










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.13 no.2 Manizales July/Dec. 2009
* Recibido 10 de marzo de 2009, aceptado 19 octubre de 2009.
1 Department of Zoology, Hungarian Natural History Museum, Budapest VIII, Baross utca 13., H – 1088, Hungary. E-mail: balint@nhmus.hu
2 Museo Entomológico Marcial Benavides, Cenicafé, Chinchiná, Colombia. E-mail: luismiguel.constantino@hotmail.com
3 M.V.Z. Centro de Museos, Historia Natural, Universidad de Caldas, A. A. 275, Manizales, Colombia. E-mail: julianadolfoster@gmail.com
4 Profesores Asociados, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, A.A. 7495, Bogotá, Colombia. E-mail: mgandradec@unal.edu.co
The genus Theorema Hewitson, 1865 is reviewed and characterized. It can be identified on the basis of the following characters: 1) the hind wing dorsal scent patch in the discal and anal areas covered by long hairs; 2) the male genital aedeagus with a terminal sclerotized claw-shaped ventral cornutus and 3) the female genital ductus with a membranous central area weakly sclerotized on its ventral surface. Four species are distinguished: 1) Theorema eumenia Hewitson, 1865 (type species) distributed from south eastern Mexico to Ecuador, 2) Theorema sapho (Staudinger, 1888) Constantino, Salazar & Johnson, 1993, distributed from Panama to Western Colombia and Ecuador, 3) Theorema dysmenia Draudt, 1919, known from Colombia and Peru, and 4) Theorema pyrczia Bálint, sp. n. being described from Venezuela. A key for identification plus a checklist of taxa with reference to type material of Theorema is given. The lectotype of Theorema eumenia is designated. The identity of Thecla werneri Hering & Hopp, 1925, mistakenly considered as Theorema sapho synonym, is clarified and placed in synonymy with Thecla crines H. H. Druce, 1907. Colombian distributional data for Theorema dysmenia, T. eumenia and T. sapho summarized from the literature and various collections are presented. Brief notes on the rediscovery and the biology of T. dysmenia in Colombia, on the biology of T. sapho and the mimicries in which Theorema are probably involved are also provided.
Key words: Theorema, Venezuela, Colombia, taxonomy, new species, new synonymy, lecotype, nomenclature, new record, mimicry.
ResumenEl género Theorema Hewitson, 1865 se revisa y caracteriza en base a las siguientes características: 1) el parche de olor dorsal de las alas traseras en las áreas discales y anales cubiertas por largos bellos; 2) el aedeago con una terminación ventral esclerotizada en forma de garra, y 3) los ductos del órgano genital femenino con un área membranosa central débilmente esclerotizada en su superficie ventral. Se distinguen 4 especies: 1) Theorema eumenia Hewitson, 1865 (especie tipo) distribuida desde el suroriente de México hasta Ecuador, 2) Theorema sapho (Staudinger, 1888) Constantino, Salazar & Johnson, 1993, distribuida desde Panamá hasta el Oriente Colombiano y Ecuador, 3) Theorema dysmenia Draudt, 1919, conocido desde Colombia hasta el Perú, y 4) Theorema pyrczia Bálint, n. sp. descrita en Venezuela. Se presenta una clave de identificación más una lista de taxa con referencia al tipo de material de Theorema. Se designa el lectotipo de Theorema eumenia. La identidad de Thecla werneri Hering & Hopp, 1925, erróneamente considerada como Theorema sapho sinónimo, se clarifica y es ubicada como sinonimia con Thecla crines H. H. Druce, 1907. Se presenta la información distribucional en Colombia para Theorema dysmenia, T. eumenia y T. sapho resumida de la literatura y varias colecciones. También se proveen apuntes sobre el rediscubrimiento y la biología de T. dysmenia en Colombia, sobre la biología de T. sapho y las mímica en las que está involucrada Theorema.
Palabras clave: Theorema, Venezuela, Colombia, taxonomía, nuevas especies, sinonimia nueva, lectotipo, nomenclatura, nuevo registro, mímica.
INTRODUCTIONIn 2006 our Polish colleague Tomasz Pyrcz collected the specimen of a curious lycaenid in north western Venezuela. It was immediately possible to associate it with the genus Theorema Hewitson, 1865 (type species: Theorema eumenia Hewitson, 1865) on the basis of wing shape, colouration and pattern. After dissecting the specimen it turned to be a female. According to the recent catalogue of neotropical butterflies edited by Gerardo Lamas (Robbins, 2004). Theorema is a genus representing the tribe Eumaeini (Theclinae, Lycaenidae) and has three species: 1) the type species, which is known from Mesoamerica to northern Ecuador (Hewitson, 1865; Godman & Salvin, 1887; Staudinger, 1888; Draudt, 1919; dAbrera, 1995), 2) the very rare Micandra sapho Staudinger, 1888 (Constantino, et. al. 1993; Salazar, 1993b) known to occur in Colombia and Ecuador (dAbrera, 1995), and also from Panama (www.americanbutterflies.com), and 3) Theorema dysmenia Draudt, 1919 known only from the Colombian holotype (Salazar, 1993a).
Comparing the known Theorema species with the curious Venezuelan specimen we arrived at the conclusion that it represents an undescribed species. With the discovery of the new Venezuelan species the monophyly of Theorema turned to be ill defined, because of similarities to Eumaeus childrenae (Gray, 1832) in wingshape and pattern. In the literature there are hints that the genus Theorema is not convincingly characterized and should be lumped with Eumaeus Hübner, 1819 (type species: Rusticus minyas Hübner, 1809) (Godman & Salvin, 1887; Staudinger, 1888; then Draudt, 1919 and Goodson, 1947). Consequently we decided to examine characters which could be used to clarify whether or not Theorema is well defined based on morphology.
And even more interesting, just a few months before the capture of the new Venezuelan Theorema species, the curious T. dysmenia has been collected again in eastern Colombia (Andrade-C. et al., 2007).
The purposes of the present paper are 1) to present a new diagnoses and a brief review of the genus Theorema; 2) to compose a key for identification of the species we place in Theorema based on wing characters; 3) to diagnose and describe Theorema species recently taken by Tomasz Pyrcz in Venezuela; 4) to compile a checklist of Theorema with reference to type material; and 5) to provide new distributional records to all the species of Theorema with special regard to Colombia.
MATERIALS AND METHODSOur results are based on the study of the following collections of public institutes and private persons (the abbreviations used in the text are given between brackets, listed here in alphabetical order): Constantino family collection, Cali, Colombia (CFC); Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá, Colombia (ICN: M. Gonzalo Andrade-C.); Magyar Természettudományi Múzeum, Budapest, Hungary (HNHM); Museum National dHistoire Naturelle, Paris, France (MNHN, Jacques Pierre); Natural History Museum, London, England, UK (BMNH; Blanca Huertas); Salazar-Escobar collection, Caldas, Manizales, Colombia (SEC); Further material from the following museums (abbreviations in parentheses) have been also consulted or mentioned directly or indirectly (in this latter case the name of the curator is indicated): Field Museum of Natural History, Chicago (FMNH); Museum für Naturkunde der Humboldt-Universität zu Berlin, Germany (MNHU; Wolfram Mey); Muzeum Zoologiczne Uniwersytet Jagiellonski, Kraków (Zoological Museum of the Jagellonian University) (MZUJ, Janusz Wojtuisak); Naturhistorisches Museum, Wien, Austria (NMW, Sabine Gaal and Martin Lödl); Senckenberg Museum, Frankfurt am Main, Germany (SMF, Wolfgang Nässig); Zoologische Staatssammlung des Bayerischen Staates, München, Germany (ZSM, Axel Hausmann).
Standard lepidopterological etymology was used for descriptive texts, and the applied methods for studying specimens were also the most used ones (Kudrna, 1990; Winter, 2000). After microscopic investigations dissected abdomens (with the genital organs) were placed and stored in a plastic micro vial containing glycerine, pinned on the pin of the relevant specimen. For comparative purpose the following dissections were used listed in alphabetic sequence (with indication to the genital preparation numerical sequence by Zsolt Bálint, HNHM) as follows: Eumaeus childrenae female (1119), Eumaeus childrenae male (1177), Eumaeus minyas female (890), Eumaeus minyas male (889), Theorema eumenia female (1179), Theorema eumenia male (1178), Theorema pyrczia female (1118), Theorema sapho male (1068).
Type concept and nomenclatural acts follow the regulation of the International Commision of Zoological Nomenclature (1999). When labels are described, the mark [//] is used to denote line breaks; words or letters written by italics are handwritten.
In the HNHM an Olympus SZ60 microscope was used during dissections. Androconial and other studies in need higher magnification were carried out under an Olympus SZX12 microscope supported by an OLYPMPUS DP70 digital camera and connected to a Personal Computer.
THE GENUS THEOREMAType species. – Theorema eumenia Hewitson, 1865, by monotypy (Figs. 1-4, 21-22).



Description and Diagnosis. – Wings as in larger eumaeine species with fore wing length 20-25 mm (measured from radial vein erection to vein R3 terminus); fore wing margins all rounded outwardly; hind wing outer margin is round; wing venation commonplace eumaeine with ten fore wing terminal veins, forewing veins R3 and M1 with shared arising point; eye with very short hairs, paraocular area white, frontoclypeal sclerite black with erect scales, vertex covered with brown scales; labial palp with long medial, shorter apical and even shorter basal segment (their comparative size: 1-5-1.5 if the shortest segment is 1), inside covered with white scales; male scent patch present in discal area, and in cell Cu2 in hind wing dorsal surface appearing as pale and less vivid blue coloured area of androconia covered by long hairs (Figs. 30-31). Male genitalia commonplace eumaeine with large brush organ; capsule laterally flat; saccus long with one fourth to one sixth entire genital capsule length; valva with two, upper and lower projection slightly more heavily sclerotized than valval clasp; aedeagus longer than genital capsule with a ratio 7 : 5 (= aedeagus : capsule); external terminus opening dorsally, with two cornutii pointed posteriorly and claw-like; cornutus ventral situated on a mebranous extension of vesica (Figs. 15-16). Female genital ductus with an almost membranous posterior and a sclerotized anterior part divided by a short membranous central area, posterior part with a membranous lamella postvaginalis, anterior part with a slightly scelrotized bursal pouch, central area with a ventral laminated sclerotized part (Figs. 17-18).
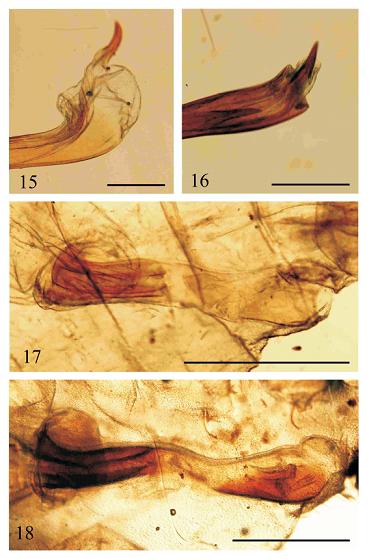

Discussion. – Most probably the generic description we provided above will be supplemented considerably or altered when more information will be available on males and female structures. The following characters are just stressed here by us because of their peculiarities, but we remark, that they are most probably diagnostic on genus level: 1) The male scent patch covered by hairs seems to be characteristic as it is situated in the discal cell of the hind wing dorsal surface (Figs. 1 and 21 for T. eumenia; Figs. 5, 24 for T. sapho and in larger magnification Figs. 30-31). 2) The terminal, strongly sclerotized claw-like ventral cornutus seems to be a peculiar character (Figs. 15-16). We are not aware of any eumaeine genus which has similarly structured and shaped terminal cornutus. As only the males of the species Theorema eumenia and T. sapho are known, having the ventral claw-like cornutus as generic character remains as a hypothesis, which must be checked after the discovery of the not yet known males. 3) Certain female genital structures seem to be also distinctive if we compare those with Eumaeus (see figs. 1c and 2b in CONSTANTINO et al., 1997), especially the weakly sclerotized ventral surface on the membranous central part of the ductus, but at the present stage of our knowledge we are not able to evaluate further this trait on the basis of our observations.

KEY TO THE SPECIES OF THEOREMA
The key is composed for females as the male phenotype in the cases of T. dysmenia and T. pyrczia is not yet known. However we think that key has a general utility for the genus as knowing T. eumenia and T. sapho we presume that wing shape and the basic hind wing ventral pattern of conspecific individuals (being either male or female) is identical.
1) Imagines with filamentous tail at terminus of hind wing vein CuA2 … (2) (T. eumenia group)
- Imagines without filamentous tail at terminus of hind wing vein CuA2 … (3) (T. sapho group)
2) Hind wing dorsal surface with structurally coloured distinct submarginal lunules in spaces between veins CuA and CuA2; ventral hind wing abdominal fold black without pattern in basal and subbasal areas (Venezuela) (Figs. 9-10) … T. pyrczia sp.n.
-- Hind wing dorsal surface with structurally coloured submarginal band in spaces between veins CuA and CuA2; ventral hindwing abdominal fold black with gleaming scales in basal and subbasal areas (Mesoamerica to Ecuador) (Figs. 1-4, 21-22)… : T. eumenia Hewitson, 1865
3) Wing dorsal surface blue; hind wing ventral surface with gleaming blue pattern (Colombia to Ecuador) (Figs. 5-8, 24-25, 27-28) … T. sapho (Staudinger, 1888)
-- Wing dorsal surfaces black with marginal gleaming green intervenial lunules; hind wing ventral surface black without any pattern (Colombia) (Figs 11-14) … T. dysmenia Draudt, 1919

Theorema pyrczia Bálint, sp.nov.
(Figs. 9-10)
Type material. – Holotype female, at moment deposited in the MZUJ (will be deposited in Museo de Zoología Agrícola de la Universidad Central de Venezuela, Maracay). Fore wing length 22 mm (measured from radial vein erection to vein R3 terminus), in good condition, but wings slightly worn, set dorsally, abdomen is missing (dissected), and labelled as 1) VENEZUELA [//] Edo. Trujillo [//] Escuque [//] El Paramito [//] 1550-1600 m [//] 15.IV.2006 [//] T. Pyrcz leg. (white paper, printed); 2) Jagelló Egyetem [//] Prof. Janusz Wojtusiak [//] Kölcsönzött anyag [//] Carlos Prieto hozta [//] 2006. október 30 (white paper, printed); 3) Theorema [//] pyrczi [//] holotípus [//] female [//] gen. prep. No. 1118 [//] det. Zs. Bálint (white paper with black frame, printed [handwritten by Bálint]); abdomen dissected and placed in a plastic microvial attached to the specimen recorded as genital preparation No. 1118 (HNHM).
Description and Diagnosis. – In colouration, pattern and wing shape it resembles to T. dysmenia but there is a filamentous tail at hind wing vein CuA2 as in T. eumenia. The fore wing ventral surface is black with extended white apical scalings with indistinct inner border as also in the mentioned species. The diagnostic features of T. pryczia compared with the female T. eumenia are as follows: 1) the dorsal surface of the fore wing is entirely black with a few gleaming scales in the tornal region (it is sputtered by blue scales with a marked gleaming green tornal area in T. eumenia); 2) the dorsal surface of the hind wing is black with small and isolated bluish green submarginal spots (submarginal spots goldish and create a continuous submarginal band in T. eumenia female); 3) the hind wing ventral surface is similarly patterned as in T. eumenia but the scalings in T. pyrczia is plain white (gleaming in T. eumenia) and 4) the abdominal fold of the hind wing ventral surface is patternless in the basal and subbasal areas (with gleaming scales in T. eumenia). The diagnostic features of T. pryczia compared with the female T. dysmenia are as follows: 1) the hind wing vein CuA2 terminus is tailed (not tailed in T. dysmenia); 2) the wing dorsal surface possess gleaming scales only in the tornal area and in submargins close to tornus (entire submarginal area in both wings coloured structurally in T. dysmenia) and 3) the ventral hind wing surface is patterned (patternless in T. dysmenia).
Etymology. – The holotype was collected by Dr. Tomasz Pyrcz, the eminent Polish specialist on Neotropical pronophilne satryid butterflies. Hence this magnificant species is named after him in rhyme with the congerners Theorema dysmenia and T. eumenia.
CHECKLIST THEOREMA Hewitson, 1865
Ttype species: Theorema eumenia Hewitson, 1865; by monotypy
dysmenia Draudt, 1919 – SMF holotype female (Figs. 11-12), COLOMBIA: Río Negro.
eumenia Hewitson, 1865 – BMNH lectotype male, designated here (see below) (Figs. 21-23), COLOMBIA: New Granada.
-- titania Strecker, 1885 – FMNH holotype female, COSTA RICA: Costa Rica.
-- chiriquensis Niepelt, 1927 – MNHN holotype female, PANAMÁ: Chiriqui.
pyrczia Bálint, sp.n. – MZUJ holotype female (Figs. 9-10), VENEZUELA: Trujillo, Escuque, El Paramito.
sapho (Staudinger, 1888) (Micandra) – MNHU syntype females (see Johnson, 1991: 152), COLOMBIA: Río San Juan.
Lectotype designation of Theorema eumenia
The nominal taxon Theorema eumenia was described by Hewitson (1865: 69, pl. 27, fig. 1: male ventrum, fig. 2: male dorsum), as the type species of the genus. The description was based on an unstated number of male specimens from Colombia (New Granada) deposited in the collection of the author. Cataloging the Hewitson collection, Kirby (1879) recorded two unidentified Theorema specimens from Ecuador and one T. eumenia specimen from New Granada. Subsequently Godman & Salvin (1887: 69) wrote that Hewitson described this [T. eumenia] species from a Colombian specimen, a male. They did not mention the other two Theorema specimens catalogued by Kirby, and they did not state that the Colombian male was in the Hewitson collection. Therefore their words cannot be regarded as an unintent lecotype designation.
One original name-bearing type specimen has been located in the BMNH (Figs. 21-23), which agrees well with the figures supplementing the original description. This specimen is in moderate condition, set dorsally, pinned on a shirt pin in a piece of soft wood, right antenna missing, right wings sligthly damaged (Figs. 21-22), labelled as follows: 1) BMNH(E) # 266272 [white, printed label]; 2) Hewitson Coll. [//] 79-69. 1. [//] Theorema. [//] eumenia Hew. (white, printed label, [handwritten by Kirby]); 3) LECTO [//] TYPE (white confetti label with blue frame, back printed letters); 4) LECTOTYPE male [//] Theorema eumenia [//] Hewitson, 1865 [//] G. Lamas des. 1997 (red, printed label, [handwritten by Lamas]); 5) SYNTYPE male [//] Theorema [//] eumenia [//] Hewitson [//] London, VI. 15 [//] Zs. Bálint, 2000 (red label with black frame, printed, [handwritten by Bálint]) (Fig. 23). This specimen is designated here as lectotype, for objectively fix the identity of the taxon. According to the key given above the species can be easily identified. The specimen bears a lectotype label from Gerardo Lamas but we are not aware about any published action which validates the labelling for nomenclatural purpose.
The female of Theorema eumenia has been described twice as a distinct species. First Stecker (1885) named it as Theorema titania from one example taken by the late Prof. Gabb in Costa Rica, now in Mus. Strecker. This holotype (by monotypy) should be now in FMNH according to Horn et al. (1990: 382). Subsequently the feamle was described again by Niepelt in 1927 as Theorema chiriquensis again on the basis of a single specimen (FAYNEL & BÁLINT, 2004: 275, figs. 18-19).
Notes on Theorema sapho
This species was described on the basis of two female syntype specimens collected in Rio San Juan, Western Colombia by E. Törtsch. The type material is in MNHU. The original documentation is perfect, the species can be recognized unambiguously, although the female figure is captioned as male (STAUDINGER, 1888: 289, plate 97, fig. M. Sapho Stgr. male). The male was described by Hering & Hopp (1925: 196), and illustrated in halftone by Constantino et al. (1993) and Salazar (1993b).
In the butterfly checklist for Atlas of Neotropical Lepidoptera (Robbins, 2004) the name Thecla werneri (Hering, 1925) is listed as junior synonym of Theorema sapho. The taxon Thecla werneri appears as distinct species in the catalogues of Bridges (1988, 1994). It seems that during the compilation of the checklist this name was not critically revised. The taxon Thecla werneri was described by Hering & Hopp (1925: 196). The male holotype (= Typus: 1 male) was deposited in the Hopp collection, which landed in the museum where the type material of sapho has been deposited (Horn et al., 1990: 178). It is mentioned that a similar male was found from Chiriqui (Panamá). We did not see these specimens but on the basis of several characters given in the text (e. g. hind wing with only one tail (= Hfl nur mit einem Schwänzchen), colouration blue somewhat as in laudonia, scent patch brown (= Färbung blau etwa wie bei laudonia, Duftfleck braun), fore wing underside black, with somewhat blue glance, basal third until discalis edge green powdered [scaled] (= Useite der Vfl Schwarz, etwas blauschimmernd, Wurzeldrittel bis zum Zellhrand grün bestäubt), hind wing underside deep blackish brown, base green powdered [scaled] (= Hfl useitig tiefer schwarzbraun, Wurzel grün bestäbt), three to four green crossing lines near the margin (= drei bis vier grüne Querlinien vor dem Saum), we are of the opinion that Thecla crines H. H. Druce, 1907 = Thecla werneri Hering & Hopp, 1925, new synonym.
In the Río Joly diary of Werner Hopp (HERING & HOPP, 1925) there are records for Theorema sapho: on the 1st of June, one male as eine prachtvolle Erycinide, die nicht im Seitz zu finden ist; on the 11th of June, an other male as ein weiteres Exemplar der wunderbaren blauen Erycinide. In the list of recorded species one male and two female sapho specimens are indicated as collected in Río Micay. The female is probably mentioned in the Hopp diary on the 13th of June as das Weib der grossen, vorher erwähnten Thecla, because from Río Micay there is no female Thecla candida (= vorher erwähnten Thecla) in the list. One male and one female of these specimens collected by Hopp can be found now in the Fournier collection of MNHN (Figs. 24-25, 27-28) with pink type labels (Johnson, 1991) (Figs. 26 and 29). Probably these specimens were sold as werneri types, or considered subsequently as type material of Thecla werneri.

THEOREMA IN COLOMBIA
Distributional records. - There is very little information about the geographical, spatial and temporal distribution of the genus Theorema. This stands also for Colombia. The biology of the species is not known. In the followings we summarize the Colombian records we collected from the literature and in various collections as an impetus for studying these magnificent butterflies in the future.
Theorema dysmenia
1. Upper Río Negro, 800 m, (= Colombia) (ZMHU holotype; Draudt 1919).
2. Camino La Almenara, Santa María, 880 m, department Boyacá, 12.X.2005, leg. G. Andrade-C.; No. GAC 10053 (male: ICN-MHN-L19759 in coll. ICN).
Theorema eumenia
3. New Granada (= Colombia) (BMNH lectotype; Hewitson 1865).
4. Nare (= Puerto Nare), department Boyacá (male: MNHN, general collection).
5. Muzo, department Boyacá, 1924, Apollinaire María (male: MNHN, Fournier collection); ditto, 1929 (male: MNHN, Fournier collection).
Theorema sapho
6. Río Micay-Joly, department Chocó, 23.V.-18.VI.1924, leg. Werner Hopp (male, female: MNHN, Fournier collection).
7. Río Tatabro, Bajo Anchicayá, 200 m, department Valle, 7. VIII.1985, leg. Luis M. Constantino (male: CFC); ditto, leg. Julián A. Salazar (male: SEC); ditto, 150 m, 10. IV. 1989, leg. Luis M. Constantino (female: CFC).
8. Yatacué, Alto Anchicayá, 850 m, department Valle, 3.VIII.1983, leg. Luis M. Constantino (female: CFC). Santa Cecilia, 500 m, department Risaralda, leg. Julián A. Salazar (female: SEC).
Notes on the rediscovery of Theorema ysmenia
This species is characterized by solitary habits, preferring the middle and upper strata of the forest interior. Sometimes individuals are found in clear sunny spots on the ground pudling or flying along the trails. Occasionally the species can be found on the ground with wings widespread.
We were informed via personal communication by Gerardo Lamas (Lima, Perú), that the species has been also collected in southern Peru. We did not seen any material.
Notes on the biology of Theorema sapho.
This beautiful species is found in habitats indicating well preserved rainforest ecosystem. The male flies high in the forest canopy very fast, therefore it is seldom to see in the understory. Individuals were attracted down to the ground with a decoy made of blue metallic paper used to collect Morpho butterflies. On the other hand females fly low above the ground and slow along trails in the forest together with her co-mimic, Brevianta saphonota (Constantino, Salazar & Johnson, 1993). These are the only hitherto known lycaenidae found in the neotropics with a characteristic wide white band on the upper wings resembling the unpalatable Heliconius cydno zelinde, and H. sapho chocoensis involved in a mimicry ring in the pacific slope (W. Colombia). Hitherto it is only the second documented case for the family Lycaenidae in the Neotropics to be involved in mimicry (see CONSTANTINO et al. 1993; BENYAMINI, 1995). The name sapho was chosen by STAUDINGER (1888) to relate T. sapho with the similar white band pattern found in Heliconius sapho.
On the mimicries which possibly involve Theorema
For a further evaluation of the mimicry which involves T. sapho (and B. saphonota) (see CONSTANTINO et al., 1993) we mention that the light (yellow or white) banded forewing dark phenotype is very widely distributed amongst female neotropical butterflies. The optical signal generated by white wingtips is described by a naturalist manner very recently (NEILD, 2008: 1199-1200).
The phenomenon raises the question whether this phenotype indicates indeed a special mimicry ring, or it is a general advertisement for the members of the ecosystem in the neotropics, that the individual which possesses such a pattern is a female. Or, alternatively it is a fixed and generalized warning pattern for unpalatable species, which were used to develop mimicry rings. We list in the followings some genera which are involved in the sapho phenomenon. Our list is not complete, probably certain examples are even not relevant and some taxa have been overlooked, but at least it gives a hint how widespread is the sapho phenotype.
Nymphalidae: Apaturinae (Doxocopa); Nymphalidae: Biblidinae (Dynaminae, Eunica, Hamadryas, Marpesia); Nymphalidae: Brassolidae (Opsiphanes); Nymphalidae: Charaxinae (Fountainea, Hypna, Memphis); Nymphalidae: Heliconiinae: Acraeini (Actinote diceus); Nymphalidae: Nymphalinae: Melitaeini (Castilia occidentalis); Nymphalidae: Satyrinae: Pronophilini (Pedaloides). Papilionidae: Papilioninae, Troidini (Parides). Riodinidae (Brachyglenis, Calospila, Cyrenia, Ithomeis, Esthemopsis, Lypericornis, Melanis, Mesosemia, Mimocastnia, Rhetus, Semomesia, Setabis, Stichelia, Xenandra, Teratophthalma, Voltinia).
The female phenotypes of Theorema dysmenia, T. eumenia and T. pyrczia suggest that the genus is probably involved in another mimicry complex. In the Neotropical Region the black dorsal hind wing surface sometimes associated with structurally coloured (often metallic green or strongly reflective white) submarginal area. Obvious examples are: Nymphalidae: Biblidinae (Asterope, Catacore, Diaethria, Perisama); Nymphalidae: Heliconiinae (Heliconius); Lycaenidae: Theclinae (Eumaeus); Riodinidae (Pandemos, Setabis, Uraneis, Xenandra). However it cannot be excluded that this phenotype reflects that this contrasting appearance is an optical tool developed independently by the various Lepidoptera genera involved. Finally we give the remark, that the trait of ventrally red coloured abdomen so typical for some Neotropical eumaeine lycaenid genera (for example Atlides, Eumaeus, Thecloxurina), which most probably advertises toxicity, lacks in Theorema (and also in Eumaeus childrenae). It indicates that the larval hostplant of the genus is not poisonous.
*Editor´s Note:
J.A. Salazar believe that the figure of the holotype of Th. dysmenia is a male, no female as well in known in the original description of DRAUDT (1919). The true female of the species appear in the figures 13-14 (plate 2) of this paper.
We express our sincere thanks to the curators, mentioned in the part of Materials and Methods that for the access of the collections under their care or for certain information. We thank for Dr. Carlos Prieto and for an anonymous reviewer for their constructive comments.
BIBLIOGRAPHY ANDRADE-C., M.G; CAMPOS-SALAZAR, L.M; GONZÁLEZ-MONTAC, L-A. & PULIDO, H., 2007.- Santa María, Mariposas Alas y Color. Universidad Nacional de Colombia-I.C.N., Bogotá, Serie Guías de Campo No. 2. 248 p. + figs. [ Links ]
BENYAMINI, D., 1995.-Synopsis of Biological Studies of the Chilean Polyommatini (Lepidoptera, Lycaenidae). Reports of the Museum of Natural History, University of Wisconsin, Stevens Point, 52: 1-51. [ Links ]
BRIDGES, CH., 1988.- Catalogue of Lycaenidae and Riodinidae (Lepidoptera: Rhopalocera). Published by the Author, Urbana, Illinois. 788 p. [ Links ]
___________., 1994.-Catalogue of the family-group, genus-group, and species-group names of the Riodinidae and Lycaenidae (Lepidoptera: Rhopalocera) of the World. Published by the Author, Urbana, Illinois. 1128 p. [ Links ]
CONSTANTINO, L. M., SALAZAR ESCOBAR, J. A. & JOHNSON, K., 1993.- Theorema sapho (Staudiner) and Two Unusual New Species of Theclinae from Colombia (Lepidoptera, Lycaenidae, Theclinae). Reports of the Museum of Natural History, University of Wisconsin, 41: 1-5. [ Links ]
_________________, & JOHNSON, K., 1997.- A new species of the eumaeus toxana / toxea clade from tepui´s in the amazon basin of colombia. rev. theclinae colomb.,1 (8): 9P. [ Links ]
D'ABRERA, B., 1995.-Butterflies of the Neotropical Region. Part VII. Lycaenidae. Victoria Black Rock, Hill House. p. i-xi + 1098-1270, figs. [ Links ]
DRAUDT, M., 1919.- Familie Lycaenidae: p. 744-831, pls. 144-159 (in) SEITZ, A. (1910-1924) Die Gross-Schmetterlinge der Erde. 5. Band. Amerikanische Tagfalter. Alfred Kernen Verlag, Stuttgart. 1139 p., 194 pls. [ Links ]
FAYNEL, C. & BÁLINT, ZS., 2004.- Supplementary information on Eumaeini primary type material and further historical specimens deposited in the Museum National d'Histoire Naturelle, Paris. Bulletin de la Société Entomologique de France, Paris, 109 (3): 263-286, 16 figs. [ Links ]
GODMAN, F. D. & SALVIN, O., 1887.-Biologia Centrali-Americana. Insecta. Lepidoptera-Rhopalocera. London, Duleau & Co., Bernard Quartrich 2, 1-112 p., pls. 48-56. [ Links ]
GOODSON, F. W., 1947.- Notes on the genus Eumaeus Hübner (Lep., Lycaenidae). The Entomologist, 80: 273-276. [ Links ]
HERING, M. & HOPP, W., 1925.- Eine Sammelausbeute des Herrn Werner Hopp aus dem Chocó Kolumbiens. Deutsche entomologische Zeitschrift Iris, 39: 181-207, 9 figs. [ Links ]
HEWITSON, W. C., 1865.- Illustrations of diurnal Lepidoptera. Part I. Lycaenidae. London: John Van Horst, (2): 37-76, pls. 17-30. [ Links ]
HORN, W., KAHLE, I., FRIESE, G. & GAEDIKE, R., 1990.-Collectiones entomologicae. Ein Kompendium über Verbleib entomologischer Sammlung der Welt bis 1960. Berlin, Akademie der Landwirtschaftswissenschaften der Deutschen Demokratischen Republik. p. 1-220 (Teil I: A bis K), 221-573 (Teil II: L bis Z). [ Links ]
INTERNATIONAL COMMISSION ON ZOOLOGICAL NOMENCLATURE., 1999.-International Code of Zoological Nomenclature, Fourth Edition adopted by the International Union of Biological Sciences. London: The International Trust for Zoological Nomenclature. xxix + 306 p. [ Links ]
JOHNSON, K., 1991.-Types of neotropical Theclinae (Lycaenidae) in the Muséum National d'Histoire Naturelle, Paris. Journal of the Lepidopterists' Society, 45 (2): 142-175. [ Links ]
KIRBY, W. F., 1879.-Catalogue of the Collection of Diurnal Lepidoptera formed by the late William Chapman Hewitson of Outlands, Walton-on-Thames; and bequeathed by him to the British Museum. London: John van Voorst. 246 p. [ Links ]
KUDRNA, O., (ed.)., 1990.- Introduction to Lepidopterology (in) Butterflies of Europe. Volume 2. Wiesbaden: Aula-Verlag. 557 p. [ Links ]
NEILD, A., 2008.-The Butterflies of Venezuela. Part II, Nymphalidae II (Acraeinae, Libytheinae, Nymphalinae, Ithomiinae, Morphinae). London: Meridian Publications. 275 p. [ Links ]
ROBBINS, R. K., 2004.-Introduction to the Checklist of Eumaeini (Lycaenidae): xx-iv-xxx; Lycaenidae. Theclinae. Tribe: Eumaeini: 118-137 (in) LAMAS, G. (ed.) Checklist: Part 4A. Hesperioidea - Papilionoidea (in) HEPPNER, J. B. (ed.) Atlas of Neotropical Lepidoptera, Volume 5A. Gainesville, Association for Tropical Lepidoptera, Scientific Publishers. xxvi + 439 p. [ Links ]
SALAZAR ESCOBAR, J. A., 1993a.- Una lista comentada de algunas especies de mariposas de distribución restringida o locales en Colombia (Lepidóptera: Rhopalocera). SHILAP Revista de lepidopterología, 21 (81): 33-46. [ Links ]
____________________., 1993b.- Noticias sobre seis raras especies de licénidos colombianos. Descripción de una nueva especie de Riodininae para Colombia (Lepidóptera: Lycaenidae). SHILAP Revista de lepidopterología, 21 (81): 47-53. [ Links ]
STAUDINGER, O., 1888.- Exotische Schmetterlinge. Theil I. Band 1 und 2. Fürth, G. Löwensohn. iv + 333 p., 100 pls., 1 map. [ Links ]
STRECKER, F. H. H., 1885.- Descriptions of new Species of Lepidoptera. Proceedings of the Academy of Natural Sciences of Philadelphia, 37: 174-179. [ Links ]
WINTER JR., W. D., 2000.-Basic Techniques for Observing and Studying Moths & Butterflies. The Lepidopterists' Society, Los Angeles, 443. [ Links ]

