










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.14 no.1 Manizales Jan./June 2010
MICROSCOPY OF PEDOCOMPONENTS AT AN ARCHAEOLOGICAL SITE IN WESTERN ANTIOQUIA. EMPHASIS IN ARCHAEOBOTANY AND PALEOECOLOGY.
* Recibido febrero 17 de 2010, aceptado Junio 29 de 2010.
1 Antropólogo, M.Sc. Profesor Universidad de Caldas. E-mail: willyposada@yahoo.es
2 Ingeniero Geólogo, Ph.D. Profesor Universidad Nacional de Colombia, sede Medellín. E-mail: lnparra@unalmed.edu.co
Se presentan los resultados de un análisis de componentes microscópicos del suelo en un contexto arqueológico, tratando de obtener diversas líneas de evidencia para entender las relaciones entre las poblaciones humanas y su medio ambiente natural. Tras la reconstrucción paleoecológica se concluyó que en estos contextos existen múltiples indicadores de las condiciones de vegetación, clima y actividad humana, distintos de los tradicionales marcadores palinológicos; entre ellos, los tejidos lignolíticos, los hongos y los fitolitos. Todos estos resultados fueron coherentes con los procesos de transformación del bosque en la zona, y plantean nuevos horizontes de investigación en ecología vegetal y etnobotánica.
Palabras clave: arqueología, paleobotánica, paleoecología, pedocomponentes, fitolitos, suelos.
AbstractThis article presents the results of a microscopic analysis of soil components in an archaeological context, in order to obtain several lines of evidence for understanding of the relationships between human populations and their natural environment. After the paleoecological reconstruction, it was concluded that in these contexts there are multiple indicators of vegetation, climate and human activity conditions, different from the traditional palynological markers, including ligninolitic tissues, fungi and phytoliths. All these results were consistent with the conversion process of forest in the area. They also present new horizons for research in plant ecology and ethnobotany.
Key words: archaeology, paleobotany, paleoecology, pedocomponents, phytoliths, soils.
El estudio de pedocomponentes es una propuesta de análisis paleoecológico que surge a partir de los avances en petrografía de la materia orgánica, mineralogía óptica, palinología y micromorfología de suelos. El análisis consiste en la identificación y cuantificación de asociaciones de componentes microscópicos orgánicos, inorgánicos o biominerales presentes en el suelo, los cuales pueden ser empleados en la interpretación de condiciones ambientales específicas. Su importancia radica en que los componentes en conjunto representan un registro integral de los diversos agentes y procesos climáticos, geológicos, edáficos y biológicos, que intervienen en el contexto de depositación y evolución del suelo. En Colombia, solamente en el ámbito de procesos de génesis de Andisoles, FLÓREZ (2000) ha desarrollado métodos de análisis de pedocomponentes para identificar la presencia de ciertos procesos de formación de suelo partiendo del reconocimiento de atributos morfológicos y fisicoquímicos en los componentes. Igualmente, PARRA-SÁNCHEZ (2005) ha empleado el mismo espectro de componentes para la identificación de variaciones climáticas y eventos volcánicos aunque en sistemas lénticos como lagunas y humedales. Pese a que en la arqueología de Colombia los estudios de polen vienen realizándose desde hace ya varias décadas, pocos trabajos en el país han complementado los resultados de tales estudios con otras líneas de evidencia, así como tampoco se ha avanzado en la búsqueda de nuevos indicadores de actividad antrópica que soporten el paso de los años y las condiciones extremas del trópico que caracterizan los suelos de los sitios arqueológicos del país (CAVELIER et al.,1995; GNECCO, 2000; MONTEJO & RODRÍGUEZ, 2001; ACEITUNO, 2002; ARCHILA, 2005). Los pedocomponentes de origen antrópico son entonces una fuente de evidencia sólida de las actividades humanas del pasado, sobre todo cuando faltan los artefactos y otros restos macroscópicos por diversas causas como el barrido de las basuras hacia depósitos secundarios, el uso diferencial de los sitios o el efecto de procesos posdeposicionales de diversa índole (SCHIFFER, 1987; MILLER-ROSEN, 1989).
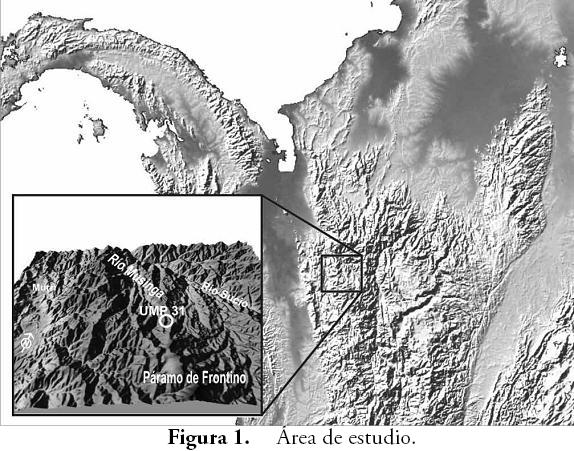
En este contexto, una investigación arqueológica llevada a cabo en el municipio de Frontino en Antioquia (PIAZZINI et al., 2009), identificó una concentración de aterrazamientos artificiales de origen antrópico localizada en la cuenca media del río Musinga, todas ellas con evidencia de material lítico y cerámico de edad variable entre los primeros ocho siglos de nuestra era. Uno de estos aterrazamientos es el sitio arqueológico UMP 31, sitio en el cual se recuperaron artefactos líticos asociados con la explotación del bosque, macrorrestos vegetales y otros rasgos estratigráficos que sugieren un proceso de alteración de los ecosistemas a través de prácticas sociales concretas. Identificar cuáles fueron estas prácticas así como el contexto natural en donde se desarrollaron, se convierte en una cuestión a resolver mediante un análisis de pedocomponentes, toda vez que las circunstancias ecológicas fueron parte fundamental de estos procesos de adaptación, transformación y domesticación social del medio ambiente natural.
Área de estudioEl sitio arqueológico UMP 31 se localiza en la cuenca media-alta del río Musinga, vereda Musinguita, municipio de Frontino en el occidente de Antioquia, Colombia (Figura 1). La zona comprende cuchillas y colinas de fuertes pendientes, desarrolladas sobre rocas de la Formación Penderisco tales como lutitas, areniscas y chert interestratificadas y plegadas, a menudo suprayacidas por depósitos de vertiente, mientras que las partes planas de los valles están ocupadas por sedimentos de origen aluvial y de edad Cuaternaria.
El clima del sector varía principalmente en función de la altitud. La temperatura oscila entre los 18 y 24 ºC. Existen dos períodos de verano y dos de invierno, siendo el último invierno en octubre más intenso y prolongado, así como más húmedo y seco (ALZATE, 1995). Las isoyetas son del orden de los 2000 y 2500 mm anuales aumentando continuamente hacia el occidente. Por estas características, las zonas de vida predominantes en el área de estudio son el bosque pluvial premontano (bp-PM) y el bosque muy húmedo premontano (bmh-PM).
METODOLOGÍA
El levantamiento estratigráfico del sitio (Figura 2) logró identificar una pedoestratigrafía compleja, cuyos patrones morfológicos fueron discontinuos y en algunos sectores del sitio ni siquiera se observaban con claridad por efecto de alteraciones posdeposicionales y otros fenómenos pedogenéticos. Esta situación llevó a que se recuperaran varios núcleos en el sitio, tratando de obtener muestras de aquellos horizontes en donde las ocupaciones estuvieran mejor representadas. Adicionalmente, se consideró el contexto geológico a partir del reconocimiento de la estratigrafía natural de toda la colina.
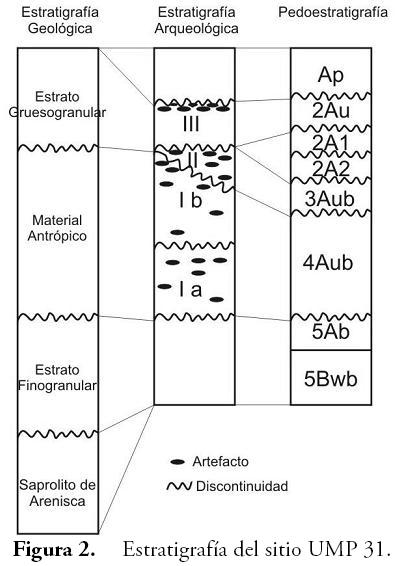
Las muestras de suelo para el análisis se tomaron sin disturbar de cuatro sectores distintos del aterrazamiento, previa lectura de los perfiles. Inicialmente se adoptó el método de extracción por cilindro, siguiendo la metodología empleada por FLÓREZ (2000); sin embargo, se identificaron problemas relacionados con la resistencia del suelo a la penetración por efecto de las gravas y los artefactos, por lo cual la recuperación de los núcleos siguientes se hizo mediante canaletas de penetración en el perfil. Como resultado, se obtuvo una columna de 105 cm de perfil de suelo repartidos en tres cilindros (núcleo 1), los cuales se marcaron con el nombre del sitio y las profundidades, se sellaron con papel plástico de envoltura y se cubrieron con bolsas plásticas para evitar la pérdida de humedad. Procedimiento similar se hizo con las canaletas de aluminio que se introdujeron lateralmente (núcleos 2, 3a, 3b y 4).
Una vez abiertas, las columnas de suelo de cada núcleo fueron descritas y se procedió a extraer una muestra de 2 cm³ por cada horizonte. Sólo en el horizonte 4Aub se extrajeron 3 muestras correspondientes a la base, el centro y el techo del mismo, pues era uno de los horizontes de mayor espesor e interés en todo el perfil ya que albergaba los principales estratos de ocupación.
El tratamiento de las muestras a pesar de seguir con algunas modificaciones la metodología descrita por FLÓREZ (2000), involucró una serie de experimentos con el fin de obtener el mejor resultado, siempre buscando la máxima dispersión del suelo y la mínima destrucción de pedocomponentes, de cuyo resultado se optó por saturar la muestra de suelo en una solución de isopropanol amoníaco 1:2 y dispersarlo mecánicamente mediante licuadora a 5 r.p.m. durante 2 minutos.
Cada muestra fue pasada por un juego de mallas -140 +635, el cual retendría de manera separada dos fracciones de material: una para el sedimento de más de 106 micras (-140) que serviría para un análisis menor y complementario de los detritos orgánicos y eventuales microartefactos, y otra para el sedimento entre 106 y 20 micras (+635), que sería el objeto final de observación al microscopio.
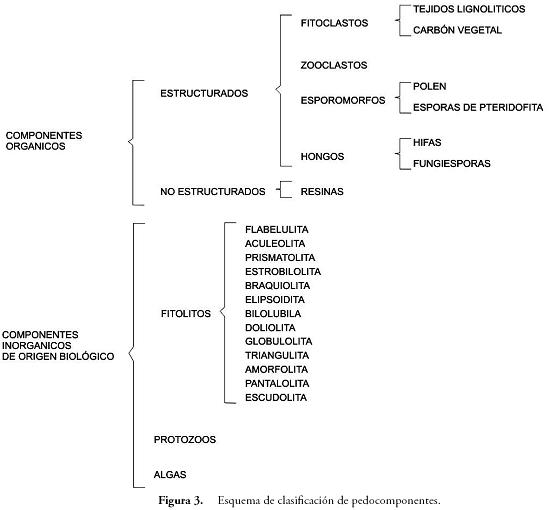
Por último, se realizó el montaje en Naphrax™ y se procedió a la lectura de placas en el microscopio petrográfico de luz transmitida, contando un total de 300 componentes bajo el objetivo de 40X.El registro de los pedocomponentes durante los conteos se hizo considerando aspectos clave de su morfología, y otros caracteres específicos según criterios formulados por varios autores (PIPERNO, 1988; TRAVERSE, 1994; TYSON, 1995; FLÓREZ, 2000; PARRA & FLÓREZ, 2001; MADELLA et al., 2005; PARRA, 2005; ERCEGOBAC & KOSTIC, 2006; FLÓREZ et al., 2006; SEBAG et al., 2006). Dicho registro consideró solamente los pedocomponentes de origen biológico tal como se muestra en la clasificación de la Figura 3, debido a que, en este caso, eran los que ofrecían mayor potencial de registro de procesos y eventos de interés arqueológico.
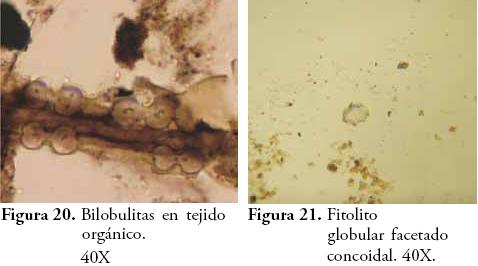
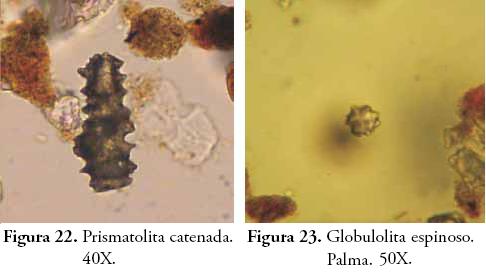
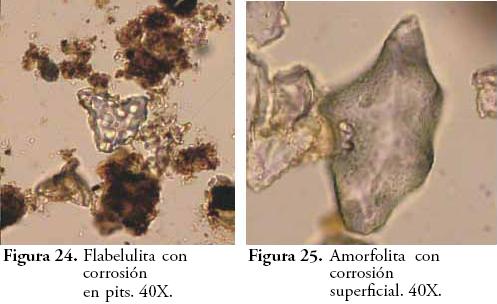
Para el caso de los fitolitos se empleó la taxonomía propuesta por PARRA & FLÓREZ (2001) y se registraron características tales como: ornamentación (verrugoso, espinoso, psilado, catenado y alado), corrosión (sin corrosión, corrosión superficial, corrosión en pits u hoyos y cavidades o cráteres), anillos de hidratación (presencia o ausencia), coloración (presencia o ausencia) y articulación (presencia o ausencia). En todos los horizontes se corrió el índice D:P (fitolitos de dicotiledóneas vs. fitolitos de Poaceae) empleado por otros autores para evaluar la variación relativa de estos dos grandes grupos de vegetación, tanto en el contexto de cambios climáticos como por efecto de la manipulación o perturbación antrópica de plantas (ALEXANDRE et al., 1997b; ALEXANDRE et al., 1999; BREMOND et al., 2005). Los diagramas de frecuencias se elaboraron en el software C2 Data Analysis versión 1.5.1. y otras estadísticas descriptivas en Microsoft Excell 2007.
DESCRIPCIÓN DE RESULTADOS
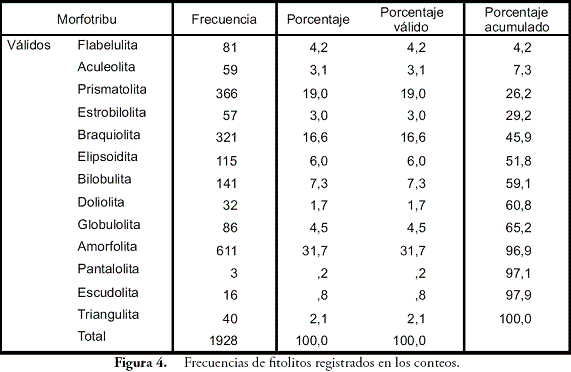
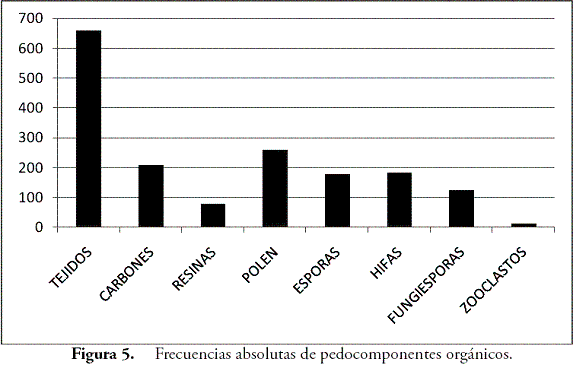
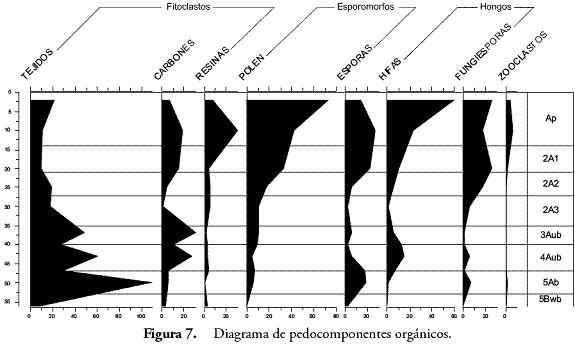
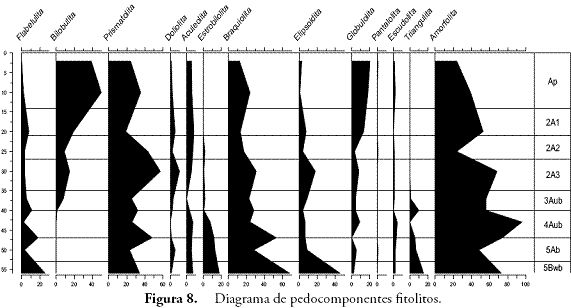
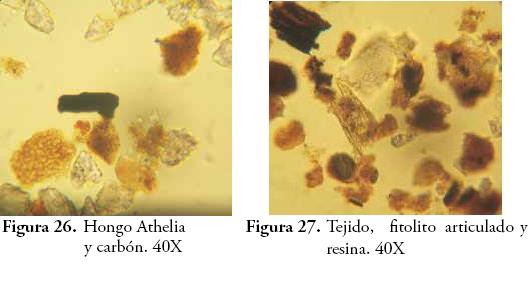
El análisis de pedocomponentes se hizo a partir de un conteo de 1.933 componentes inorgánicos de origen biológico y 2.265 componentes orgánicos. En el primer caso, los fitolitos ocuparon el 99% de los casos, estando el 1% restante representado en algas como las diatomeas y las crisofitas. Por esta razón, se pondrá cierto énfasis en los fitolitos. En el segundo caso, tejidos, esporomorfos y hongos dominaron en ese orden las frecuencias de los conteos. En el caso de los fitolitos, las formas asimétricas o irregulares registradas por la morfotribu Amorfolita ocuparon gran parte de los porcentajes (Figura 4). Los componentes orgánicos estuvieron dominados por los tejidos vegetales de naturaleza lignocelulosa, opacos, estructurados y perforados, con un total de 659 partículas distribuidas principalmente hacia los horizontes inferiores, seguidos por el polen con 260 y el carbón con 210 partículas (Figura 5).
INTERPRETACIÓN DE RESULTADOS Y PALEORECONSTRUCCIÓN
A continuación se hará la interpretación diacrónica de los pedocomponentes comenzando de lo más antiguo hasta lo más reciente, esto es, del horizonte más profundo 5Bwb al más superficial Ap.
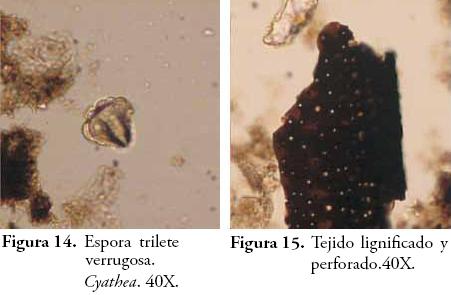
Los últimos dos horizontes muestran claramente la existencia de un suelo estable bajo cobertura boscosa. El cociente D:P ha arrojado valores de 2,0 y 2,3 que son altos y sugieren la presencia mayoritaria de plantas dicotiledóneas, tal como se observa con las formas esféricas, polihédricas y amorfas pero de contorno regular y definido que constituyen la reserva fitolítica de estas plantas según varios autores (PIPERNO, 1988; BOZARTH, 1992; ALEXANDRE et al., 1997a; ALEXANDRE et al., 1997b; PREBBLE et al., 2002; BREMOND et al., 2005; ALBERT, 2006; TSARTSIDOU, 2007). El ambiente arbóreo que se advierte por el registro de fitolitos es confirmado por la gran cantidad de tejidos lignificados o maderas pertenecientes a especies de bosque (Figura 9) (BOULDER, 1994; GASTALDO, 1994; TYSON, 1995; ERCEGOBAC & KOSTIC, 2006; SEBAG et al., 2006).
Muchos de estos tejidos han sido identificados como pertenecientes a helechos arbóreos del género Cyathea, gracias a una colección de referencia de fitolitos y tejidos perennes obtenida de algunas plantas de la zona (POSADA et al., 2009) y a su correlación con las esporas trilete de ornamentación verrugosa halladas en este horizonte. Estos helechos conocidos por los lugareños de la región como zarros, abundan en los escasos bosques de la región y son indicativos de condiciones climáticas frías y húmedas. Las poblaciones Embera actuales de la región emplean el tronco de estos helechos para la fabricación de sus viviendas, pues consideran que es una madera dura muy propicia para los cimientos de la estructura palafítica de los tambos. Otros tejidos silíceos ocluídos y lignificados diagnósticos de Asteraceae (PIPERNO, 1988; BOZARTH, 1992) fueron hallados también en este contexto. La humedad sugerida por Cyathea que a su vez es una característica del ambiente edáfico de los bosques, es confirmada por el aumento de hongos representados en la densidad de fungiesporas.
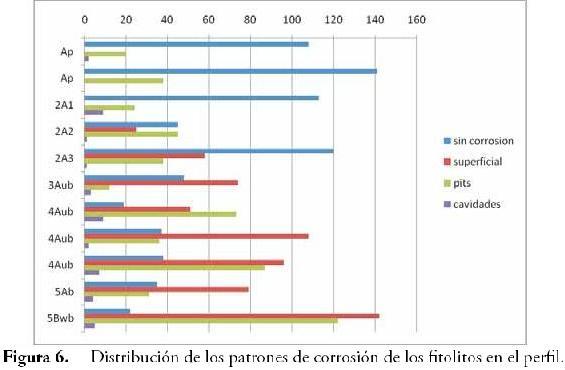
Dado que este es el horizonte más antiguo del perfil (suelo enterrado), los fitolitos presentan un mayor grado de corrosión. De acuerdo con la Figura 6, el grado de corrosión de los fitolitos se generaliza a medida que avanza la profundidad, mostrando hacia los niveles más superficiales una mayor cantidad de fitolitos en buen estado de conservación. Según FLÓREZ & PARRA (1999) y FLÓREZ et al. (2006) la corrosión de los fitolitos en paleosuelos ándicos está fuertemente relacionada con la materia orgánica. Sin embargo, aun sin descartar esa posibilidad, es claro que horizontes tales como el 5Bwb cuyo contenido orgánico es prácticamente nulo, posee una mayor densidad de fitolitos corroídos comparada con la del horizonte Ap u otros horizontes humificados. En este sentido y pese a la translocación de fitolitos en el suelo, podría plantearse que la corrosión en este caso está más influenciada por el tiempo que llevan los fitolitos en el ambiente edáfico que por los procesos pedogenéticos en sí mismos, aun cuando estos actúan de manera diferencial en los fitolitos generando distintos patrones de corrosión. Este aspecto, observado también por ALEXANDRE et al., (1999), manifiesta a su vez el valor cronológico o estratigráfico que subyace a la corrosión de los fitolitos, sobretodo en patrones de corrosión en pits (hoyos) y de corrosión superficial, pues los cráteres o corrosión en cavidades no mostraron ser sensibles a los cambios en la profundidad y por consiguiente anulan la posibilidad de servir como marcadores estratigráficos. Resta identificar qué pautas o rasgos pedogenéticos determinan la variación en uno y otro contexto de las causas de la corrosión (i.e. FLÓREZ & PARRA, 1999; FLÓREZ et al., 2006).

Se observa entonces cómo la llegada de los primeros grupos humanos al lugar estuvo acompañada de cambios significativos en el ecosistema circundante: la apertura del bosque mediante la tumba y quema fue motivo del cambio súbito en la vegetación y en las propiedades del suelo. Cuando un claro aparece en el bosque una nueva sucesión vegetal toma lugar, las plantas pioneras aparecen acelerando sus ciclos reproductivos y produciendo abundantes flores, frutos y semillas que resultan atractivos para animales e insectos (COLINVAUX & BUSH, 1991; GUREVITCH et al., 2006). Esta situación, tiene además implicaciones en las propiedades bioorgánicas del suelo, en nuestro caso, el incremento de la actividad biológica inferida de la cantidad de hifas y esporas de hongos tras el banqueo del área desmontada. Dicha actividad, seguramente fue enriquecida también por el aporte de residuos orgánicos al suelo como producto de sus actividades cotidianas.
En el horizonte antrópico 4Aub2 disminuyen los fitolitos de dicotiledóneas y tejidos de plantas arbóreas respecto al horizonte anterior, tal como se observa en el diagrama y en el cociente D:P. No obstante, la presencia de árboles o maderas sigue siendo predominante junto con los carbones e hifas de hongos. Estos componentes son evidencia de posibles actividades de tumba y quema de los elementos del bosque para la apertura de claros para vivienda y/o cultivo. Es una de las prácticas más comunes entre indígenas y colonos de los bosques tropicales y se ha documentado ampliamente en la literatura antropológica y arqueológica en América (HERRERA, 1985; DESCOLA, 1987; BUSH & COLINVAUX, 1994; RANERE & COOK, 1995; PIPERNO & PEARSALL, 1998; CABRERA et. al., 1999; PIPERNO, 2007). El aumento exagerado de Amorfolitas en este horizonte puede explicarse como un resultado del aumento de biomasa vegetal arbórea en el suelo tras las actividades de desmonte. Pese a que no hay muchos tejidos, esta hipótesis también explicaría la diversidad de Amorfolitas, de hongos y de tejidos epidérmicos silicificados (fitolitos articulados) y tricomas intactos. La abundancia de hongos en el suelo –además de la humedad– está fuertemente relacionada con la presencia de materia orgánica en descomposición, ya que el componente fungi del suelo es mayoritariamente saprófito acelerando su crecimiento y reproducción con la adición de hojarasca y la consecuente liberación de nutrientes como C y N al suelo (PAUSTIAN & SCHNURER, 1987; NADELHOFFER et al., 2001. Los fitolitos globulares espinosos considerados diagnósticos de Palma (Arecaeae) (PIPERNO, 1988; MILLER-ROSEN, 1992; SCOTT, 1992; ALEXANDRE et al., 1997b), fueron hallados también en este horizonte. Aun cuando se desconoce la taxa específica de origen, su presencia está relacionada con su uso alimenticio o como insumo para la fabricación de artefactos o estructuras de vivienda durante esta primera etapa de la ocupación. Las palmas han sido descritas en las crónicas como parte fundamental de la reserva vegetal de uso cotidiano entre los aborígenes de estas tierras en el siglo XVI (CIEZA DE LEÓN, 1984: 32), tal como lo han demostrado otros autores en diversos contextos arqueológicos en América (CAVELIER et al., 1995; PIPERNO & PEARSALL, 1998; MORCOTE & BERNAL, 2001).
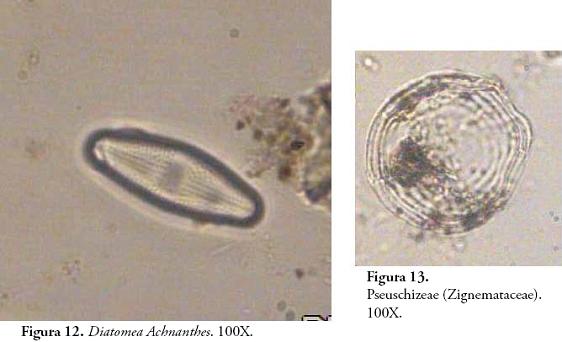
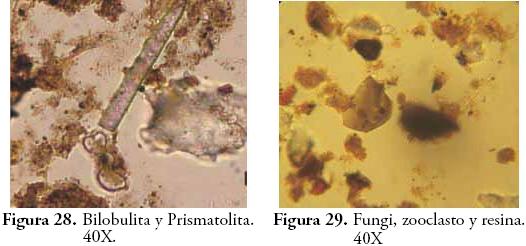
Posteriormente, hacia el horizonte 3Aub a pesar de la predominancia de Amorfolita, descienden las taxa arbóreas representadas en Triangulita, Globulolita y Aculeolita y comienzan su ascenso las Gramíneas con Bilobulita, Prismatolita y Doliolita en aumento. Este cambio es coherente con la sucesión vegetal que ha de ocurrir luego de la apertura del bosque por parte de los seres humanos mediante la tumba y quema del bosque. La relativa constancia de tejidos silíceos ocluídos (platelets), es evidencia de dicotiledóneas pioneras como Asteraceae (BOZARTH, 1992). Pteridofitos como Lycopodium que crece a orillas de caminos o en zonas abiertas, es otro indicador de la dinámica de sucesión en el contexto de perturbación antrópica y es frecuente en el registro palinológico de este horizonte. Si bien existen diatomeas en las películas de agua del suelo, la presencia de Achnanthes sp. no deja de ser inquietante ya que son algas que por lo general requieren condiciones de saturación de agua y que según DÍAZ & RIVERA (2004: 391), son más frecuentes ante valores elevados de fósforo disuelto. Por consiguiente, su ocurrencia en este horizonte puede estar relacionada con procesos de eutrofización de la solución del suelo con aportes orgánicos o con el transporte de dichas algas desde algún cuerpo de agua permanente rico en P soluble. Esta posibilidad abre un interesante panorama de estudio de microfósiles limnológicos en relación con la organización de actividades y tareas domésticas (captación de recursos hídricos, almacenamiento de agua, adición de nutrientes al suelo, preparación de alimentos, etc.).
Hacia 2A2 la actividad antrópica en el sitio se ve interrumpida por un proceso de depositación de sedimentos gruesos. Aunque el cociente D:P sigue disminuyendo y está por debajo de 1,0 mostrando la predominancia de vegetación abierta, el aumento de morfotipos es generalizado incluyendo también plantas dicotiledóneas arbóreas. Los componentes orgánicos en cambio, disminuyen de manera general ofreciendo un panorama contrastado de pedocomponentes. Estos patrones lejos de indicar una nueva variación en la composición vegetal del lugar, son producto de la depositación de fitolitos alóctonos por la acción de una corriente hidráulica muy fuerte que se encargó además de lavar buena parte del material particulado fino, tal como lo son los componentes orgánicos sedimentarios. Esta deducción se hace luego de un análisis estratigráfico previo en el sitio (POSADA et al., 2009, en prensa).
Seguidamente, el horizonte 2A1 es otro testigo más del ascenso gradual de pedocomponentes, incluyendo los de la fracción orgánica, aun siendo parte de un proceso de transporte y sedimentación similar al del horizonte 2A2 pero evidentemente de menor energía. Aunque no es clara la dominancia de una determinada asociación vegetal, el cociente D:P continúa sugiriendo la presencia dominante de vegetación abierta.Hacia el horizonte 2Au se nota un aumento fuerte de minerales en detrimento de los pedocomponentes orgánicos y microfósiles. La irregular morfología de la mayoría de los fitolitos encontrados aquí (Amorfolita), podría estar relacionada con la fracturación y el redondeamiento de los mismos por transporte fluvial, tal como lo sugieren los sedimentos (POSADA et al., 2009, en prensa). Las gramíneas y los elementos pioneros continúan aumentando, pero resulta particularmente interesante un fitolito de forma globular y de ornamentación facetada-concoidal similar a los morfotipos diagnósticos de Cucurbitáceae y Annonaceae, conocidas por ser cultivadas o explotadas con frecuencia en tiempos prehispánicos. No obstante, el fitolito encontrado posee diferencias estructurales que no permiten asimilarlo completamente con las formas reportadas en Cucurbita sp., aunque podría pertenecer a algún otro género como Peponopsis o Sicana cuya ornamentación es también similar (BOZARTH, 1987; PIPERNO, 1988; PIPERNO & PEARSALL, 1998; PIPERNO et al., 2000; PIPERNO et al., 2002; PIPERNO, 2009). Vale mencionar que se ha reportado polen de Cucurbitaceae en un sitio arqueológico cercano a UMP 31 dentro de la cuenca de Musinga y asociado a un estrato del mismo período (PIAZZINI et al., 2009), lo cual podría apoyar la asociación de esta planta con el fitolito mencionado.
El contenido de pedocomponentes del horizonte Ap es ciertamente representativo de las condiciones ambientales actuales, por lo que se constituye en el principal referente del perfil para efectos de comparar las variaciones ecológicas en perspectiva diacrónica. La vegetación que actualmente soporta este horizonte está compuesta por pastos del género Axonopus (imperial y micay) y eventualmente Bambusa guadua H. et B. (Guadua), plantas que explican la densidad de Bilobulitas y Prismatolitas que caracterizan el espectro fitolítico de este horizonte. Sin embargo, la presencia de fitolitos globulares no se ajusta a la fisonomía actual de la vegetación, pues su morfología ha sido asociada con plantas dicotiledóneas generalmente de tipo arbóreo (BOZARTH, 1992; ALEXANDRE et al., 1997a; ALEXANDRE et al., 1997b; PREBBLE et al., 2002; TSARTSIDOU et al., 2007) abriendo la posibilidad de la presencia de maderas en el sitio como desecho de actividades humanas recientes (antiguas construcciones, polines de establo, saladeros para ganado, etc.). Además, se ha registrado una vez más un fitolito globular de ornamentación facetada y concoidal, esta vez sin una explicación clara.
Aunque la anterior interpretación se ha hecho como una reconstrucción intrasitio dada la escala espaciotemporal del estudio, es claro cómo los resultados en el contexto paleoambiental se ajustan a la información disponible en otros escenarios naturales. La fecha obtenida del horizonte 4Au2 de 1780±70 años A.P. (PIAZZINI et al., 2009), permite establecer una correlación positiva con los estudios paleoecológicos realizados en el Páramo de Frontino. Como ya se ha dicho, la vegetación arbórea y la gran cantidad de tejidos lignolíticos de Cyathea en este horizonte, permiten plantear la predominancia de condiciones húmedas y frías hacia el siglo segundo de nuestra era. Así mismo, VELÁSQUEZ (2005) ha registrado una fase húmeda y fría entre 1.900-1.200 años B.P. en el registro polínico del núcleo Puente Largo II de Frontino, al igual que MONSALVE (2004), quien encontró un momento frío y húmedo en sedimentos datados entre 2.063-1.656 años B.P. en el núcleo Puente Largo I. Pese a la escasez de información sistemática, datada y de alta resolución para este período cronológico, PLAZAS et al. (1993: 127) han reportado una época húmeda representada en el alto nivel del río San Jorge hacia los primeros siglos de nuestra era, la cual podría estar relacionada con un período húmedo descrito por VAN DER HAMMEN (1992: 116) en el río Amazonas hace cerca de 2.000 años A.P. Todo esto indicaría junto con los datos anteriores la ocurrencia de un fenómeno climático de gran escala que los pedocomponentes han podido registrar. Esta época además, coincide con el desarrollo de grandes sociedades en Colombia caracterizadas por una organización sociopolítica compleja, en donde la intensificación y diversificación de las prácticas productivas que caracteriza tales sociedades pudo haberse favorecido por las condiciones de humedad del clima (PIÑEROS, 1989; VAN DER HAMMEN, 1962 en LANGEBAEK, 1992: 47; REICHEL-DOLMATOFF, 1997: 103). En Frontino durante esta época se consolidan centros aldeanos como los registrados en la vereda Musinguita donde se encuentra UMP 31, cuyas evidencias paleobotánicas y artefactuales sugieren el desarrollo de prácticas agrícolas concretas que fueron importantes modos de producción para enfrentar el aumento demográfico y mantener el control de la población durante aquel período.
CONSIDERACIONES FINALESEn UMP 31 la variedad microfósil permitió reconstruir las transformaciones que sufrieron el suelo y la vegetación tras la ocupación humana. Estos resultados plantean entonces nuevos proxys para la investigación arqueológica en la medida en que amplían el panorama de indicadores paleoecológicos.
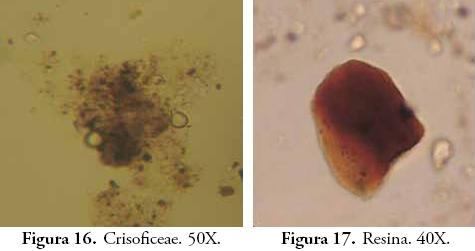
Los fitolitos y la materia orgánica sedimentaria han revelado la presencia de plantas útiles y comestibles en los períodos precolombinos de UMP 31, pese a que su frecuencia de aparición es baja en el registro. Lejos de fetichizar estas evidencias, su baja representatividad podría indicar un bajo consumo de estas plantas o su uso esporádico en aspectos distintos a la alimentación. Infortunadamente, el énfasis que se le ha puesto a la reconstrucción de paleodietas en la arqueología colombiana ha invisibilizado el valor farmacológico, artesanal o etnobotánico de gran parte de las plantas identificadas como cultivadas o domesticadas en períodos prehispánicos, aun cuando estas han sido reconocidas por su importancia medicinal o como importante materia prima para la fabricación de artefactos o estructuras de diversa índole. Las palmas por ejemplo, además de su consumo alimenticio, están entre las plantas de mayor uso doméstico y especializado en aspectos distintos a los estrictamente nutricionales (DESCOLA, 1987; CAVELIER et al., 1995; RODRÍGUEZ, 1995; POLITIS, 1996; CABRERA et al., 1999). Esta consideración puede encontrar eco a propósito de la propuesta multiproxy que plantea el estudio de pedocomponentes, baste con mencionar algunas de ellas: si bien las resinas en este trabajo no obtuvieron considerable significado, su producción mayoritaria por coníferas, Moraceaes, Fabaceas y otras especies arbóreas de valor combustible (SCHERY, 1956; GASTALDO, 1994) las augura como útiles microfósiles para la investigación de áreas de actividad específica. Los zooclastos (pelos, mandíbulas y corazas quitinosas) que en este caso sólo fueron indicativos de la actividad biológica intensa de algunos horizontes, pueden ser evidencia indirecta de la presencia de cultivos o de silos de almacenamiento, considerando las relaciones de parasitismo, comensalismo y coevolución que existen entre algunas especies de insectos y determinadas plantas (THOMPSON, 1994; FONTURBEL & MOLINA, 2002; ZAMORA, 2005). También ofrecen un interesante panorama para la reconstrucción paleoclimática y para los estudios de entomofagia en sociedades precolombinas (VELÁSQUEZ, 1993; CABRERA et al., 1999; ABRIL, 2007). Diatomeas, Tecamebas, Crisoficeae, Pseudoschizeae y otros componentes de cuerpos de agua permanente, son buenos indicadores de condiciones de saturación de agua o excesiva humedad; RICH & PIRKLE, 1994; TRAVERSE, 1994; SALGADO & STEMPER, 1995). Estas algas y protozoos también abren la posibilidad de indagar sobre las fuentes de recursos hídricos, sobre todo en aquellos casos donde el estudio del agua puede ser crítico como en los sitios de explotación precolombina de aguasal. Los tejidos lignolíticos aunque sólo pueden hablarnos de la existencia de maderas, su estudio complementado con el de fitolitos puede ser concluyente en términos etnobotánicos y de actividades artesanales.
A propósito del estudio de fitolitos, su desarrollo ha ocurrido de manera exponencial en los últimos 20 años, tiempo en el cual se han mitigado fuertemente los efectos de la multiplicidad y la redundancia en la sistemática e interpretación de estos microfósiles (RAPP & MULHOLLAND, 1992; MADELLA et al., 2005; ALBERT & MADELLA, 2008; PIPERNO, 2009). Sus ventajas en el contexto de la investigación arqueológica son múltiples, como por ejemplo su resistencia y conservación en ambientes ácidos y oxidativos, su representatividad netamente local o incluso puntual y específica, la resolución taxonómica que en algunas plantas puede alcanzar a informar incluso sobre el órgano específico de procedencia de la planta, la información de interés edáfico o estratigráfico que proviene de su superficie, etc. Todas estas bondades se han documentado en la literatura sobre fitolitos de la última década y en el caso de UMP 31, su presencia, densidad y características fueron decisivas para interpretar la alteración en los patrones de vegetación y de las propiedades bioorgánicas del suelo, así como también el efecto del tiempo en la morfología de los fitolitos.
Con este panorama, la investigación arqueológica puede valerse de más argumentos para la consolidación de hipótesis de trabajo, ampliando así no sólo las posibilidades técnicas sino interpretativas de la disciplina. Simultáneamente, plantea otra fórmula para comprender la relación entre los factores sociales y medioambientales en los procesos de configuración territorial, cambio social y cambio ambiental. Dicha comprensión exige la adopción de nuevas categorías metodológicas y conceptuales acordes con los fenómenos descubiertos, las cuales servirán de base no sólo a la discusión teórica entre biólogos, pedólogos, geólogos y arqueólogos, sino a la reanimación de espacios naturales y prácticas sociales antiguas que puedan ofrecernos hoy nuevas alternativas de manejo y apropiación de los ecosistemas de los cuales hacemos parte.
PEDOCOMPONENTES DEL SITIO ARQUEOLÓGICO UMP 31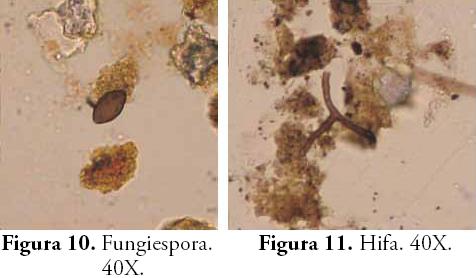

AGRADECIMIENTOS
A Diana Isabel Vergara Gómez, María Teresa Flórez Molina y César Augusto Velásquez por las valiosas contribuciones hechas a este trabajo.
ABRIL, G., 2007.- Macroinvertebrados en dos cuerpos de agua del Páramo de Frontino (Antioquia), Colombia, con énfasis en los Quironómidos: Tesis de Maestría, Universidad Nacional de Colombia, Medellín. [ Links ]
ACEITUNO, F.J., 2002.- Interacciones fitoculturales en el cauca medio durante el Holoceno Temprano y Medio. Arqueología del Área Intermedia, 4: 89-113. [ Links ]
ALBERT, R.M., 2006.- Reconstrucción de la vegetación en África oriental durante el Plio-Pleistoceno a través del estudio de fitolitos [La garganta de Olduvai (Tanzania)]. Ecosistemas, 15 (1): 47-58. [ Links ]
ALBERT, R.M. & MADELLA, M., 2008.- Perspectives on phytolith research: 6th International meeting on phytolith research. Quaternary International doi:10.1016/j.quaint.2008.07.002. [ Links ]
ALEXANDRE, A.; MEUNIER, J.D.; FABRICE, C. & JEAN, M.K., 1997a.- Plant impact in the biogechemical cycle of silicon and related weathering processes. Geochimica et Cosmochimica Acta, 61 (3): 677-682. [ Links ]
ALEXANDRE, A.; MEUNIER, J.D.; LEZINE, A.M.; VINCENS, A. & D. Schwartz., 1997b.- Phytoliths: indicators of grassland dynamics during late Holocene in intertropical Africa. Palaeogeography, Palaeoclimatology, Palaeoecology, 136: 213-229. [ Links ]
ALEXANDRE, A.; MEUNIER, J.D.; MARIOTTI, A. & FRANCOIS, S., 1999.- Late Holocene phytolith and Carbon-Isotope record from a Latosol et Salitre, South Central Brazil. Quaternary Research, 51: 187-194. [ Links ]
ALZATE, M.E., 1995.- Neotectónica de la falla San ruperto. Municipio de Frontino, Antioquia: Tesis de pregrado, Universidad Nacional de Colombia, Medellín. [ Links ]
ARCHILA, S., 2005.- Arqueobotánica en la Amazonía colombiana. Un modelo etnográfico para el análisis de maderas carbonizadas. FIAN, UNIANDES-CESO, Bogotá. [ Links ]
BOULDER, M., 1994.- An approach to standard terminology for palynodebris (en) TRAVERSE, A. (ed.) Sedimentation of organic particles. Cambridge University Press, Cambridge. [ Links ]
BOZARTH, S., 1987.- Diagnostic opal phytoliths from rinds of selected Cucurbita species. American Antiquity, 52 (3): 607-615. [ Links ]
__________., 1992.- Classification of opal phytoliths formed in selected dicotyledons native in the great plains (en) RAPP, G. & MULHOLLAND, S. (eds.) Phytolith Systematics. Plenum Press, New York. [ Links ]
BREMOND, L.; ALEXANDRE, A.; HELY, C. & GUIOT, J., 2005.- A phytolith index as a proxy of tree cover density in tropical areas: calibration with Leaf Area Index along a forest-savanna transect in southestern Cameroon. Global and Planetary Change, 45: 277-293. [ Links ]
BUSH, M. & COLINVAUX, P., 1994.- Tropical forest disturbance: a paleoecological records from Darien, Panamá. Ecology, 75 (6): 1761-1768. [ Links ]
CABRERA, G.; FRANKY, C. & MAHECHA, D., 1999.- Los Nukak. Nómadas de la Amazonia colombiana. Editorial Unibiblos. Universidad Nacional de Colombia. Programa COAMA. Gobierno de Dinamarca. [ Links ]
CAVELIER, I.; RODRÍGUEZ, C.; HERRERA, L.; MORCOTE, G. & MORA, S., 1995.- No sólo de caza vive el hombre. Ocupación del bosque amazónico, Holoceno temprano (en) CAVELIER, I.; RODRÍGUEZ, C.; HERRERA, L.; MORCOTE, G. & MORA, S. (eds.) Ámbito y ocupaciones tempranas de la América tropical. Instituto Colombiano de antropología. Colcultura. Fundación ERIGAIE, Bogotá. [ Links ]
CIEZA DE LEÓN, P., 1984 (1589).- La crónica del Perú. Edición de Manuel Ballesteros. Historia 16. Madrid. [ Links ]
COLINVAUX, P. & BUSH, M., 1991.- The rain forest ecosystem as a resource for hunting and gathering. American Anthropologist, 93 (1):83-106. [ Links ]
DESCOLA, P., 1987.- La Selva Culta: simbolismo y praxis en la ecología de los Achuar. Ediciones Abya-Yala, Ecuador. [ Links ]
DÍAZ, C. & RIVERA, C.A., 2004.- Diatomeas de pequeños ríos andinos y su utilización como indicadoras de condiciones ambientales. Caldasia, 26 (2): 381-394. [ Links ]
ERCEGOVAC, M. & KOSTIC, A., 2006. Organic facies and palynofacies. Nomenclature, classification and applicability for petroleum source rock evaluation. International Journal of Coal Geology, 68: 70-78. [ Links ]
ESPINAL, L.S., 1992.- Geografía ecológica de Antioquia. Zonas de vida. Universidad Nacional de Colombia, Medellín. [ Links ]
FLÓREZ, M.T., 2000.- Génesis de paleosuelos ándicos a partir del estudio de pedocomponentes: Tesis de Maestría, Universidad Nacional de Colombia, Medellín. [ Links ]
FLÓREZ, M.T. & PARRA, L.N., 1999.- Fitolitos en los paleosuelos ándicos Altoandinos, San Félix, departamento de Caldas (en) FLÓREZ, M.T. & LOZANO, G.L. (eds.) Silicofósiles Altoandinos. Colciencias-BID, Universidad Nacional de Colombia, Medellín. Universidad de Antioquia, Universidad de Ponta Grossa, Brasil. [ Links ]
FLÓREZ, M.T.; PARRA, L.N. & JARAMILLO, D., 2006.- Los fitolitos como herramienta pedogenética de un Andisol de la cuenca de Piedras Blancas. Suelos Ecuatoriales. Revista de la Sociedad Colombiana de Ciencia del Suelo, 36 (1): 43-51. [ Links ]
FONTURBEL, F. & MOLINA, C., 2002.- Mecanismos y estrategias de coevolución en plantas: un breve análisis de la coevolución planta-insecto. Santiago de Chile Ciencia abierta, Universidad de Chile. [ Links ]
GASTALDO, R.A., 1994.- The génesis and sedimentation of phytoclasts with examples from coastal environments (en) TRAVERSE, A. (ed.) Sedimentation of organic particles. Cambridge University Press, Cambridge. [ Links ]
GNECCO, C., 2000.- Ocupaciones tempranas en bosques tropicales de montaña. Editorial Universidad del Cauca, Popayán. [ Links ]
GUREVITCH, J.; SCHEINER, S. & FOX, G., 2006.- The ecology of plants. Second edition. Sinauer Associates, INC. Publishers, Sunderland, Massachusetts. [ Links ]
HERRERA, L.F., 1985.- Agricultura aborigen y cambios en la vegetación en la Sierra Nevada de Santa Marta. Fundación de Investigaciones Arqueológicas Nacionales, Banco de la República, Bogotá. [ Links ]
LANGEBAEK, C.H., 1992.- Noticias de caciques muy mayores. Origen y desarrollo de sociedades complejas en el nororiente de Colombia y norte de Venezuela. Ediciones Uniandes, Bogotá. [ Links ]
MADELLA, M.; ALEXANDRE, A. & BALL, T., 2005.- International Code for phytolith nomenclature 1.0. Annals of Botany, 96: 253-260. [ Links ]
MILLER-ROSEN, A., 1989.- Ancient town and city sites: a view from the microscope. American Antiquity, 54 (3): 564-578. [ Links ]
_______________., 1992.- Preliminary identification of silica skeletons from near eastern archaeological sites: an anatomical approach (en) G. RAPP & S. MULHOLLAND (eds.) Phytolith Systematics. Emerging issues. Plenum Press, New York. [ Links ]
MONSALVE, C., 2004.- Palinología del Holoceno superior en la laguna Puente Largo, Páramo de Frontino, Antioquia, Cordillera Occidental colombiana: Tesis de Maestría, Universidad Nacional de Colombia, Medellín. [ Links ]
MONTEJO, R. & RODRÍGUEZ, E., 2001.- Antiguos pobladores y labranzas en el valle medio del río Otún, Risaralda. Boletín de Arqueología, 16 (1): 37-115. [ Links ]
MORCOTE, G. & BERNAL, R., 2001.- Remains of Palms (Palmae) at archaeological sites in the new world: a review. The botanical Review, 67 (3): 186-206. [ Links ]
NADELHOFFER, K.; BOWDEN, R.; MCDOWELL, W. & LAJTHA, K., 2001.- Soil organic matter formation: insights from chronic manipulations of plant inputs to soils. In Press. www.ansp.org/~goulden/2001_MLTER_Meeting_Papers/Nadelhoefer.pdf [ Links ]
PARRA, L. & FLÓREZ, M., 2001.- Propuesta de clasificación morfológica para los fitolitos Altoandinos colombianos. Crónica forestal y del medio ambiente, 16. Posgrado en Bosques y Conservación Ambiental, Universidad Nacional de Colombia, sede Medellín. [ Links ]
PARRA-SÁNCHEZ, L.N., 2005.- Análisis facial de alta resolución de sedimentos del Holoceno tardío en el Páramo de Frontino, Antioquia: Tesis Doctoral, Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Bogotá. [ Links ]
PAUSTIAN, K. & SCHNURER, J., 1987.- Fungal growth response to carbon and nitrogen limitations: application of a model to laboratory and field data. Soil Biochemistry, 19 (5): 26-39. [ Links ]
PIAZZINI, E.; POSADA, W.; ESCOBAR, D. & ARANGO, C., 2009.- Arqueología de Frontino. Espacio, tiempo y sociedad en el noroccidente de Antioquia durante la época precolombina y colonial. Informe. Gobernación de Antioquia, Banco de la República, Universidad de Antioquia. [ Links ]
PIÑEROS, F., 1989.- Análisis de polen del cráter de Merhenberg (en) HERRERA, L.F.; DRENNAN, R. & URIBE, C. A. (eds.) Cacicazgos prehispánicos del Valle de la Plata, tomo 1: el contexto medioambiental de la ocupación humana. University of Pittsburg, Universidad de los Andes. [ Links ]
PIPERNO, D., 1988. Phytolith Analysis. An archaeological and geological perspective. Academic Press, INC, San Diego, CA. [ Links ]
__________., 2007.- Fitolitos y carbón; evidencia de agricultura de roza y quema prehistórica en el bosque húmedo tropical de Darién en Panamá: 601-607 (en) E. GILES; E. ALLEN; J. JACKSON; F. SANTOS-GRANERO (eds.) Ecología y evolución en los trópicos. Editora Nova Art, Panamá. [ Links ]
__________., 2009.- Identifying crop plants with phytoliths (and starch grains) in Central and South America: A review and an update of the evidence. Quaternary international, 193: 146-159. [ Links ]
PIPERNO, D. & PEARSALL, D., 1998.- The origins of agriculture in the lowland neotropics. Academic Press, INC, San Diego, CA. [ Links ]
PIPERNO, D.; ANDRES, T. & STOTHERT, K., 2000.- Phytoliths in Cucurbita and other neotropical Cucurbitaceae and their occurrence in early archaeological sites from the lowland American tropics. Journal of Archaeological Science, 27: 193-208. [ Links ]
PIPERNO, D.; HOIST, I.; WESSEL-BEAVER, L. & ANDRES, T., 2002.- Evidence for the control of phytolith formation in Cucurbita fruits by the Hard Rind (Hr) genetic locus: archaeological and ecological implications. Proceedings of the National Academy of Sciences of the United States of America, 99 (16):13-27. [ Links ]
PLAZAS, C.; FALCHETTI, A.; SÁENZ, J. & ARCHILA, S., 1993.- La sociedad hidráulica Zenu. Estudio arqueológico de 2000 años de historia en las llanuras del Caribe colombiano. Banco de la República, Museo del Oro, Santafé de Bogotá. [ Links ]
POLITIS, G., 1996.- Nukak. Instituto Amazónico de Investigaciones Científicas, Bogotá. [ Links ]
POSADA, W.; PARRA, L. & JARAMILLO, D., 2009.- Procesos naturales y procesos antrópicos a escala de sitio. Un caso de geoarqueología en el municipio de Frontino, noroccidente colombiano. En prensa. Arqueología del Área Intermedia. Sociedad Colombiana de Arqueología. [ Links ]
POSADA, W.; VERGARA, D. & PARRA, L., 2009.- Fitolitos de algunas plantas útiles y cultivadas por la comunidad Embera del Llano, Municipio de Frontino, Antioquia. Informe sin publicar. Universidad Nacional de Colombia, Medellín. [ Links ]
PREBBLE, M.; SCHALLENBERG, M.; CRTER, J. & SHULMEISTER, J., 2002.- An analysis of phytolith assemblages for the cuantitative reconstruction of late Quaternary environments of lower Taieri plain, Otago, South Island, New Zeland I. Modern Assemblages and transfer functions. Journal of Paleolimnology, 27: 393-413. [ Links ]
RANERE, A. & COOK, R., 1995.- Evidencias de ocupación humana en Panamá a postrimerías del Pleistoceno y a comienzos del Holoceno (en) CAVELIER, I.; RODRÍGUEZ, C.; HERRERA, L.; MORCOTE, G. & MORA, S. (eds.) Ámbito y ocupaciones tempranas de la América tropical. Instituto Colombiano de antropología, Colcultura, Fundación ERIGAIE, Bogotá. [ Links ]
RAPP, G. & MULLHOLAND, S., 1992.- Phytolith systematics: an introduction (en) RAPP, G. & MULLHOLAND, S. (eds.) Phytolith Systematics. Emerging issues. Plenum Press, New York. [ Links ]
REICHEL-DOLMATOFF, G., 1997.- Arqueología de Colombia. Un texto introductorio. Biblioteca Familiar Presidencia de la República, Bogotá. [ Links ]
RESTREPO, D.; ORREGO, C. & SYLVA, D., 1989.- Estudio florístico de la cuenca alta del río Cuevas. Municipio de Frontino, Antioquia: Tesis de pregrado, Universidad Nacional de Colombia, Medellín. [ Links ]
RICH, F. & PIRKLE, F., 1994.- Paleoecological interpretation of the trail ridge sequence, and relates deposits in Georgia and Florida, based on pollen sedimentation and clastic sedimentology (en) TRAVERSE, A. (ed.) Sedimentation of organic particles. Cambridge University Press, Cambridge. [ Links ]
RODRÍGUEZ, C., 1995.- Asentamientos de los bosques subandinos durante el Holoceno medio (en) CAVELIER, I.; RODRÍGUEZ, C.; HERRERA, L. MORCOTE, G. & MORA, S. (eds.) Ámbito y ocupaciones tempranas de la América tropical. Instituto Colombiano de antropología, Colcultura, Fundación ERIGAIE, Bogotá. [ Links ]
SALGADO, H. & STEMPER, D., 1995.- Cambios en alfarería y agricultura en el centro del litoral Pacífico colombiano durante los dos últimos milenios. Fundación de Investigaciones Arqueológicas Nacionales, Banco de la República, INCIVA, Santa fe de Bogotá. [ Links ]
SCHERY, R., 1956.- Plantas útiles al hombre. Barcelona: Salvat editores. [ Links ]
SCHIFFER, M., 1987.- Formation Processes of the Archaeological Record. University of Utah Press, Utah, Salt Lake City. [ Links ]
SCOTT, L., 1992.- Ilustrated phytoliths from assorted food plants (en) RAPP, G. & MULHOLLAND, S. (eds.) Phytolith Systematics. Emerging issues. Plenum Press, New York. [ Links ]
SEBAG, D.; COPARD, Y.; DI-GIOVNNI, C.; DURAND, A.; LAIGNEL, E.; OGIER, S. & LALLIER-VERGES, E., 2006.- Palynofacies as useful tool to study origins and transfers of particulate organic matter in recent terrestrial environments: synopsis and prospects. Earth Science Reviews, 79: 241-259. [ Links ]
THOMPSON, J., 1994.- The coevolutionary process. University of Chicago Press, Chicago. [ Links ]
TRAVERSE, A., 1994.- Sedimentation of land derived palynomorphs in the Trinity Galveston bay area, Texas (en) TRAVERSE, A. (ed.) Sedimentation of organic particles. Cambridge University Press, Cambridge. [ Links ]
TSARDSIDOU, G.; LEV-YADUM, S.; ALBERT, R.; MILLER-ROSEN, A.; EFSTRATIOUS, N. & WEINER, S., 2007.- The phytolith archaeological record: strenghts and weaknesses evaluated base don a cuantitative modern reference collection from Greece. Journal of Archaeological Science, 34: 1262-1275. [ Links ]
TYSON, R.V., 1995.- Sedimentary organic matter. Organic facies and palynofacies. Chapman and Hall, London, U.K. [ Links ]
VAN DER HAMMEN, T., 1992.- Historia, ecología y vegetación. Fondo de Promoción de Cultura Banco Popular, Fondo FEN Colombia, Corporación Colombiana para la Amazonía -Araracuara-, Bogotá. [ Links ]
VELÁSQUEZ, I., 1993.- Flórez e Insectos en la dieta prehispánica y actual de México. Folleto. [ Links ]
VELÁSQUEZ, C., 2005.- Paleoecología de alta resolución del Holoceno tardío. Páramo de Frontino. Universidad Nacional de Colombia, Medellín. [ Links ]
ZAMORA, J., 2005.- Los escarabeidos coprófagos (Coleóptera, Scarabaeoideae) en la evaluación del estado de conservación de agroecosistemas tradicionales mediterráneos. Boletín de la Sociedad Española de entomología, 29 (3-4): 99-120. [ Links ]

