










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.14 no.2 Manizales July/Dec. 2010
VARIATIONS IN THE AVAILABILITY OF FLESHY FRUITS IN A FOREST OF THE COLOMBIAN MAGDALENA MEDIO
* FR: 4-VI-2010. FA: 19-VII-2010
1 Ingeniera Agrónoma, Universidad de Caldas. Proyecto Primates Colombia. E-mail: paogirbe@gmail.com
2 Center for the Study of Human Origins, Department of Anthropology, New York University. Proyecto Primates Colombia. E-mail: al898@nyu.edu
Entre enero de 2006 y julio de 2008, estudiamos la variación temporal en el número de plantas con frutos carnosos maduros en un remanente de bosque húmedo tropical (Bh-T) en la Serranía de las Quinchas, Magdalena Medio colombiano. Hicimos recorridos a través del bosque, durante 61 quincenas, empleando un transecto de 8294 m de longitud. Registramos la presencia de frutos sobre el transecto, identificamos la planta parental, su distancia perpendicular al centro del transecto y la dirección; paralelamente, registramos la precipitación diaria. Calculamos el número de plantas fructificando cada quincena, en términos de densidad y realizamos una regresión entre ésta y la precipitación. En este estudio registramos 773 individuos pertenecientes a 145 especies y 37 familias, lo que corresponde a una representatividad entre 85% y 89%. Los meses con mayor número de plantas en fructificación fueron marzo, abril y diciembre mientras que enero, junio y octubre presentaron un menor número; variación que estuvo determinada en un 6% por el régimen de lluvias. Este estudio nos permitió resaltar 15 especies clave para el sostenimiento del gremio de frugívoros de la zona, las cuales muestran picos de fructificación que se distribuyen en el tiempo y conjuntamente producen frutos durante todo el año. Estos resultados pueden ser empleados para diseñar planes de conservación y restauración del Bh-T.
Palabras clave: bosque húmedo tropical, climadiagrama, frugivoría, frutos carnosos maduros, precipitación.
AbstractBetween January, 2006 and July, 2008, we studied the temporal variation in the number of plants with ripe fleshy fruit in a remnant tropical humid forest (Bh-T) in the mountainous region Serranía de las Quinchas, in the Colombian Magdalena Medio. We traveled across the forest during 122 weeks crossing a 8,294 m. length transect. We registered the presence of fruit on this transect, we identified the parental plants, their perpendicular distance from the center of the transect and their direction; at the same time we recorded the daily rainfall. We calculated the number of fruiting plants in terms of density every other week and we carried out a regression between density and rainfall. In this study we registered 773 individuals belonging to 145 species and 37 families which correspond to 85% and 89% representativeness. The months with higher quantities of fruiting plants were March, April and December while January, June and October presented lower quantities; this variation was determined in a 6% because of the rainfall periods. This study allowed us to highlight 15 key species for the sustainability of the frugivorous community in the area, since such species show productivity peaks distributed in time and together produce fruits all year long. These results can be used to design Bh-T conservation and restoration plans.
Key words: tropical humid forest, climate diagram, frugivory, rainfall, ripe fleshy fruits.
El bosque húmedo tropical (Bh-T) se caracteriza por ser uno de los ecosistemas de mayor complejidad estructural, estratificación y diversidad de especies en el mundo (ETTER, 1998). Muchas de estas características están relacionadas con complejas redes de interacciones que se forman entre animales frugívoros y plantas (BASCOMPTE & JORDANO, 2008; CASTAÑO, 2009).
Un alto porcentaje de las plantas leñosas en el Bh-T (entre 79% y 94%) dependen de los animales frugívoros para dispersar sus semillas (CHAPMAN & WRANHAM, 1994; JORDANO, 2000). A su vez, muchos vertebrados silvestres principalmente mamíferos y aves, pero también algunos reptiles, son frugívoros en mayor o menor grado y dependen de las frutas para su supervivencia, al menos durante una parte del año (FLEMING et al., 1987; JORDANO, 2000; WALLACE & PAINTER, 2002). Por esta razón, la frugivoría es un proceso esencial en poblaciones de plantas del Bh-T, donde la regeneración natural es fuertemente dependiente de la dispersión de semillas por animales (JORDANO, 2000).
Diferentes estudios enfocados en la ecología de fructificación de árboles tropicales y sus consumidores, han tenido como objetivo cuantificar la disponibilidad de frutos carnosos (CHAPMAN et al., 1992), ya que importantes aspectos cíclicos de los frugívoros, tales como reproducción, cría, movimientos migratorios, abundancia y comportamiento, son el reflejo de los patrones de abundancia de los recursos alimenticios y están asociados con los picos estacionales de fructificación (JORDANO, 2000; ROLDÁN & LARREA, 2003; STEVENSON, 2004).
Los factores climáticos y bióticos que podrían explicar la estacionalidad de la fructificación son aún materia de discusión (MOLINARI, 1993; STEVENSON, 2004). En términos generales, se ha sugerido que la maduración de los frutos de muchas especies de plantas tropicales ocurre durante la época de lluvias (WILLIAMS-LINERA & MEAVE, 2002), y varios estudios han registrado que existen uno o dos picos de fructificación durante la época más húmeda y un período relativamente corto de baja disponibilidad durante la estación seca, en este momento se ha detectado la presencia de especies vegetales clave que mantienen al gremio comunidad de frugívoros (ROLDÁN & LARREA, 2003). Por esta razón, los estudios fenológicos son fundamentales para comprender la dinámica de los recursos en los ecosistemas (VÍLCHEZ et al., 2004) y entender su influencia sobre la ecología y comportamiento social de los frugivoros que dependen de éstos.
En ambientes complejos tales como el Bh-T, comúnmente se han realizado estimaciones de producción de frutos empleando transectos fenológicos, que evalúan la disponibilidad de recursos para especies de alta movilidad que se alimentan de una gran variedad de frutos carnosos (STEVENSON, 2004). Este método, permite cubrir una amplia área de muestreo e incrementa la probabilidad de encontrar especies que no son registradas con facilidad por otros métodos (CHAPMAN et al., 1992; HEMINGWAY & OVERDORFF, 1999; WALLACE & PAINTER, 2002; STEVENSON, 2004).
El objetivo de este trabajo fue cuantificar la disponibilidad de plantas con frutos carnosos maduros, evaluar la variación quincenal a lo largo de un año, estimar la relación entre esta variación y la precipitación. Por último, detectar la presencia de especies vegetales clave para el sostenimiento de los frugívoros presentes en uno de los últimos relictos de Bh-T ínter-andino ubicado en la Serranía de las Quinchas, importante centro de endemismos del país (HERNÁNDEZ-CAMACHO et al., 1992; LAVERDE et al., 2005).
ÁREA DE ESTUDIOHicimos este trabajo en un remanente de bosque, de ahora en adelante llamado "La Reserva", ubicado a 6º 3' N y 74º 3' O en el departamento de Boyacá y hace parte de la Reserva Natural de las Aves El Paujíl de PROAVES (Figura 1). La zona de estudio tiene un área de 81,3 hectáreas, una elevación de 260 ± 30 msnm y una temperatura media anual de 28,4 ºC (T. min 24,4 ºC y T. max 32,4 ºC), datos registrados durante el tiempo de muestreo (2006-2008). Este lugar tiene una historia de deforestación y tala selectiva de aproximadamente 10-12 años, donde fueron extraídos árboles madereros de gran porte y tamaño (GUERRERO, 2007). Sin embargo, después del año 2003 la extracción de madera ha cesado y "La Reserva" ha sido empleada para realizar estudios de ecología y conservación del mono araña café (Ateles hybridus), con este fin, se ha construido un sistema de trochas debidamente georeferenciadas y marcadas cada 25 m, las cuales fueron empleadas en el desarrollo del presente estudio.
MÉTODOS
Datos climáticos
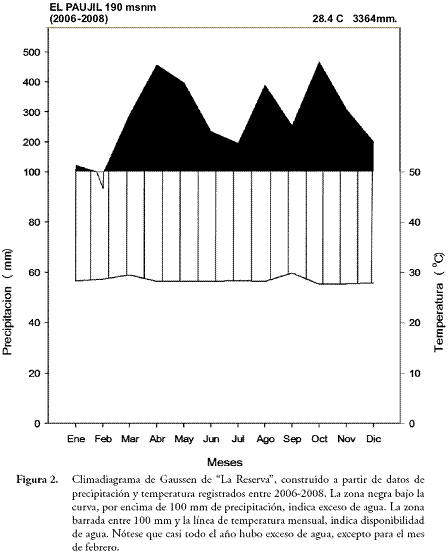
Entre el 17 de enero de 2006 y el 15 de julio de 2008 registramos diariamente a las 6:00 y a las 18:00 horas los datos de precipitación y temperatura. Para esto empleamos un pluviómetro manual y un termo-higrómetro digital de máximas y mínimas. Con los datos construimos un climadiagrama de Gaussen de "La Reserva".
Transectos fenológicos
Para cuantificar la variación en el número de plantas con frutos carnosos maduros empleamos un transecto de 8294 m, formado por el sistema de trochas ya mencionado. A través del cual hicimos recorridos durante 61 quincenas, en busca de frutos carnosos que se encontraran sobre el transecto. Posteriormente, identificamos la planta de la cual provenían los frutos (planta parental), medimos la distancia perpendicular (DP) de cada planta al centro del transecto y la dirección (Izquierda-Derecha). De esta forma, registramos la presencia de plantas con frutos carnosos maduros en cada quincena.
Construimos curvas de acumulación de especies con ayuda del programa EstimateS (COLWELL, 2005), para conocer la representatividad del muestreo, en función de las especies con frutos carnosos maduros registradas en cada quincena. Estas curvas y sus estimadores no paramétricos (ACE y Chao 1) nos permitieron conocer la representatividad de especies con frutos carnosos maduros con respecto al total de especies estimadas (COLWELL & CODDINGTON, 1994).
Para hallar la densidad de arboles por quincena, empelamos las DP de las plantas al centro del transecto y calculamos un área efectiva de muestreo (AEM = longitud del transecto * 2 ancho efectivo de detección), con el programa Distance 5.0 (THOMAS et al., 2006). El ancho efectivo de detección (AED) es la distancia perpendicular al transecto en que la especie puede ser registrada con alta confiabilidad, dependiendo de la distribución que muestren los registros de la DP (THOMAS et al., 2002). Para cada quincena promediamos el número de plantas en fructificación y la precipitación, de esta forma cada quincena representa el promedio de los años evaluados. Realizamos un ANDEVA de una vía para evaluar el efecto del tiempo (quincenas) en la variación de plantas con frutos carnosos maduros. Finalmente, hicimos una regresión exponencial entre la precipitación y las plantas con frutos carnosos maduros.
Colectamos muestras de las plantas con frutos carnosos, registradas durante el muestreo, las cuales fueron determinadas por especialistas de la Universidad de los Andes y depositadas en el herbario de la misma universidad (ANDES).
RESULTADOS Y DISCUSIÓN Datos climáticos
El régimen de lluvias fue bimodal-tetraestacional, con una precipitación media anual de 3364 mm y una media mensual de 280,3 mm. El primer periodo de lluvias fue entre abril y mayo, y el segundo estuvo repartido entre agosto y noviembre. Durante estos seis meses registramos el 66,2% del total de lluvias, siendo octubre el mes más lluvioso con 467 mm y febrero el más seco con 93,3 mm. Al construir el climadiagrama de Gaussen (Figura 2) encontramos que, en promedio para los tres años, no se presentaron meses secos o con deficiencia de agua. Según DI CASTRI & HAJEK (1976), un mes seco es aquel en que el total mensual de las precipitaciones, expresado en milímetros, es igual o inferior al doble de la temperatura media mensual, expresada en grados centígrados (P < ó = 2T). Sin embargo, en febrero de 2007, hubo deficiencia de agua para las plantas debido a una baja precipitación (11 mm). En general, el área de estudio presentó una precipitación ligeramente superior a la reportada por BALCÁZAR-VARGAS et al. (2000) para las estaciones climatológicas de Puerto Boyacá (Boyacá) y Otanche (Boyacá), lo cual puede estar asociado con la presencia de elevaciones montañosas cercanas al área de estudio, que actúan como barreras de choque de las nubes, y de esta forma incrementan la precipitación (JARAMILLO, 2005).
Transectos fenológicos
Durante los 30,5 meses de toma de datos, en un AEM de 7,3 ha, registramos un total de 773 individuos pertenecientes a 145 especies y 37 familias (Anexo 1). La Figura 3 muestra las curvas de acumulación de plantas con frutos carnosos maduros, registradas durante las 61 quincenas, con sus intervalos de confianza y los estimadores (ACE y Chao 1). La pendiente de la curva de acumulación de especies aún tiene una tendencia creciente (sin llegar a un valor asintótico), lo que indica que el muestreo de plantas no fue completo. Además, el número estimado de especies aún está por encima de los intervalos de confianza de las especies observadas. En este estudio, registramos 145 especies de plantas con frutos carnosos maduros y los estimadores ACE y Chao 1 calculan que, durante el tiempo de muestreo, en el área de estudio debería haber entre 162 y 170 especies de plantas respectivamente, lo cual quiere decir que alcanzamos una representatividad entre el 89,4% y el 85,2% de las especies que presentaron producción de frutos carnosos durante el periodo de estudio.
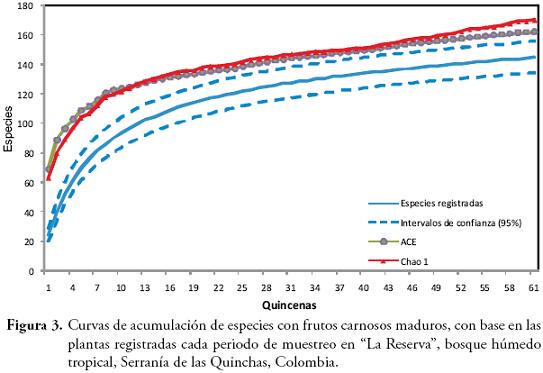
Algunas de las razones que pudieron influir, para que no hayamos alcanzado la saturación de especies de plantas con frutos carnosos maduros, son: la existencia de plantas con producciones de fruta discretas, y/o frutos poco llamativos, que no caen con frecuencia al suelo (ej. frutos de Cecropia spp.) y de plantas con frutos caulinares que sólo serían registrados si la planta parental estaba muy cerca del transecto (ej. frutos de la Fam. Sterculiaceae), lo cual disminuye la detectabilidad de la planta y afecta negativamente la saturación de la curva. Además de esto, en algunos grupos taxonómicos de gran diversidad como las plantas del Bh-T, resulta casi imposible registrar la totalidad de las especies presentes en un área determinada, ya que en estos ambientes hay especies que se encuentran en densidades extremadamente bajas (ej. uno o dos individuos cada 50 hectáreas), que posiblemente requieran de un esfuerzo de muestreo mayor al realizado en este estudio (JIMÉNEZ-VALVERDE & HORTAL, 2003; VILLARREAL et al., 2006). Por lo tanto consideramos que, pese a todas las dificultades inherentes en la obtención de la asíntota de la curva de acumulación de especies de plantas, nuestro muestreo representa razonablemente bien a las especies de plantas con frutos carnosos maduros en el área de estudio, durante el periodo de muestreo.
La producción de frutos carnosos mostró variación entre quincenas (F23,37 = 2,12; P = 0,02), donde los meses con mayor número de plantas en fruto fueron marzo y abril, seguido por diciembre. A su vez, los meses con menor número de plantas en fruto fueron enero, junio y octubre (Figura 4) y en ninguno de los meses reportamos ausencia de plantas con frutos. Con respecto a la contribución mensual, 15 especies de plantas fueron las más representativas, dado que éstas, tuvieron una participación de casi el 50% en el total de plantas con frutos carnosos para cada quincena (Figura 4), en orden de importancia fueron: Pourouma cf. melinonii (Pou mel), Apeiba tibourbou (Ape tib), Oenocarpus bataua (Oen bat), Oxandra venezuelana (Oxa ven), Pourouma sp.2 (Pou sp.2), Passiflora vitifolia (Pasvit), Passiflora cf. ambigua (Pas amb), Pouteria cf. lucuma (Pou luc), Cecropia cf. peltata (Cec pel), Solanum sp. (Sol sp.), Spondias mombin (Spo mom), Grias cf. phoetidisima (Gri pho), Virola cf. flexuosa (Vir fle), Dialium guianense (Dia gui) y Xylopia sp. (Xyl sp.). Cabe resaltar su importancia principalmente durante los meses de baja disponibilidad de fruta, ya que éstas podrían estar sosteniendo el gremio de frugívoros residentes en el área de estudio, convirtiéndose en especies vegetales clave.
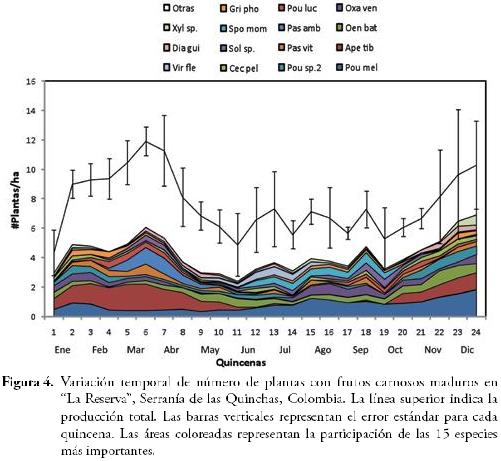
El número de plantas con frutos carnosos maduros por hectárea, presentó una débil relación con la precipitación registrada a lo largo del periodo de estudio (r² = 0,06; F = 3,6; P = 0,06), lo que indica que la estacionalidad en la disponibilidad de frutos carnosos en el Bh-T de "La Reserva" está influenciada por otras variables adicionales a la precipitación.
Otros autores han planteado que la fenología de frutos en las selvas tropicales es el producto de múltiples presiones selectivas, entre las que se agrupan interacciones con los frugívoros, con otros factores bióticos y climáticos (MOLINARI, 1993; WILLIAMS-LINERA & MEAVE, 2002). La conjunción de todos estos factores no evaluados en el presente estudio, podrían ser la causa del bajo efecto que tuvo la precipitación (6%) sobre la variación de plantas con frutos carnosos maduros. Además, el patrón fenológico presentado en este trabajo, es el resultado de la respuesta de múltiples especies y en algunos casos estas descripciones generales de los ritmos biológicos de las comunidades tienden a ocultar las respuestas propias de cada especie, ya que en algunos casos una sola especie puede representar toda la relación existente entre la fructificación y la precipitación (WILLIAMS-LINERA & MEAVE, 2002; MORALES, 2007).
Pese a lo anterior, algunos estudios en ambientes tropicales han encontrado una relación alta entre la producción de fruta y la precipitación, sin embargo, estos estudios se han llevado a cabo en ambientes donde el agua es un factor limitante (ej. sub-xerofíticos), y la formación de frutos depende del agua absorbida por la planta en la época de lluvias (SORIANO et al., 1991; CASADO-BERNABELA, 2009). Sin embargo, en "La Reserva", como lo muestra el climadiagrama, el agua no parece ser un factor limitante y ésta puede ser otra razón por la cual la relación encontrada entre la producción de frutos maduros y la precipitación fue baja.
Al observar la dinámica de la producción individualmente para las 15 especies más importantes (Figura 5), encontramos que éstas muestran picos de fructificación distribuidos en el tiempo, de manera que, conjuntamente producen frutos durante todo el año, de esta forma se podría favorecer la permanencia de las poblaciones actuales de frugívoros en la zona de estudio, tales como primates, otros mamíferos voladores y no voladores, aves , etc., ya que estos frugívoros encontrarían recursos alimenticios durante todo el año, incluso en las épocas de baja producción de fruta. Esta disponibilidad de frutos durante todo el año, ha sido resaltada por MOLINARI (1993) como uno de los factores que ha posibilitado el gran desarrollo de la frugivoría en los trópicos.
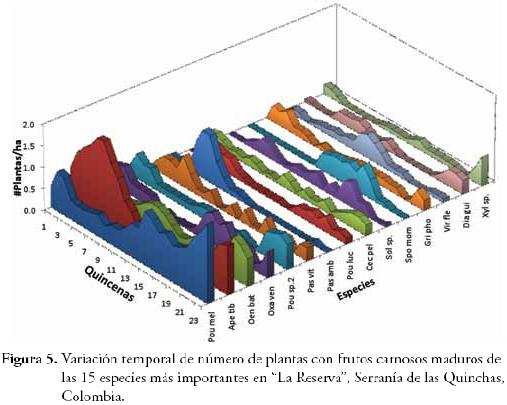
Así mismo, el patrón de fenologías distribuidas en el tiempo ha sido registrado para otros ensamblajes de plantas, y se ha planteado que, de esta manera se favorecería la convivencia de un alto número de especies, reduciendo la competencia interespecífica por el servicio de dispersión de semillas; esta interpretación supone una organización de la comunidad por medio de co-adaptaciones mutualistas entre plantas y sus dispersores (MOLINARI, 1993). A pesar de las fuertes críticas que han recibido estas suposiciones, estudios recientes han demostrado que las comunidades de plantas y animales mutualistas (como las plantas y sus dispersores o polinizadores) se organizan de manera co-evolutiva, permitiendo la convivencia de un alto número de especies (BASCOMPTE & JORDANO, 2008; CASTAÑO, 2009).
Finalmente, podemos concluir que la producción de frutos carnosos en "La Reserva" muestra una variación temporal que estuvo afectada sólo en un 6% por la precipitación, lo que indica que probablemente esté influenciada por factores adicionales como longitud del día u horas luz, diferencias de temperatura entre la planta y el ambiente, estado hídrico de la planta, o factores bióticos como patrones de actividad de sus dispersores y variación en los sistemas de polinización de las plantas. Por lo tanto, es necesario continuar en la búsqueda de las causas de los patrones de fructificación observados en este estudio, además de desarrollar estudios que nos permitan identificar la variación existente entre especies con respecto a sus patrones de fructificación.
Por otro lado, durante los meses de baja producción, observamos la presencia de plantas con frutos, lo que indica que las poblaciones de frugívoros pueden encontrar recursos alimentarios durante todo el año, lo cual favorece su permanencia en "La Reserva". Sin embargo, aún desconocemos en qué medida esta disponibilidad de frutos observada puede mantener o permitir un incremento en las poblaciones de frugívoros actuales. Por esta razón, sería de gran importancia poder complementar la información presentada en este trabajo con estudios comportamentales y poblacionales de los frugívoros en la zona.
En el desarrollo de este estudio pudimos evidenciar la importancia de 15 especies de plantas, ya que tuvieron una alta contribución en la producción total para cada quincena, por lo cual consideramos que estas especies podrían ser resaltadas como especies clave para la comunidad de frugívoros en "La Reserva", las cuales podrían ser implementadas en procesos de restauración del Bh-T en el Magdalena Medio.
AGRADECIMIENTOSEste trabajo fue financiado por Primate Conservation Inc., Conservation International Primate Action Fund y The Rufford Small Grants Foundation. Agradecemos especialmente a los diferentes integrantes del Proyecto Primates Colombia por su apoyo en la toma de datos. A la Fundación PROAVES por permitirnos realizar este trabajo en sus predios, en especial a Eliana Machado y a la familia Montero por su importante labor logística. A John Harold Castaño por sus valiosos aportes en la elaboración de este trabajo y al Dr. Pablo Stevenson por su ayuda en la identificación del material vegetal.
BALCÁZAR-VARGAS, M.P.; RANGEL, J.O. & LINARES E.L., 2000.- Diversidad florística de la Serranía de las Quinchas, Magdalena Medio (Colombia). Caldasia, 22 (2): 191-224. [ Links ]
BASCOMPTE, J. & JORDANO, P., 2008.- Redes mutualistas de especies. Investigación y ciencia, 384: 50-59. [ Links ]
CASADO-BERNABELA, R., 2009.- Consumidores de frutos y su relevancia en la dispersión y germinación de semillas del cactus globoso Melocactus schatzlii. Tesis de Maestría, Universidad de Los Andes, Facultad de Ciencias, Mérida, Venezuela. [ Links ]
CASTAÑO, J.H., 2009.- Murciélagos frugívoros y plantas quiropterocoras: descubriendo la estructura de sus interacciones mutualistas en una selva semicaducifolia. Tesis de Maestría, Universidad de Los Andes, Facultad de Ciencias, Mérida, Venezuela. [ Links ]
CHAPMAN, C.A.; CHAPMAN, L.J.; WRANGHAM, R.; HUNT, K.; GEBO, D. & GARDNER, L., 1992.- Estimators of fruit abundance of tropical trees. Biotropica, 24 (4): 527-531. [ Links ]
CHAPMAN, C.A. & WRANGHAM, R., 1994.- Indices of habitat-wide fruit abundance in tropical forests. Biotrópica, 26 (2): 160-171. [ Links ]
COLWELL, R.K., 2005.- EstimateS: statistical estimation of species richness and shared from samples. University of Connecticut, USA. [ Links ]
COLWELL, R.K. & CODDINGTON, J.A., 1994.- Estimating terrestrial biodiversity through extrapolation. Phil. Trans. R. Soc. Lond. B., 345 (1311): 101-118. [ Links ]
DI CASTRI, F. & HAJEK, E.R., 1976.- Bioclimatología de Chile. Universidad Católica de Chile. [ Links ]
ETTER, A., 1998.- Bosque húmedo tropical: 106-133 (en) CHAVES, M.E. & ARANGO, N. (eds.) Informe nacional sobre el estado de la diversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, PNUMA, Ministerio del Medio Ambiente. Bogotá, Colombia. [ Links ]
FLEMING, T.H.; BREITWISH, R. & WHITESIDES G.H., 1987.- Patterns of tropical vertebrate frugivore diversity. Ann. Rev. Ecol. Syst., 18: 91-109. [ Links ]
GUERRERO, J., 2007.- Descripción de algunos aspectos de la ecología y composición social de un grupo de Ateles hybridus (i. Geoffroyi-St. Hilaire, 1829) en la Serranía de las Quinchas, Colombia. Tesis, Pontificia Universidad Javeriana, Facultad de Estudios Ambientales y Rurales, Bogotá, Colombia. [ Links ]
HEMINGWAY, C.A. & OVERDORFF, D.J., 1999.- Sampling effects on food availability estimates: Pheological method, simple size and species composition. Biotrópica, 31 (2): 354-364. [ Links ]
HERNÁNDEZ-CAMACHO, J.; ORTIZ, R.; WALSHBURGER, T. & HURTADO, A., 1992.- Estado de la biodiversidad en Colombia: 41-225 (en) HALFFTER, G. (ed.) Diversidad biológica de Iberoamérica I. Instituto de Ecología, A.C. México. [ Links ]
JARAMILLO, R.A., 2005.- El clima andino y café en Colombia. Cenicafé. [ Links ]
JIMÉNEZ-VALVERDE, A. & HORTAL, J., 2003.- Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8: 151-161. [ Links ]
JORDANO, P., 2000.- Fruits and frugivory: 125-166 (en) FENNER, M. (ed.) Seeds: the ecology of regeneration in plant communities. CABI Publ., Wallingford, UK. [ Links ]
LAVERDE, O.R.; STILES, F.G. & MÚNERA, C., 2005.- Nuevos registros e inventario de la avifauna de la Serranía de las Quinchas, un área importante para la conservación de las aves (aica) en Colombia. Caldasia, 27 (2): 247-265. [ Links ]
MOLINARI, J., 1993.- El mutualismo entre frugívoros y plantas en las selvas tropicales: Aspectos paleobiológicos, autoecológicos, papel comunitario. Acta Biológica, Venezuela, 14 (4): 1-44. [ Links ]
MORALES, J.A., 2007.- Ecología alimentaria de tres especies de saltarines (Aves: Pipirdae) en una plantación forestal mixta de los Andes centrales colombianos. Tesis, Universidad de Caldas, Facultad de Ciencias Exactas y Naturales, Manizales. [ Links ]
ROLDÁN, A. & LARREA, D., 2003.- Fenología de 14 especies arbóreas y zoócoras de un bosque yungueño en Bolivia. Ecología en Bolivia, 38 (2) :125-140. [ Links ]
SORIANO, P.; SOSA, M. & RUSELL, O., 1991.- Hábitos alimentarios de Glossophaga longirostris Miller (Chiroptera: Phyllostomidae) en una zona árida de los Andes Venezolanos. Rev. Biol. Trop., 39 (2): 263-268. [ Links ]
STEVENSON, P., 2004.- Phenological patterns of woody vegetation at Tinigua park, Colombia: Methodological comparisons with emphasis on fruit production. Caldasia, 26 (1): 125-150. [ Links ]
THOMAS, L.; BUCKLAND, S.T.; BURNHAM, K.P.; ANDERSON, D.R.; LAAKE, JL.; BORCHERS, D.L. & STRINDBERG, S., 2002.- Distance sampling: 544-552 (en) EL-SHAARAWI, A.H. & PIEGORSCH, W.W. Encyclopedia of Environmetrics. Vol 1. John Wiley & Sons, Ltd, Chichester. [ Links ]
THOMAS, L.; LAAKE, J.L.; STRINDBERG, S.; MARQUES, F.F.C.; BUCKLAND, S.T.; BORCHERS, D.L.; ANDERSON, D.R.; BURNHAM, K.P.; HEDLEY, S.L.; POLLARD, J.H.; BISHOP, J.R.B. & MARQUES, T.A., 2006.- Distance 5.0. Research Unit for Wildlife Population Assessment, University of St. Andrews, UK. En: http://www.ruwpa.st-and.ac.uk/distance/ [ Links ]
VÍLCHEZ, B., CHAZDON, R. & REDONDO, A., 2004.- Fenología reproductiva de cinco especies forestales del Bosque Secundario Tropical. Kurú: Revista Forestal (Costa Rica), 1 (2): 1-10. [ Links ]
VILLARREAL, H.; ÁLVAREZ, M.; CÓRDOBA, S.; ESCOBAR, F.; FAGUA, G.; GAST, F.; MENDOZA, H.; OSPINA, M. & UMAÑA, A.M., 2006.- Métodos para el análisis de datos: una aplicación para resultados provenientes de caracterizaciones de biodiversidad: 185-226 (en) Manual de métodos para el desarrollo de inventarios de biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá. Colombia. [ Links ]
WALLACE, R.B. & PAINTER, R.L.E., 2002.- Phenological patterns in suothern Amazonian tropical forest: implications for sustainable management. Forest Ecology and Management, 160: 19-33. [ Links ]
WILLIAMS-LINERA, G. & MEAVE, J., 2002.- Patrones fenológicos: 407-431 (en) GUARIGUATA, M. & KATTAN, G. (eds.) Ecología y conservación de bosques neotropicales. Ediciones LUR, Costa Rica. [ Links ]







