










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.14 no.2 Manizales July/Dec. 2010
COLLEMBOLA (HEXAPODA) IN A THREE ESTABLISHMENT AGES SILVOPASTORAL SYSTEM AND IN A RICE-GROWING AREA OF THE TROPICAL DRY FOREST IN THE MUNICIPALITY OF PIEDRAS, TOLIMA
* FR: 18-III-2010. FA: 6-VIII-2010
1 Grupo de Investigación Biodiversidad y Dinámica de Bosques Naturales, Universidad del Tolima. E-mail: claudial98@yahoo.com
2 CORPOICA. C.I. Nataima, km 9 vía Espinal-Ibagué. E-mail: mardeloz@hotmail.com
3 Grupo de Investigación Genética, Biodiversidad y Fitomejoramiento, GEBIOME. Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Caldas. E-mail: fredy.rivera@ucaldas.edu.co
El presente estudio se realizó con el fin de contribuir al conocimiento de colémbolos, en el bosque seco tropical (bs-T). El área de trabajo correspondió a un sistema silvopastoril de tres edades de establecimiento y un área arrocera, ubicados a 4º29'6'' L.N. y 74º59'15'' L.O. hacienda El Chaco (Piedras - Tolima). Se establecieron parcelas de 1 hectárea para cada zona de estudio, en cada bloque se trazó una parcela central de 50 x 20 m, tomando 320 muestras (8 muestras por sistema, durante 10 meses) con un nucleador o barreno y procesadas en embudos de Berlese-Tullgren. Los 11.448 colémbolos capturados, comprenden 10 familias, 16 géneros y 17 morfoespecies. El sistema silvopastoril de 8 años fue el de mayor abundancia y densidad, y el de menor número de organismos el cultivo de arroz, explicable por las prácticas de manejo del cultivo. Se encontró un grupo de doce morfoespecies comunes a todos los sistemas y las morfoespecies Xenilla sp., Cyphoderus sp., Isotoma sp. y Mesaphorura sp., asociadas al sistema silvopastoril y Lepidocyrtus sp., exclusivo para el cultivo. Se presentó una alta dominancia, favorecida por un elevado número de organismos isotomidos. La abundancia de géneros, mostró diferencias significativas entre los sistemas y los meses de colecta. El análisis de correspondencia y la clasificación jerárquica, mostraron separación de poblaciones, los sistemas con mayor cobertura vegetal (silvopastoril 3 y 8 años), presentan mayor similaridad faunistica, y las comunidades más degradadas y con menor cobertura (arroz y silvopastoril 7 meses), son cercanas.
Palabras clave: suelo, sitio productivo, collembola.
AbstractThis study was carried out with the aim of contributing to the knowledge of collembola, in the tropical dry forest (bs-T). The area of study corresponded to a silvopastoral system of three establishment ages and a rice-growing area located at Latitude 4º29'6'' N and 74º59'15'' W, El Chaco estate (Piedras- Tolima). Three 1 hectare lots were established for each study site; in each block a central 50x20 m lot was traced taking 320 samples (8 samples per system during 10 months) with a drill in Berlese-Tullgren funnels. The 11,448 captured collembola include 10 families, 16 genera and 17 morphospecies. The 8 years silvopastoral system was the one that showed greater abundance and density and the least number of organisms in the rice-growing area which is understandable because of the practices in the cultivation management. A group of twelve morphospecies common to all the systems was found as well as the morphospecies Xenilla sp., Cyphoderus sp., Isotoma sp. and Mesaphorura sp., associated to the silvopastoral system and Lepidocyrtus sp., exclusive for the cultivation. A high dominance was present, favored by a high number of isotomidae organisms. The abundance of genera showed significant differences between the systems and the collection months. The correspondence analysis and the hierarchical classification showed a separation of populations, systems with greater vegetal coverage (silvopastoral 3 and 8 years), greater faunistic similarity, and the most degraded communities and with less coverage (rice and silvopastoral 7 months) are close.
Key words: soil, productivity site, collembola.
Los colémbolos son artrópodos pequeños abundantes en suelo y hojarasca, importantes por su influencia en la estructura del suelo, ya que participan en la descomposición de la materia orgánica, ciclaje de nutrientes, control de microorganismos y en las cadenas tróficas (CUTZ et al., 2003). Se conocen alrededor de 7.000 especies de colémbolos, con una amplia distribución geográfica en todos los continentes, se encuentran desde las nieves del Himalaya hasta los desiertos de Australia (PALACIOS et al., 2000). En Colombia se evidencia un gran desconocimiento del grupo, con tan sólo 12 especies registradas (MUTT, 1987), 16 familias y 62 géneros (PEÑARANDA et al., 2004), aunado a una información en el campo biológico y ecológico aún más incipiente.
En Colombia los sistemas agroforestales han estado presentes en el desarrollo agrícola, se estima que de los 40 millones de hectáreas de uso agrícola en el país, 3,7 millones son de uso agroforestal (URIBE, 2000). La implementación de sistemas agroforestales, contribuye a la recuperación de las características y capacidad productiva de los agroecosistemas. Dentro de los sistemas agroforestales, los silvopastoriles han demostrado la importancia de la integración del componente arbóreo en las pasturas, como elemento mejorador de las condiciones productivas en áreas dedicadas a la ganadería. En este contexto, y debido a la cantidad, diversidad y funciones de los colémbolos en el suelo, y ante la expansión actual de los sistemas silvopastoriles como alternativa para la producción sostenible, es necesario profundizar en la dinámica e interacciones de la flora, fauna, patógenos, biocontroladores y edáfofauna, asociados a especies vegetales en el sistema silvopastoril. En el presente trabajo se determinó la diversidad de colémbolos edáficos, encaminando la investigación a contribuir en la evaluación del componente biológico, que pueda servir de soporte en modelos silvopastoriles sostenibles en áreas arroceras potenciales de reconversión en el Valle Cálido del Alto Magdalena.
MATERIALES Y MÉTODOSEl trabajo de campo se realizó en la hacienda El Chaco, localizada a 4º29'6'' L.N y 74º59'15'' L.O (Piedras-Tolima). El muestreo se llevó a cabo en cuatro sitios seleccionados según el uso de la tierra: I. Cultivo de arroz; II. Sistema silvopastoril de siete meses de establecido; III. Sistema silvopastoril de tres años de establecido; IV. Sistema silvopastoril de ocho años de establecido. Se realizaron parcelas de 1 hectárea para cada zona de estudio, en cada bloque se trazó una parcela central de 50 x 20 m (CUTZ, 2003). Trazada la parcela, se tomaron 8 muestras de suelo de la capa arable (0-20 cm) al azar, con un nucleador de 10 cm de diámetro y una longitud de 10 cm, cada mes y por un periodo de 10 meses, para un total de 320 muestras. De las muestras almacenadas, se extrajo una muestra de suelo de cada uno de los bloques para realizar los análisis edáficos en el laboratorio de suelos de CORPOICA.
Las muestras se procesaron utilizando el método del embudo de Berlese-Tullgren. La fauna se colectó en frascos de vidrio de 125 ml con alcohol etílico al 70%. Los colémbolos se separaron y agruparon por morfotipos y estos a su vez por familias, simultáneamente se realizaron preparaciones permanentes en líquido de Hoyer para su determinación, siguiendo la técnica modificada de CHRISTIANSEN & BELLINGER (1980). Para la determinación de los ejemplares se utilizaron dibujos, fotografías y claves (CHRISTIANSEN & BELLINGER, 1980, 1981; PALACIOS, 1990, 1991; JANSSENS, 2003; OSPINA et al., 2003) entre otras, con ayuda de un microscopio de campo claro y observando algunas estructuras a través de la microscopía de contraste de fase. Los ejemplares determinados en placa, se corroboraron con colecciones del Museo Entomológico de la Universidad Nacional (UNAB) y del Laboratorio de Manejo Integrado de Plagas (MIP), del Centro Internacional de Agricultura Tropical (CIAT). Cada uno de los morfotipos fue gentilmente verificado, gracias a un convenio de investigación Universidad Nacional Autónoma de México (UNAM) - CIAT, por el Doctor José Guadalupe Palacios Vargas y la Ingeniera agrónoma Claudia Marcela Ospina.
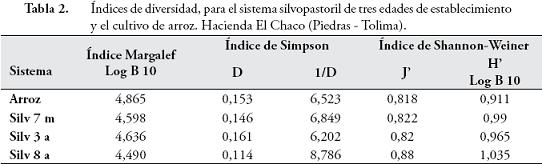
Se determinó la densidad anual por sitio, la diversidad Alfa fue evaluada teniendo en cuenta: 1. Métodos basados en la cuantificación del número de especies presentes (riqueza específica); 2. Métodos basados en la estructura de la comunidad. En este sentido, para evaluar la riqueza específica se calculó el índice de Margalef DMg = (S – 1) / Log N, se construyeron curvas de acumulación, que son gráficas del número acumulado de especies, en función de alguna medida del esfuerzo aplicado para obtener esa muestra, y que es una herramienta predictiva en estudios de biodiversidad, además se estimó la abundancia relativa de cada especie con el índice de Simpson:
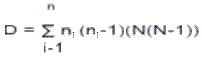
Se calculó el índice de equidad de Shannon-Wiener, el cual expresa la uniformidad de los valores de importancia a través de todas las especies de la muestra H' = – ∑pi ln pi.
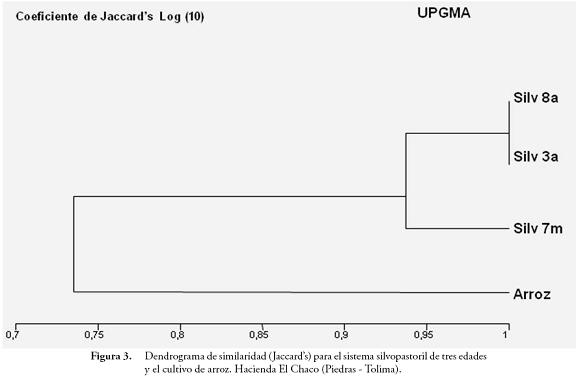
La diversidad Beta se evaluó con el índice cualitativo de Jaccard, que permite comparar la similitud de la composición de especies entre los agroecosistemas Ij = jl(a+b-j). Se efectuó un análisis factorial de correspondencia, para determinar los agrupamientos de los mismos en función de sus perfiles faunísticos. Las pruebas estadísticas se realizaron con los programas: MVSP 3.0 (Multi-variate Statistical Package); Estimates Win 800; BioDiversity Professional 2.
RESULTADOS Composición, distribución y densidad de colémbolos
La composición del orden Collembola en los sistemas silvopastoriles y el cultivo de arroz correspondieron a 11.448 ejemplares, agrupados en 17 morfoespecies, 16 géneros y 10 familias (Tabla 1). La familia Isotomidae representó el 70% de los individuos, este patrón se apreció también en cada una de las zonas de muestreo. La familia Entomobrydae alcanzó un 8% y el 22% restante correspondió a las otras ocho familias. La abundancia de Microisotoma sp. (Figura 1), alcanzó el 19% del total de los organismos colectados, seguido por el 16% de Folsomides americanus, 15% de Isotomiella sp., y un 13% para Proisotoma sp., las 13 morfoespecies restantes ocuparon tan sólo el 36% de la abundancia total (Figura 2).

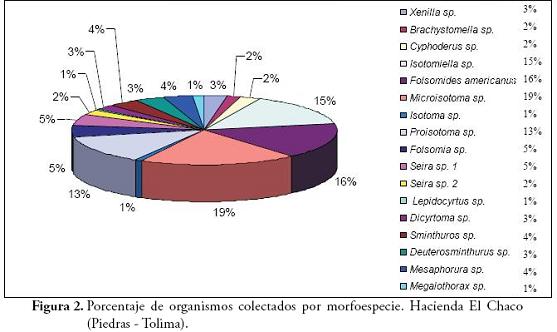
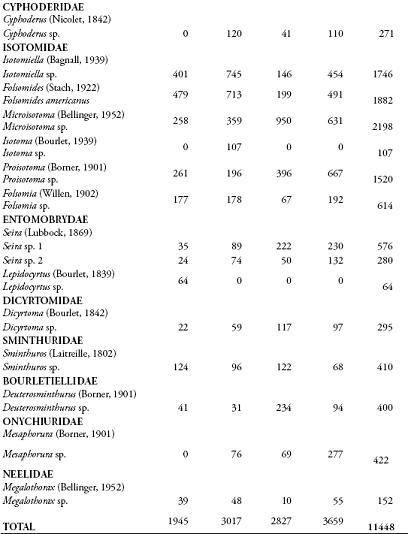
Folsomides americanus es la especie más abundante (25%) para el sistema arroz, seguida por la morfoespecie Isotomiella sp. (21%), Proisotoma sp. y Microisotoma sp. con un 13 %. Para el sistema silvopastoril de 7 meses las morfoespecies mejor representadas son Isotomiella sp. con un 25%, F. americanus con un 24% y Microisotoma sp. con el 12%. Para el sistema silvopastoril de 3 años, Microisotoma sp. alcanzó un 34%, seguido por Proisotoma sp. con un 14%. Y para el sistema silvopastoril de 8 años, las morfoespecies más abundantes son Microisotoma sp. y Proisotoma sp. con un 17 y 18%, respectivamente.
Teniendo en cuenta el número de colémbolos por m-2, y un área del barreno de 7.854 x 10-3 m-2, se encontró que el sistema con mayor número de organismos es el silvopastoril 8 años (5823,5), y el de menor densidad el cultivo de arroz (3095,6), donde no se encontró diferencia entre el número de organismos y los sistemas a través de los muestreos.
Medición de la diversidad Alfa
El índice de Margalef presentó una alta diversidad en cuanto a la riqueza de especies, mientras que el índice de Shannon-Weiner, mostró valores considerados como bajos (≤ 1) en todos los sistemas, presentando dominancia de algunas especies como Microisotoma sp., que se ratificó con el Índice de Simpson (Tabla 2).
Las curvas de acumulación de especies basadas en el promedio del estimador Jack 2 con base en la presencia/ausencia de las especies en las muestras tomadas mensualmente, comprobaron que el muestreo realizado fue suficiente para hallar el número de especies en cada sistema, debido al comportamiento asintótico de los estimadores. Para los sistemas silvopastoril siete meses y ocho años se logró capturar el 100% de las especies estimadas con Jack 2 y para los sistemas arroz y silvopastoril tres años, este porcentaje alcanzó el 96,43% y 95,8%, respectivamente.
Medición de la diversidad Beta
El índice cualitativo de similaridad de Jaccard's (Figura 3), mostró dos agrupamientos en cuanto a la composición faunística: un grupo correspondiente a los tres sistemas silvopastoriles y el otro al cultivo arroz, donde los silvopastoriles de 3 y 8 años, comparten las mismas especies y el cultivo de arroz muestra la menor similaridad en cuanto a especies comunes. Este comportamiento se corroboró bajo el criterio del índice cuantitativo de Bray-Curtis, donde se observó que el sistema silvopastoril 7 meses es el más similar al cultivo de arroz y el silvopastoril 3 años y el cultivo son los más disímiles.
El análisis factorial de correspondencia (Figura 4), ubicó a los sistemas silvopastoriles de 3 y 8 años como cercanos, así mismo muestra la cercanía entre el silvopastoril 7 meses y el cultivo de arroz. También se observó algunas morfoespecies asociadas a los diferentes sistemas, como Lepidocyrtus sp., al cultivo de arroz, e Isotoma sp., al silvopastoril 7 meses, así como un grupo de morfoespecies comunes a los sistemas silvopastoriles.
DISCUSIÓN Composición, distribución y densidad de colémbolos
Todas las familias son similares a las encontradas por OSPINA et al. (2003), en cultivos de pasto en el departamento de Antioquia, y por CASSAGNE et al. (2004) en bosques de confieras. Los géneros son similares a los reportados por OSPINA et al. (2004), en cultivos de maíz y algodón en el Valle del Cauca. Es de resaltar, que para cultivos de arroz no existía ningún reporte de la clase Collembola en Colombia, lo que en principio significa que se encontraron géneros nuevos para esta área de estudio.
El menor número de organismos en el sistema arroz, puede ser atribuido a las prácticas de manejo del sistema, tal como lo encontraron DRESS & BOERNER (2004), donde las prácticas agrícolas generan un efecto sobre la reproducción y una disminución del alimento, que afecta la dinámica de la población de microartrópodos. La mayor abundancia de isotomidos, se atribuye a que sus especies han sido registradas como de gran adaptabilidad a perturbaciones ocasionadas por prácticas agrícolas y forestales, y que tal como lo mencionan MENDOZA et al. (1999), pueden proliferar en suelos con bajo o alto contenido de materia orgánica. Así tenemos que las especies de isotomidos encontradas en el presente estudio, son cosmopolitas y su presencia puede estar relacionada con las alteraciones provocadas por la actividad agrícola, donde las especies de colémbolos estenotípicas del bosque primario, son desplazadas. MORALES & SARMIENTO (2002), revelan que algunas especies son muy afectadas por la perturbación y otras son beneficiadas por la intervención de los ecosistemas. Esto coincide con lo expuesto por KOGAN (1981) y WOLTERS & EKSCHMITT (1997), quienes indican que en algunos casos la perturbación agrícola es beneficiosa para ciertas especies animales estrategas "r", capaces de usar nuevos recursos disponibles y recolonizar rápidamente el área afectada.
La densidad de colémbolos en la zona de estudio (3.095 a 5.823 Ind/m²), es baja, si se compara con otros registros como el de BANASCO (1985), en Cuba, que obtuvo una densidad media de colémbolos que osciló entre 14.700 y 43.100 Ind/m², en caña de azúcar y plátano, o los resultados encontrados por CHAUVAT et al. (2007), donde la abundancia de colémbolos fue de 9.300 Ind/m² a 58.900 Ind/m², demostrando un incremento en la abundancia, inmediatamente después de la conversión de cultivo a pastos. Contrariamente, nuestras densidades son muy altas si se tienen en cuenta los registros de MENDOZA et al. (1999), en cultivos de maíz en Chiapas (México) o los encontrados por WATANABE & RUAYSOONGNERN (1989), de 195 ind/m² en la Selva Perennifolia del noreste de Tailandia.
Cuando las localidades de colecta presentaron un mayor número de prácticas de manejo, las especies y el número de organismos decreció y la mayor abundancia y densidad correspondió a áreas en reconversión (sistemas silvopastoriles), que presentan una serie de características físicas y químicas que pueden estar influyendo las poblaciones de colémbolos (contenidos de materia orgánica medios y altos (3,7%-5,09%), pH cercanos a la neutralidad, textura franco arcillosa), además, de una menor temperatura y una mayor humedad del suelo, debido a que en los sistemas silvopastoriles el suelo está menos expuesto a altas temperaturas, por la protección brindada por especies forrajeras como la leucaena, y cultivos de pastos como Cynodon nlemfuensis, que hacen las fluctuaciones en la temperatura menos profundas a nivel del suelo, evitando la evaporación del agua, máxime si se tienen en cuenta las altas temperaturas de la región de estudio. Estos resultados son similares a los de GÓMEZ (1998), quien atribuye la variación de las poblaciones de colémbolos al contenido de materia orgánica y humedad del suelo, lo que se hace evidente en nuestro estudio donde el sistema silvopastoril 8 años con materia orgánica (5,09%, clasificado como alto), fue el de mayor abundancia.
El número de colémbolos fue menor en los meses de mayor sequía, donde probablemente como lo registran ARBEA & BLASCO (2001), los organismos no se encuentran en un estado fisiológico activo que permita su captura con los métodos de extracción utilizados. Es de anotar, que la precipitación no es el único limitante para la abundancia de colémbolos, es así, que el menor número de organismos, que se registró en los meses de octubre y noviembre, puede ser también atribuido al aumento en las prácticas de manejo en el cultivo de arroz (aplicación de insecticidas y fungicidas, remoción de la vegetación), además de que en el mes de octubre, en los sistemas silvopastoriles se realizó pastoreo durante 35 días.
Tanto a nivel general como por familias, se observó variación temporal, que puede indicar que existen fluctuaciones estacionales en la tasa reproductiva de algunas especies que modifican la estructura de la comunidad, tal como lo encontraron FRAILE & SERAFINO (1978) y TAKEDA (1987), donde cambios estacionales en las condiciones ambientales, pueden promover explosiones demográficas en los colémbolos y que ARBEA & BLASCO (2001), asocian la humedad y la temperatura, como factores determinantes del hábitat óptimo de los colémbolos, que influyen en la tasa de reproducción, crecimiento de los individuos y en su distribución vertical a lo largo del perfil; es así que GONZÁLEZ et al. (2001), encontraron que la producción de espermatoforos disminuye con la temperatura, y bajo condiciones de altas temperaturas se incrementa el riesgo de depredación, ya que los animales se hacen más vulnerables a sus enemigos debido al elevado número de mudas.
Medición de la diversidad Alfa
La alta diversidad en cuanto a la riqueza de especies en cada uno de los sistemas encontrado por el índice de Margalef, puede ser explicado por el alto número de especies euritípicas encontradas en el estudio y la escasa presencia de especies específicas para cada sistema. La evaluación de la diversidad Alfa mediante curvas de acumulación, donde las curvas de los estimadores no paramétricos tuvieron un comportamiento asintótico, mientras las curvas de las especies representadas sólo en una o dos muestras (uniques, duplicates) tendieron a descender, indicó que el muestreo realizado fue suficiente para hallar el número de especies en cada uno de los sistemas. El índice de Shannon-Weiner, mostró que las especies no se distribuyen de manera proporcional, y existe dominancia de algunas especies, que puede ser explicada por el incremento en la proporción de individuos de la familia Isotomidae en todos los sistemas, sumado a que los valores son variables en otro estudios y sus comparaciones son difíciles por las diferencias en el método de muestreo y extracción de la fauna.
El índice de Simpson, interpretado según CAVIEDES (1999), también presentó una alta dominancia, que es favorecida por un elevado número de organismos isotomidos, entre los cuales pueden presentarse especies oportunistas, que se ven favorecidas en el cultivo y en los sistemas silvopastoriles, como son las morfoespecies: Microisotoma sp., F. americanus, Isotomiella sp., Proisotoma sp. y Folsomia sp.
Medición de la diversidad Beta
El dendrograma correspondiente a la clasificación jerárquica de los sistemas construido con base al índice de Jaccard, mostró dos agrupamientos: un grupo correspondiente a los tres sistemas silvopastoriles y el otro al cultivo arroz, varias de las especies que comparten todos los sistemas, son isotomides y algunos Entomobrydes, que se consideran cosmopolitas. La presencia exclusiva de especies como Isotoma sp., a un ecosistema, sugiere sensibilidad a la perturbación ocasionada por el aprovechamiento agrícola, tal como lo ha encontrado CUTZ (2003). La similitud encontrada entre sistemas, concuerda con los resultados de CHAUVAT et al. (2007), quienes reportan que dos poblaciones pueden compartir un gran número de especies, pero cada especie tiene una representación diferente en el total de biomasa de la comunidad y por lo tanto las comunidades son disímiles, presentando una tendencia a incrementar la similaridad con la edad del pastizal.
En el Análisis Factorial de Correspondencia, se puede observar que de acuerdo a sus poblaciones de colémbolos, se distinguen dos grupos que corresponden con una cobertura vegetal y un grado de perturbación del medio natural diferente. Por un lado, están los sistemas con mayor cobertura vegetal (silvopastoriles de 8 y 3 años), que representan una similaridad faunistica muy alta; y por otro lado, comunidades más degradadas, como el arroz o en proceso temprano de reconversión (silvopastoril 7 meses), que poseen una cobertura vegetal escasa. Los resultados del análisis factorial de correspondencia, efectuado sobre las morfoespecies presentes en los sistemas estudiados, muestran que los dos primeros ejes del análisis, explican el 85% del total de la varianza, y reflejan lo más importante de la estructura de las poblaciones de colémbolos. El eje 1 (62,75% de la varianza), separa los sistemas más similares, con igual número de especies en común (silvopastoril 3 y 8 años), en los valores más positivos del eje 1. En los valores más negativos del eje 1, aparecen los sistemas arroz y silvopastoril 7 meses, junto con las especies características de estos biotopos.
Esta división entre sistemas, puede deberse a los estados sucesionales, donde las especies mejor adaptadas a suplir recursos locales (isotomidos), podrían dividir el espectro de recursos disponibles de una manera jerárquica, y forzar a otras especies a roles secundarios en la comunidad, tal como lo registran VEGTER et al. (1988). Estos resultados son concordantes con los de SIEMANN et al. (1999), en la dinámica sucesional de la diversidad de plantas y artrópodos en una pradera norteamericana y que encuentran una clara correlación entre la diversidad de ambas comunidades, la cual MORALES & SARMIENTO (2002), interpretan en función que un mayor número de especies vegetales representan una mayor variedad de recursos y de hábitat para los artrópodos, y que LONGCORE (2003) encontró que el lapso de tiempo de revegetación, tiene efecto sobre la estructura de la comunidad de artrópodos, pero no es gradual el retorno a una comunidad natural, exhibiendo un menor número de artrópodos nativos.
HAZRA & CHOUDHURI (1993), encontraron que la densidad y número de especies de estos organismos disminuye con la pérdida de materia orgánica, en hábitat naturales y sitios cultivados, está perdida de materia orgánica en suelos agrícolas, CURRY & GOOD (1992), la asocian principalmente al manejo de las parcelas y periodos de monocultivo continuo, lo cual es evidente en el cultivo de arroz, donde además se presentan contenidos de materia orgánica bajos. Las especies que tienen rangos de tolerancia más restringidos, como es el caso de las morfoespecies Xenilla sp., Cyphoderus sp., Mesaphorura sp., y que se presentaron como exclusivas para los sistemas silvopastoriles, y Lepidocyrtus sp., que fue encontrado en arroz, al igual que Isotoma sp., exclusiva para el sistema silvopastoril 7 meses, determinaron las diferencias entre sistemas.
DUNGER et al. (2004), han mostrado la importancia de los Onychiuridea como indicadores, en especial algunas especies del género Mesaphorura, indicadoras de suelos en zonas de baja precipitación, sin exceso de humedad y posibles indicadores de fertilidad. Estos organismos, también son pioneros en sucesiones, sobre suelos recultivados, estas observaciones, pueden validar la presencia de Mesaphorura en los sistemas silvopastoriles. Los miembros de la familia Cyphoderidae, son comensales facultativos u obligados a insectos sociales, según los registros de VÁSQUEZ & PALACIOS (1990), además según PALACIOS et al. (2000), su distribución está limitada por la propia distribución de sus comensales (himenópteros y termitas), razones por las cuales se puede justificar la aparición de esta morfoespecie sólo en los sistemas silvopastoriles, que presentaron una mayor abundancia de insectos sociales asociados (datos no publicados).
En cuanto a los Entomobriomorfos, los géneros Entomobrya, Heteromurus, Lepidocyrtus y Seira, son especies epigeas, que presentan adaptaciones morfológicas para reducir la transpiración de la superficie del cuerpo (escamas, cubierta densa de pelos, intensa pigmentación), que las favorecen de condiciones extremas de humedad y temperatura y que según BILDE & TOFT (2000), algunas especies del género Lepidocyrtus, utilizan estrategias que le sirven como defensa contra los predadores (escamas o sustancias fenólicas). Estas características de la familia Entomobridae, en especial del género Lepidocyrtus, puede explicar la presencia de este organismo sólo en el cultivo de arroz, a pesar de ser un sistema con una gran cantidad de prácticas de manejo, que afectan a los colémbolos.
AGRADECIMIENTOSAl Comité Central de Investigaciones - Universidad del Tolima y a la Corporación Colombiana de Investigaciones Agropecuaria –CORPOICA–, por la financiación de este trabajo. A Roberto Mejía, propietario hacienda El Chaco (Piedras - Tolima), José Guadalupe Palacios Vargas (Laboratorio de Ecología y Sistemática de Microartrópodos - UNAM), Claudia Marcela Ospina y Jairo Rodríguez Chalarca (CIAT). A Jaime Vicente Estévez y Luis Alfredo Lozano, por sus valiosas sugerencias en el análisis de datos.
ARBEA, J. & BLASCO, J., 2001.- Ecología de los Colémbolos (Hexapoda, Collembola) en los Monegros (Zaragoza, España). Aracnet, 7 (28): 35-48. [ Links ]
BANASCO, J., 1985.- Colémbolos en el complejo de microartrópodos de los suelos cultivados de Cuba: Tesis, Universidad de Moscú, Departamento de Ciencias Biológicas, Moscú. [ Links ]
BILDE, J. & TOFT, S., 2000.- The value of collembolan from agricultural soils as food for a generalist predator. Journal of Applied Ecology, 37: 672-683. [ Links ]
CASSAGNE, N.; BAL-SERIN, M.; GERS, C. & GAUQUELIN, T., 2004.- Changes in humus properties and collembolan communities following the replanting of beech forests with spruce. Pedobiología, 48: 267-276. [ Links ]
CAVIEDES, B., 1999.- Manual de Métodos y Procedimientos Estadísticos. Universidad Jorge Tadeo Lozano. [ Links ]
CHAUVAT, M.; WOLTERS, V. & DAUBER, J., 2007.- Response of collembolan communities to land-use change and grassland succession. Ecography, 30: 183-192. [ Links ]
CHRISTIANSEN, K. & BELLINGER, P., 1980.- The Collembola of North America, North of the Rio Grande Iowa. Grinnell College, 3: 785-1042. [ Links ]
________., 1981.- The Collembola of North America, North of the Rio Grande Iowa. Grinnell College, 4: 1043-1322. [ Links ]
CURRY, J. & GOOD, J., 1992.- Soil faunal degradation and restoration. Soil Science, 17: 171-215. [ Links ]
CUTZ, L., 2003.- Colémbolos edáficos de dos agrosistemas de San Salvador, Hidalgo: Tesis, Universidad Nacional Autónoma de México, Facultad de Ciencias, México. [ Links ]
CUTZ, L.; PALACIOS, J. & VÁZQUEZ, M., 2003.- Comparación de algunos aspectos ecológicos de Collembola en cuatro asociaciones vegetales de Noh-Bec, Quintana Roo, México. Folia Entomológica Mexicana, 42 (1): 91-101. [ Links ]
DRESS, W. & BOERNER, R., 2004.- Patterns of microartropod abundance in oak-hickory forest ecosystems in relation to prescribed fire and landscape position. Pedobiología, 48: 1-8. [ Links ]
DUNGER, W.; SCULZ, H.; ZIMDARS, B. & HOHBERG, K., 2004.- Changes in collembolan species composition in Easterm German mine sities over fifty years of primary succession. Pedobiología, 48: 503-517. [ Links ]
EATON, R.; BARBERCHCK, M.; BUFORD, M. & SMITH, W., 2004.- Effects of organic matter removal, soil compaction, and vegetation control on Collembolan populations. Pedobiología, 48 (2): 121-128. [ Links ]
FRAILE, M. & SERAFINO, A., 1978.- Variaciones mensuales en la densidad de microartrópodos edáficos en un cafetal de Costa Rica. Rev. Biología Tropical, 26: 291-301. [ Links ]
GÓMEZ, J., 1998.- Ecología de collembola (Hexapoda: Apterygota) de Chamela, Jalisco, México: Tesis, Universidad Nacional Autónoma de México, Facultad de Ciencias, División de Estudios de Postgrado, México. [ Links ]
GONZÁLEZ, V.; DIAZ, M. & PRIETO, D. 2001.- Comunidades de la mesofauna edáfica en el ciclo de caña-planta (Saccharum spp.). Revista Biología, 15 (2): 1-6. [ Links ]
HAZRA, A. & CHOUDHURI, D., 1993.- A study of collembola communities in cultivatedand uncultivated sities of West Bengal in relation to threemajor soil factors. Rev. Ecol. Biol. Sol., 20: 385-401. [ Links ]
JANSSENS, F., 2003.- Checklist of the collembola: Families. [On Line]. Belgium. University of Antwerp (RUCA). [Citado en octubre 16 de 2004]. Available from internet: http://www.collembola.org/taxa/collembo.htm [ Links ]
KOGAN, M., 1981.- Dynamics of insect adaptacions to soybeans: impact of integrated pest management. Environmental Entomology, 10 (3): 663-670. [ Links ]
LONGCORE, T., 2003.- Terrestrial arthropods as indicators of ecological restoration success in coastal sage scrub (California, USA). Restoration Ecology, 11 (4): 397-409. [ Links ]
MENDOZA, S.; VILLALOBOS, F.; RUIZ, L. & CASTRO, A., 1999.- Patrones ecológicos de los Colémbolos en el cultivo de maíz en Balun Canal, Chiapas, México. Acta Zool. Mex., 78: 83-101. [ Links ]
MORALES, J. & SARMIENTO, L., 2002.- Dynamics of soil macroinvertebrates and its relationships to vegetation in a secondary sucesión of the Venezuelan paramo. Ecotropicos, 15 (1): 99-110. [ Links ]
MUTT, A., 1987.- Collembola from two localities near Buenaventura, Colombia. Journal of the Kansas Entomological Society, 60 (3): 364-379. [ Links ]
OSPINA, C.; RODRÍGUEZ, J.; MOJOCOA, M.; SERNA, F. & PECK, D., 2004.- Colémbolos asociados a cultivos de maíz y algodón en el Valle del Cauca, Colombia: 57 (en) Resúmenes XXXI Congreso SOCOLEN: Sistemática y Taxonomía. [ Links ]
OSPINA, C.; SERNA, F.; PEÑARANDA, M. & SERNA, S., 2003.- Colémbolos asociados con cultivos de pastos en tres zonas de vida de Holdridge en Antioquia (Colombia). Agronomía Colombiana, 21(3): 129-141. [ Links ]
PALACIOS-VARGAS, J., 1990.- Diagnosis y clave para determinar familias de los collembola de la región neotropical: 1-15 (en) Manuales y guías para el estudio de microartrópodos. UNAM, México. [ Links ]
________., 1991.- Introducción a los insectos sin alas (Protura, Diplura, Collembola, Thysanura): 1-23 (en) Manuales y guías para el estudio de microartrópodos. UNAM, México. [ Links ]
PALACIOS-VARGAS, J.; CASTAÑO, G. & MEJÍA B., 2000.- Collembola: 249-273 (en) LLORENTE J., GONZÁLEZ E. & PAPAYERO N. Biodiversidad, taxonomía y biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento. UNAM, México. [ Links ]
PEÑARANDA, M.; SERNA, F. & PALACIOS, J., 2004.- Colémbolos (Hexapoda: Collembola): 61 (en) Resúmenes XXXI Congreso SOCOLEN: Sistemática y Taxonomía. [ Links ]
SIEMANN, E.; HAARSTAD, J. & TILMAN, D., 1999.- Dynamics of plant and arthropod diversity during ald field succession. Ecography, 22: 406-414. [ Links ]
TAKEDA, H., 1987.- Dynamics and maintenance of collembolan community structure in a forest soil system. Res. Pop. Ecol., 29: 291-346. [ Links ]
URIBE, F., 2000.- Revalorización de la investigación hacia la innovación en sistemas de producción tropical: 8 (en) Memorias Seminario Internacional sobre Modelos y Metodologías de Investigación en Sistemas Agroforestales. [ Links ]
VÁZQUEZ, M. & PALACIOS, J., 1990.- Nuevos registros y aspectos biogeográficos de los colémbolos de la Sierra de la Laguna, México. Folia Entomologica Mexicana, 78: 5-22. [ Links ]
VEGTER, E.; ERNSTING, G. & JOOSSE, G., 1988.- Community structure, distribution and population dynamics of entomobrydae (collembola). Journal of Animal Ecology, 57: 971-981. [ Links ]
WATANABE, H. & RUAYSOONGNERN, S., 1989.- Estimation of arboreal arthropod density in a dry evergreen forest in Northeastrrn Thailand. Tropical Ecology, 5: 151-158. [ Links ]
WOLTERS, V. & EKSCHMITT, K., 1997.- Gastropods, Isopods; Diplopods, and Chilopods: Neglected Groups of the decomposer food. Fauna in Soil Ecosystems, 1: 265-306. [ Links ]

