1-23. Verteneuil & Desmet, Bruxelles. [ Links ]
__________., 1930.- Gattung Mesosemia Hbn., 1816 -Terathopthalma Stich., 1909 (in) Lepidopterorum Catalogus, Riodinidae. Das Tierreich: 253-254, 289-292. [ Links ]
VÉLEZ, J. & SALAZAR, J., 1991.- Mariposas de Colombia: 167 pp + figs. Villegas Editores, Bogotá [ Links ].
WATSON, A. & WHALLEY, P.E.S., 1975.- The Dictionary of Butterflies and Moths in color: 296 pp. + figs. McCraw-Hill book company, Ltd. NY. [ Links ]

