










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.13 n.2 Manizales jul./dic. 2009
DIVERSIDAD DE ESCARABAJOS (COLEOPTERA, STAPHYLINIDAE) EN DOS LOCALIDADES DEL DEPARTAMENTO DEL QUINDÍO*
ROVE BEETLES (COLEOPTERA, STAPHYLINIDAE) DIVERSITY IN TWO LOCALITIES IN THE DEPARTMENT OF QUINDIO
Diana M. Méndez-R1, Margarita M. López-G2. y Rocío García-C3
* Recibido 10 de marzo de 2009, aceptado 23 de octubre de 2009.
1 Estudiante Programa de Biología, Universidad del Quindío. E-mail: dianamendez04@gmail.com
2 Estudiante Programa de Biología, Universidad del Quindío. E-mail: margaralopezg@gmail.com
3 Docente Programa de Biología, Universidad del Quindío. E-mail: rociogarcia06@yahoo.es
Se comparó la composición de estafilínidos de un bosque del Cañón del río Barbas y el bosque secundario suburbano, Sendero Cedro Rosado, empleando necrotrampas y escrutinio visual de 1 m2 hojarasca, intercaladas a lo largo de 3 transectos de 2 x 100 m, para un total de 66 unidades de muestreo. Se colectó un total de 801 individuos separados en 79 morfoespecies, 47 géneros y 8 subfamilias. Las subfamilias más representativas en cuanto a riqueza de especies fueron Aleocharinae (25,3%), Pselaphinae (17,7%), Staphylininae (15,2%) y Paederinae (12,6%). El Cañón del río Barbas presentó la riqueza más alta (44 especies). Sin embargo, no se encontraron diferencias significativas entre los bosques, en cuanto a la riqueza (U=-1,22, p=0,21) y abundancia de especies (U=-0,835, p=0,403) soportado por los índices de similitud de Jaccard y Sorensen-cuantitativo. Las necrotrampas y el escrutinio de hojarasca muestran diferencias significativas (U=4,67, p=0,0002) ya que el número de especies exclusivas por cada método fue mayor que el número de especies compartidas por ambos. Este estudio constituye el primer aporte sobre la diversidad de estafilínidos en el departamento del Quindío.
Palabras clave: estafilínidos, estructura y composición de comunidades, necrotrampas, hojarasca.
AbstractStaphylinidae composition was compared in a forest of the Cañón del río Barbas and the Sendero Cedro Rosado suburban secondary forest, using carrion traps and visual screening of 1 m2 of leaf litter. These methods were interspersed among three 2 x 100 m transects, for a total of 66 sampling units. A total of 801 individuals, 79 morphospecies, 47 genera and 8 subfamilies were collected. The most abundant subfamilies were Aleocharinae (25.3%), Pselaphinae (17.7%), Staphylininae (15.2%) and Paederinae (12.6%). Although the Cañón del río Barbas had the highest richness (44 species), there were no significant differences in species richness (U=-1.22, p=0.21) and abundance (Z=-0.835, p=0.403) between forests. Said information was supported by the Jaccard and Sorensen- quantitative similarity indexes. The carrion traps and leaf litter show significant differences (U=4.67, p=0.0002), since the number of exclusive species in each method was higher than the number of shared species. This study constitutes the first contribution on rove beetle diversity in the department of Quindío.
Key words: rove beetles, community composition and structure, carrion traps, leaf litter.
INTRODUCCIÓNColeóptera se ha considerado como el orden más rico y diverso, no sólo de Insecta sino de toda la fauna existente (MARTÍN, 1997; SKALSKI & POSPIECH, 2006). Dentro de este orden, Staphylinidae con 46.275 especies descritas hasta 1998, es una de las familias más representativas y diversas biológicamente (BLACKWELDER, 1936; NEWTON, 1990; KLIMASZEWSKI et al., 1996; NEWTON et al., 2000; NAVARRETE-HEREDIA et al., 2002; NEWTON et al., 2005; GUTIÉRREZ-CHACÓN & ULLOA-CHACÓN, 2006). Sin embargo, se estima que hasta ahora sólo se ha descrito una cuarta parte de las especies tropicales (NAVARRETE-HEREDIA et al., 2002). Del mismo modo, para Colombia, en el 2005 se reportó un total de 796, pero se calcula que la fauna de estafilínidos en el país puede estar constituida por unas 5.000 especies (NEWTON et al., 2005).
El alto número de especies de estafilínidos se relaciona con una alta diversidad ecológica que se evidencia en la gran variedad de hábitats, hábitos alimenticios y comportamientos, lo que les ha permitido ser uno de los grupos más exitosos (KLIMASZEWSKI et al., 1996; AL DHAFER, 2003). Dado que cerca de la mitad de sus especies se encuentra en suelo y hojarasca, esta familia forma uno de los componentes de insectos más importantes ecológicamente en la fauna edáfica (NEWTON, 1990), influyendo en procesos como la descomposición de materia orgánica y el ciclado de nutrientes (SKALSKI & POSPIECH, 2006).
Teniendo en cuenta la gran diversidad de la familia Staphylinidae y los pocos estudios realizados hasta el momento en Colombia (GARCÍA et al., 2001; GARCÍA & CHACÓN, 2005; GUTIÉRREZ-CHACÓN & ULLOA-CHACÓN, 2006), la comparación de la diversidad de estafilínidos en el bosque secundario suburbano del Sendero Cedro Rosado y el bosque primario del Cañón del río Barbas es un aporte significativo al conocimiento de la diversidad de la región.
De esta manera, los objetivos de este trabajo fueron caracterizar la composición de estafilínidos en ambas localidades, comparar su diversidad en cada uno de los bosques, y evaluar la eficiencia de los métodos de colecta empleados en el muestreo.
MATERIALES Y MÉTODOSEl Sendero Cedro Rosado (SCR: 4º3240 N, 75º4613 W) es un bosque secundario intervenido de aproximadamente 8 hectáreas (CASTRO, 1995), que se encuentra en la zona suburbana de la ciudad de Armenia (Quindío), a una altura de 1450 msnm. El Cañón del río Barbas (CRB: 4º4226,1 N, 75º3911 W), se encuentra ubicado a una altura de 1869 msnm, en el municipio de Filandia; su vegetación se caracteriza por la presencia de taxa típicos de bosques subandinos, con un dosel entre los 20 m (MENDOZA et al., 2007).
El muestreo en el SCR fue realizado entre Junio-Julio de 2007 y en el CRB en Marzo de 2008. En ambos bosques, se realizaron tres transectos de 100 x 2 m separados por 100 m. En cada transecto se establecieron dos métodos de colecta, necrotrampas y cernido de 1 m2 de hojarasca, intercaladas cada 10 m, para un total de 66 unidades de muestreo (GARCÍA et al., 2001; GARCÍA & CHACÓN, 2005). Las necrotrampas fueron empleadas con atrayente (pescado descompuesto) y se dejaron actuar por 24 horas.
Los especímenes colectados fueron determinados y almacenados en el Museo de Artrópodos de la Universidad del Quindío (MAUQ), hasta el nivel taxonómico más bajo posible, usando claves dicotómicas para la familia Staphylinidae (NEWTON et al., 2000; NAVARRETE-HEREDIA et al., 2002), y en algunos casos se contó con la colaboración del Dr. A.F. Newton del Field Museum of Natural History.
Análisis de los datos
Para evaluar la diversidad local en cada fragmento de bosque se determinó la abundancia de estafilínidos con base en el número de individuos por grupo taxonómico, y la riqueza como el número de especies en cada localidad. Asimismo se calcularon índices de diversidad de Shannon a través del programa Krebs. La diversidad beta, se analizó comparando la similitud entre bosques, a través del índice cualitativo de Jaccard y Sorensen-cuantitativo (MORENO, 2001).
La prueba de normalidad (Shapiro-Wilk) y homogeneidad de varianza (Levene) se aplicó a las abundancias en cada bosque y para cada método. Mediante la prueba U Mann-Whitney, se compararon las abundancias de las especies en cada fragmento de bosque y para cada método de colecta, a través de Statistica Release 7. Para evaluar la efectividad de los muestreos, se realizaron curvas de acumulación de especies, con los índices no paramétricos Chao 1 y ACE, usando el programa Estímate 6.0 (COWELL, 2000).
RESULTADOSSe colectaron 801 individuos separados en 78 morfoespecies de 36 géneros pertenecientes a 8 subfamilias. Las subfamilias más representativas en cuanto a riqueza de especies fueron Aleocharinae (25,3%), Pselaphinae (17,7%), Staphylininae (15,2%), Paederinae (12,6%), Oxytelinae (10,1%), Osoriinae (8,9%), Tachyporinae (7,6%) y Euasthetinae (1,3%). En la Tabla 1, se muestra el número de morfoespecies encontradas en el Sendero Cedro Rosado y el Cañón del río Barbas según cada método de colecta.
De las 66 unidades de muestreo empleadas en ambos bosques, el 78,8% presentaron captura de estafilínidos. En el SCR se registraron 41 especies, 11 colectadas en necrotrampas, donde Styngetus sp. 2 mostró un mayor número de individuos; mientras que en hojarasca se encontraron 36 especies siendo Arthmius sp. 1, Anotylus sp. 6, Arctophysis sp., las más abundantes. En el CRB también se registró una mayor abundancia en hojarasca (31 especies) comparada con las necrotrampas (21 especies), donde Holotrocus y Anotylus fueron los géneros más abundantes respectivamente, para un registro total de 45 especies en este bosque.
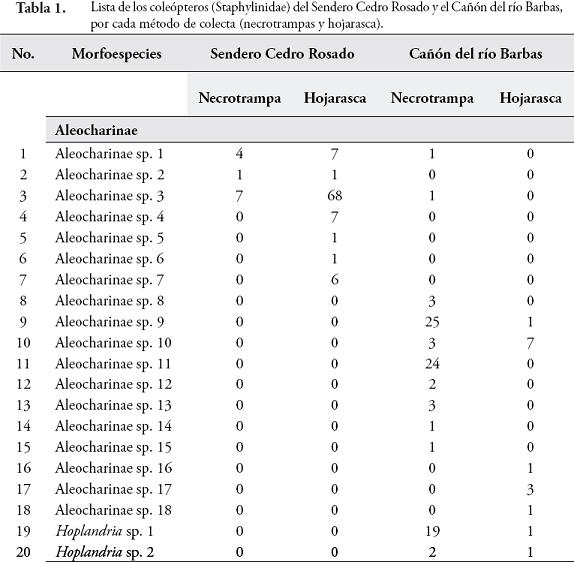
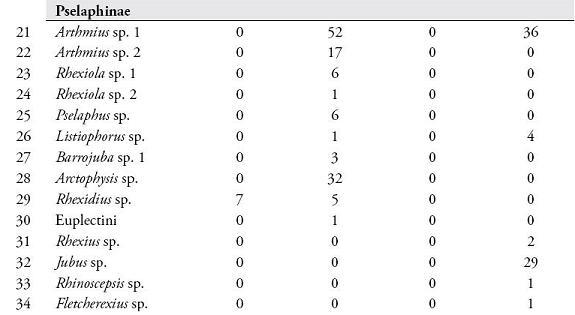
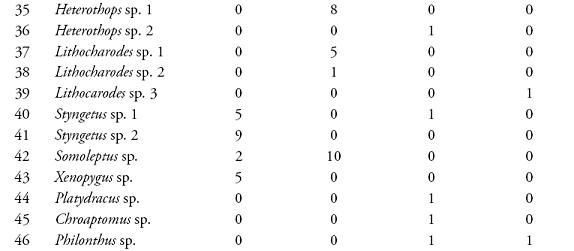
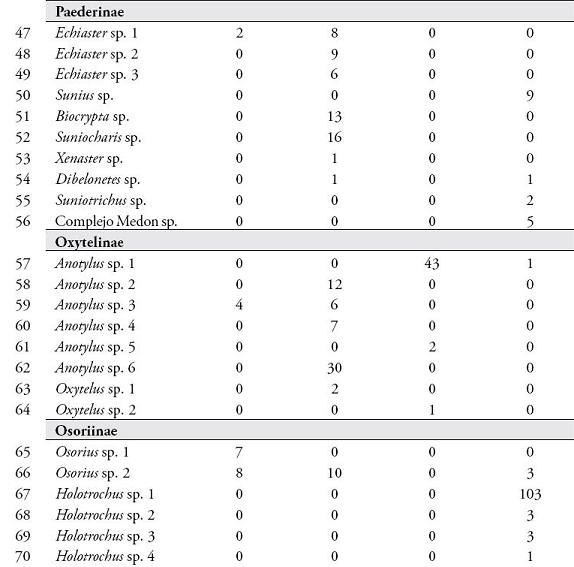
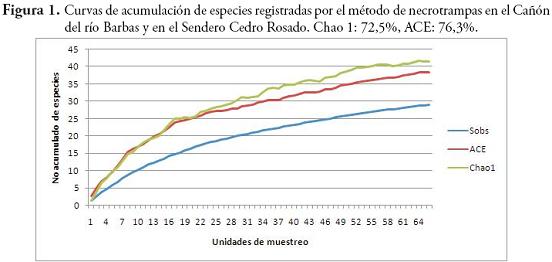
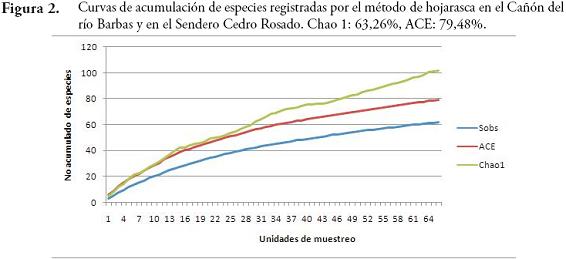
La diversidad de especies arrojada por el índice de Shannon-Wiener fue mayor en el SCR (H=4,54) que en el CRB (H=3,87). La similitud en la composición de especies de ambos bosques fue baja según los índices de Sorensen-cuantitativo (0,1073) y Jaccard (0,082). El SCR y CRB no presentaron diferencias significativas en cuanto a la riqueza (U=-1,22, p=0,21) y abundancia de especies (U=-0,835, p=0,403). Las abundancias registradas por los métodos de hojarasca y necrotrampa presentaron diferencias significativas (U=4,67, p=0,000). Esta diferenciación coincide con los resultados obtenidos al evaluar la eficacia de los métodos de colecta usados, donde se obtuvo un esfuerzo de muestreo de 72,5 % (Chao 1) y 76,3 % (ACE) para las necrotrampas (Figura 1); mientras que para hojarasca los valores de Chao 1 y ACE fueron 63,26 % y 79,48 %, respectivamente (Figura 2).
DISCUSIÓN
La representatividad por riqueza de especies encontrada para cada subfamilia, refleja su proporción dentro de Staphylinidae. De este modo, Aleocharinae, Pselaphinae, Staphylininae y Paederinae, que en orden son las subfamilias con mayor número de especies descritas mundialmente (NAVARRETE-HEREDIA et al., 2002), fueron las más dominantes en todo el muestreo; concordando con los resultados obtenidos en otros estudios (GUTIÉRREZ-CHACÓN & ULLOA-CHACÓN, 2006). El número de capturas de estafilínidos por unidad de muestreo en ambos bosques, fue alto (78,8%) comparado con un estudio de estafilínidos realizado en bosques secos en el Valle del Cauca (GARCÍA et al., 2001) donde sólo el 44% de las estaciones mostró presencia de estos insectos. Tales diferencias podrían deberse a que los estafilínidos responden a algunos factores ambientales como la temperatura del suelo y la humedad de la hojarasca (Ibíd.).
La diversidad alfa fue alta para ambos bosques, y no se encontraron diferencias estadísticamente significativas en cuanto a riqueza y abundancia entre los dos sitios de muestreo. Es posible que su diferenciación esté dada en mayor medida por una composición de especies distinta, que se refleja en los bajos valores de diversidad beta, ya que los índices Sorensen-cuantitativo y Jaccard, indican que no existe similitud en la composición de especies en cada uno de los bosques. Esto se debe a que el número de especies compartidas por ellos, no excede a 14 morfoespecies. Por lo tanto, las diferencias en la altitud y en la riqueza-abundancia de la vegetación, podrían llegar a establecer condiciones microclimáticas propicias para especies particulares en cada bosque (SARMIENTO-M, 2000). Otro factor determinante podría ser la diferencia entre las fechas de muestreo, pues durante un año pueden ocurrir cambios en las condiciones climáticas de los sitios que afecten la composición de las especies.
Al evaluar la eficacia de los métodos de colecta usados se obtuvo un esfuerzo de muestreo mayor para las necrotrampas que para la hojarasca. No obstante, debido a que los métodos se relacionan directamente con los hábitos de las especies (POWEL et al., 1996; NAVARRETE-HEREDIA et al., 2002; SARMIENTO-M, 2003), las necrotrampas permiten colectar una pequeña fracción de la comunidad, incluyendo sólo géneros saprófagos y depredadores como Hoplandria, Styngentus, Xenopygus, Chroaptomus y Philonthus que son abundantes en materia orgánica en descomposición (NAVARRETE-HEREDIA et al., 2002). Por otro lado, las morfoespecies de los géneros Holotrochus, Arthmius, Jubus, Arctophysis, Aneucamptus entre otros, fueron en este estudio géneros únicos y abundantes en hojarasca y han sido reportados como géneros propios de este microhábitat (Ibíd.). De esta manera, varios autores reportan que los métodos con necrotrampa y hojarasca son complementarios e indispensables para obtener una mayor representatividad de la fauna de estafilínidos (GARCÍA et al., 2001; GARCÍA & CHACÓN, 2005; GUTIÉRREZ-CHACÓN & ULLOA-CHACÓN, 2006).
Debido a la alta diversidad que presenta Staphylinidae (BLACKWELDER, 1936; NEWTON, 1990; KLIMASZEWSKI et al., 1996; NEWTON et al., 2000; NAVARRETE-HEREDIA et al., 2002; NEWTON et al., 2005) y a que los inventarios de grupos tan diversos como éste, raramente pueden ser completos (SACKMANN, 2006), se requiere una evaluación más amplia usando los métodos complementarios sugeridos, y teniendo en cuenta tanto la variación temporal como espacial de la distribución de las especies para construir un inventario representativo de esta comunidad (SACKMANN, 2006; JIMÉNEZ et al., 2007). Sin embargo, el trabajo realizado en el bosque del CRB y el SCR es un gran aporte para el conocimiento de la familia en el Quindío, donde hasta el momento no se habían desarrollado trabajos de este tipo. Por lo que por algunas morfoespecies listadas aquí pueden considerarse como nuevos registros para el departamento.
AGRADECIMIENTOSAgradecemos al Laboratorio de Biología y al Museo de Artrópodos de la Universidad del Quindío (MAUQ) por el préstamo de equipos; al Dr. Alfred F. Newton del Field Museum of Natural History, por su colaboración en la determinación de los especímenes; a Alba Lucía Sánchez, por facilitar alojamiento e ingreso al Cañón del río Barbas; a Jannet Molina Rico, por su asesoría y sugerencias; y a Carlos A. Soto por su colaboración en la fase de campo.
BIBLIOGRAFÍA AL DHAFER, H., 2003.- A review of specialized behaviors within the Staphylinidae. p. 1-18. Disponible en: http://www.colostate.edu/Depts/Entomology/courses/en507/papers_2003/aldhafer.pdf. [ Links ]
BLACKWELDER, R.E., 1936.-Morphology of the Coleopterous Family Staphylinidae. Smithsonian Miscellaneuos Collections, 94 (13): 1-102. [ Links ]
CASTRO, G. I., 1995.- Inventario de la flora del relicto vegetal y elaboración de folletos para visitantes al sendero de interpretación ambiental "Cedro Rosado" de la Universidad del Quindío: Tesis, Universidad del Quindío, Facultad de Ciencias Básicas y Tecnológicas, Armenia. [ Links ]
COWELL, R., 2000.- Estimates, Estatistical estimation of species richness and shared species from Simples. Version 6.01b. University of Connecticut. Disponible en: http://www.eeb.uconn.edu. [ Links ]
GARCÍA, R., ARMBRECHT, I. & ULLOA-CHACÓN, P., 2001.-Staphylinidae: Coleóptera. Composición y mirmecofilia en bosques secos relictuales de Colombia. Folia Entomológica Mexicana, 40 (1): 1-10. [ Links ]
GARCÍA, R. & CHACÓN, P., 2005.- Estafilínidos (Coleóptera: Staphylinidae) en fragmentos de bosque seco del valle geográfico del río Cauca. Revista Colombiana de Entomología, 31 (1): 43-50. [ Links ]
GUTIÉRREZ-CHACÓN, C. & ULLOA-CHACÓN, P., 2006.-Composición de estafilínidos (Coleóptera: Staphylinidae) asociados a hojarasca en la Cordillera Oriental de Colombia. Folia Entomológica Mexicana, 45 (002): 69-81. [ Links ]
JIMÉNEZ, E., LOZANO-ZAMBRANO, F. & ÁLVAREZ-SAA, G., 2007.-Diversidad alfa y beta de hormigas cazadoras del suelo en tres paisajes ganaderos de los Andes Centrales de Colombia: 439-459 (en) JIMÉNEZ, E., FERNÁNDEZ, F., ARIAS, T. & LOZANO-ZAMBRANO, F. (eds.) Sistemática, biogeografía y conservación de hormigas cazadoras de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá. [ Links ]
KLIMASZEWSKI, J., NEWTON, A. F. & THAYER, M.K., 1996.-A review of New Zealand rove beetles (Coleoptera: Staphylinidae). New Zealand Journal of Zoology, 23: 143-160. [ Links ]
MARTÍN, F., 1997.-Apuntes sobre Biodiversidad y Conservación de Insectos: Dilemas, Ficciones y ¿Soluciones? Bol. SEA, (20): 25-55. [ Links ]
MENDOZA, J.E., JIMÉNEZ, E., LOZANO-ZAMBRANO, F.H., CAICEDO-ROSALES, P. & RENJIFO, L.M., 2007.- Identificación de elementos del paisaje prioritarios para la conservación de biodiversidad en paisajes rurales en los Andes Centrales de Colombia (en) SÁENZ, J. & HARVEY, C.A. (eds.) Conservación de biodiversidad en paisajes fragmentados y agropaisajes. Heredia, Costa Rica: Editorial UNA. [ Links ]
MORENO, C.E., 2001.-Métodos para medir la biodiversidad. M&T-Manuales y Tesis SEA. [ Links ]
NAVARRETE-HEREDIA, J.L., NEWTON, A.F., THAYER, M.K., ASHE, J.S. & CHANDLER, D.S., 2002.-Guía ilustrada para los géneros de Staphylinidae (Coleóptera) de México. Universidad de Guadalajara y CONABIO, México D.F. [ Links ]
NEWTON, A.F., 1990.- Insecta: Coleoptera Staphylinidae adults and larvae: 1138-174 (en) DINDAL, D.L. (ed.) Soil Biology Guide. J Wiley and Sons. [ Links ]
NEWTON, A. F., THAYER, M. K., ASHE, J. S. & CHANDLER, D. S., 2000.- Family 22. Staphylinidae Latreille (1802): 272-418 (en) ARNETT, R. JR. & THOMAS, M. C. (eds.) American Beetles, Volume 1, Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press LLC, Boca Ratón. [ Links ]
NEWTON, A.F., GUTIÉRREZ-CHACÓN, C. & CHANDLER, D.S., 2005.- Checklist of Staphylinidae (Coleoptera) Colombia. Biota Colombiana, 6 (1): 1-72. [ Links ]
POWEL, W., WALTON, M. & JERVIS, M., 1996.- Populations and Communities:, 223-292 (en) JERVIS, M. & KIDD, N. (eds.) Insects natural enemies. Chapman & Hall. [ Links ]
SACKMANN, P., 2006.- Efectos de la variación temporal y los métodos de captura en la eficiencia de un muestreo de coleópteros en la Reserva Natural Loma del Medio, El Bolsón, río Negro. Rev. Soc. Entomol. Argent., 65 (3-4): 35-50. [ Links ]
SARMIENTO-M, C.E., 2000.- Comparación de tres clases de transectos para la captura de hormigas en dos formaciones vegetales. Caldasia, 22 (2): 317-326. [ Links ]
________________., 2003.-Metodología de captura y estudio de hormigas: 201-210 (en) FERNÁNDEZ, F. (ed.) Introducción a las Hormigas de la región Neotropical. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá. [ Links ]
SKALSKI, T. & PÓSPIECH, N., 2006.- Beetles community structures under different reclamation practices. European Journal of Soil Biology, 42: 316-S320. [ Links ]

