










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.14 n.2 Manizales jul./dic. 2010
PARTE I: REGIONES CORPORALES, ORGANIZACIÓN ANATÓMICA E INTEGUMENTO.
HISTOLOGY OF APIS MELLIFERA LINNAEUS, 1758 (HYMENOPTERA) AS AN ENTOMOLOGICAL CONTRIBUTION.
PART I: BODY REGIONS, ANATOMICAL ORGANIZATION AND INTEGUMENT.
* FR: 2-III-2010. FA: 22-VI-2010
1 Director Grupo de Investigaciones Entomológicas y Ambientales –GENA–. Profesor Asociado UIS. Calle 9 con carrera 27, Edificio Camilo Torres. Consultor Corporación CTAS. Consultor FUSDECOL. E-mail: alfvillalmo@gmail.com
2 Biólogo UIS. Estudiante de Maestría en Entomología, Universidad Nacional de Colombia. E-mail: jcagudelo@gmail.com
3 Profesor Asociado UIS. Director Laboratorio de Histotecnia, Universidad Industrial de Santander. Calle 9 con carrera 27, Edificio Camilo Torres. E-mail: darrietadap@gmail.com
Se realizaron cortes histológicos en planos horizontal, vertical y sagital, usando cabezas y protórax de abeja, con el objetivo de aportar material docente a la asignatura: Electiva Profesional I – Entomología Avanzada. Las láminas fueron seleccionadas por su calidad y se escogieron las mejores para tomar fotos digitales. Se señalaron estructuras y detalles importantes, con los cuales se realizó el presente documento.
Palabras clave: docencia, epidermis, histología, insectos.
AbstractHistological cuts were made in transverse, vertical and sagittal planes, using bees' heads and prothorax with the purpose of contribute with teaching material to the subject: Professional Elective I – Advanced Entomology. The prints were selected because of their quality and the best ones were chosen to be digitally photographed. Structures and important details were pointed out which made possible the making of this document.
Key words: teaching, epidermis, histology, insects.
En entomología, el conocimiento de la morfología del insecto es muy importante para comprender su origen, fisiología, taxonomía, evolución, comportamiento, etc., razón por la cual, dicho estudio es ampliamente utilizado. Sin embargo, la anatomía y la histología se estudian sólo a nivel teórico o se realizan algunas pocas prácticas puntuales, creando serios vacíos cuando se pretende profundizar en temas específicos. Está claro que existen gráficos, esquemas y dibujos detallados, que a pesar de estar a blanco y negro (y tener cierta antigüedad), fueron realizados con calidad y precisión, y son utilizados como textos de apoyo en procesos de disección e interpretación de la anatomía de los insectos. Algunos de estos esquemas pueden ser encontrados en SNODGRASS (1935), WIGGLESWORTH (1956), ROSS (1967), CORONADO & MÁRQUEZ (1976), BORROR et al. (1989), DE LA FUENTE (1994) y OSUNA (1995).
También es posible encontrar fotografías y descripciones histológicas de gran calidad y con un excelente contenido. Algunos son el resultado de disecciones y montajes in toto como los que se observan en SÁNCHEZ et al. (2000) y VILLALOBOS et al. (2009), en otros casos son citologías como las de BRUN et al. (1995) y CASELÍN et al. (2008), cortes para microscopía óptica como en AYALI et al. (2002) y NICULITA et al. (2007), o imágenes de microscopía electrónica como en HARTFELDER (2000), SÁNCHEZ et al. (2000) y CURRIE et al. (2006).
El objetivo de este trabajo fue establecer una metodología para realizar montajes permanentes de tejidos de insectos bajo las condiciones del Laboratorio de Histotecnia-UIS, generando material didáctico que complemente el estudio de la anatomía y la histología de insectos, y finalmente, realizar breves descripciones de los cortes obtenidos usando la literatura disponible.
TÉCNICAS HISTOLÓGICASLas técnicas histológicas usadas en entomología van desde la observación macroscópica de órganos y estructuras, hasta el reconocimiento de modificaciones celulares y estudio de cromosomas politénicos. En general, estas técnicas se dividen en:
Montajes in toto. Se usa la estructura completa y se realizan las observaciones directas, haciendo tinción con productos poco específicos, como azul de metileno. Algunas aplicaciones de esta técnica se observan en el análisis de estructuras en ontogenia pupal, seguimiento de modificaciones tegumentarias, estructura de gónadas y tubo digestivo (SÁNCHEZ et al., 2000; VILLALOBOS et al., 2009). Los montajes in toto son usados en procesos de diafanización para analizar genitalia de lepidópteros, coleópteros y hemípteros (WOODRUF, 1973; YÉLAMOS, 1990; HERNÁNDEZ & ORTUÑO, 1992; BAHILLO DE LA PUEBLA, 1996; GUTIÉRREZ-S., 1999; CASTAÑEDA et al., 2007; MURILLO, 2007).
Cortes histológicos. Son usados para el estudio detallado de tejidos, y pueden ser seriados o no, dependiendo de los objetivos del trabajo. En el presente documento se aplica esta técnica, la cual presenta un mayor grado de complejidad y requiere gran experiencia en la elaboración de los montajes e interpretación de los mismos. Como los insectos presentan un exoesqueleto rígido y un interior con órganos blandos y delicados, se plantean desafíos en el momento de elaborar los cortes, pues se podrían deteriorar los órganos internos o fracturar el integumento. La elaboración de cortes histológicos tiene cinco pasos básicos:
1. Fijación. Consiste en someter los tejidos a sustancias que impidan la variación sustancial tanto de la forma como de la constitución celular. En entomología son usados por lo general fijadores como la solución Carnoy, la solución Bouin, el FAA (usado en plantas pero también en insectos) y el FAATD, este último usado en el presente trabajo. Es importante ajustar la información encontrada en otras experiencias, a las necesidades propias y especialmente a las dimensiones de la muestra.
2. Deshidratación. Consiste en reducir el nivel de agua de la muestra a través del paso por una serie de alcoholes que van desde etanol 70% hasta etanol 100%, mezcla de etanol-xilol 50-50 y xilol 100%. Este proceso garantiza que el fijador sea reemplazado por xilol que por su alta volatilidad facilita el paso siguiente.
3. Impregnación. Consiste en incorporar a la muestra algún tipo de medio sólido o semisólido que permita realizar un buen corte. Para este fin, pueden ser usados geles, plástico, combinaciones de gel-parafina o sencillamente parafina, que por su economía y fácil manipulación suple las necesidades de cualquier laboratorio de histotecnia. En el caso de los insectos, este paso debe realizarse con especial cuidado, puesto que la naturaleza impermeable del exoesqueleto genera problemas en la entrada de parafina. Así, la entrada está determinada por el nivel de esclerotización del segmento a tratar, de tal manera que en el abdomen se facilita la entrada de parafina mientras que en la cabeza el proceso se hace más lento.
4. Corte. De manera tradicional se han usado micrótomos con cuchillas de acero. También existe el criostato, que corta muestras que han sido congeladas con nitrógeno líquido.
5. Tinción. La coloración celular y de tejidos es una combinación de fenómenos físicos y químicos. Los físicos son absorción, capilaridad y ósmosis, y los químicos están definidos por la afinidad de colorantes básicos por componentes ácidos y viceversa. Las sustancias utilizadas son muy diversas, pero en la UIS se usa frecuentemente hematoxilina, eosina, azul de metileno, azul de toluidina y Sudan III, entre otras.
MATERIALES Y MÉTODOSPara la realización de este trabajo, se contó con un micrótomo marca American Optical Corporation, un baño maría marca Lipshaw, cuchillas desechables de acero inoxidable, un dispensador de parafina Tissue TeK, parafina en lentejas Paraplast, láminas portaobjetos y cubreobjetos, canastillas metálicas, cajas de madera, una batería de alcoholes, albúmina de Mayer, bálsamo del Canadá y una batería de tinción de hematoxilina-eosina (Figura 1).

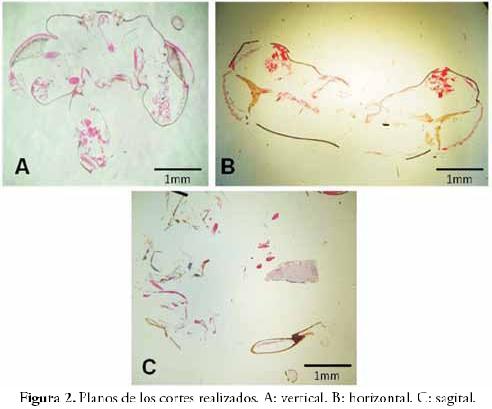
Se realizaron cortes histológicos seriados de cabeza y protórax de A. mellifera, en planos vertical, horizontal y sagital. En la Figura 2 se observan ejemplos de los tipos de cortes realizados. Vertical: (a la izquierda) se identifican los ocelos en la parte superior, los ojos a los lados y componentes del aparato maxilolabial abajo. Horizontal: (a la derecha) se reconocen los ojos en posición lateral. Sagital: (abajo) se observa el cerebro, apéndices cefálicos y partes del cuello (este plano presentó el mayor nivel de fragmentación). Se ensayó con tres tiempos de ablandamiento con KOH (10, 15 y 30 minutos), tres soluciones de fijación (FAA, FAADMSO y FAATD) y cuatro tiempos de fijación (12, 24, 48 y 72 horas). Además, se evaluaron tres tiempos de impregnación en parafina (24, 48 y 72 horas).
RESULTADOS
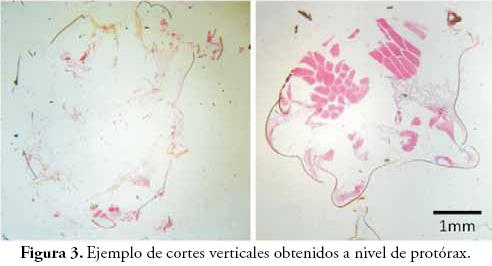
Se realizaron más de mil cortes, que fueron organizados en 310 montajes permanentes, de los cuales se seleccionaron 152 láminas por la calidad del montaje. En la Figura 3, se observa un corte con evidente deterioro (izquierda) y uno de los cortes seleccionados por buena calidad y poca fragmentación (derecha), e incluso se observan numerosos detalles. A los mejores montajes se les hicieron fotos digitales con una cámara SONY Cybershot adaptada a un microscopio Olympus, obteniéndose 191 fotografías que fueron analizadas y procesadas con la ayuda de los programas Microsoft Office Picture Manager, CorelDraw Graphics Suite X3 y Adobe Photoshop CS2.
Los ensayos demuestran que el uso de KOH permite lograr mejores resultados, pero que más de 15 minutos provocan notable deterioro de los tejidos. Además, se concluyó que debido a que la impregnación de parafina es uno de los pasos que presenta mayores inconvenientes por la impermeabilidad y grosor del integumento (especialmente la cabeza de los insectos), el uso de FAATD durante más de 48 horas permitió obtener los mejores montajes, puesto que se aumenta la entrada de parafina. También se concluyó que el mayor tiempo de impregnación en parafina determinó la mejor calidad de los montajes.
Con el propósito de comprender e interpretar mejor los resultados, la información está organizada en siete secciones de las cuales se presentan las primeras tres en el presente documento. En cada sección se muestran algunas de las imágenes digitales obtenidas, se señalan algunas estructuras y se hacen breves descripciones.
I. Regiones corporales y organización anatómica
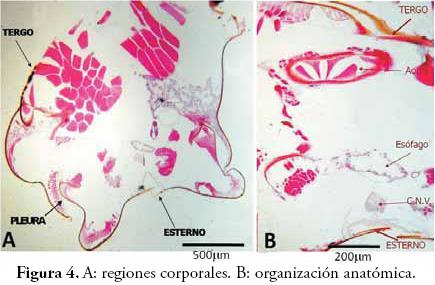
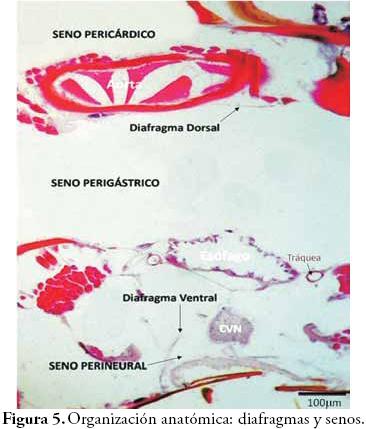
En la Figura 4A, se observan las regiones corporales de la siguiente manera: el tergo en posición dorsal y que se reconoce como la placa más grande, el esterno en posición ventral es la placa más pequeña, y las pleuras se observan como zonas laterales plegadas. En la Figura 4B, se observa la organización anatómica de los insectos (similar a la de anélidos y artrópodos en general). Dorsalmente se encuentra el sistema circulatorio constituido por el vaso aórtico, en el centro está el sistema digestivo representado por un corte de esófago, y ventralmente el sistema nervioso específicamente el cordón nervioso ventral (CNV). En la Figura 5 se indican los diafragmas dorsal y ventral que separan la cavidad corporal de los insectos en sus tres senos o lagunas (lo que define al sistema circulatorio como abierto o lagunar). Dorsalmente se señala el seno pericárdico, hacia el centro se observa el seno perigástrico y ventralmente el seno perineural.
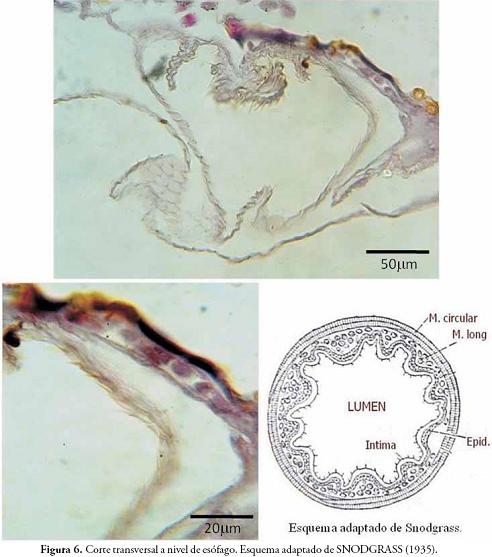
En la Figura 6 se observan detalles de un corte transversal de esófago, donde se evidencia una capa quitinosa interna, ubicada hacia la luz del tubo, lo cual expone su origen tegumentario, incluso se notan las formas hexagonales características de las deposición de quitina en lamelas. En la misma figura, se observan cortes transversales de la musculatura oblicua y circular del tubo digestivo.
II. Integumento
El integumento de los insectos, y de los artrópodos en general, presenta dos componentes básicos: epidermis y cutícula (Figura 7). El primero es una sola capa de células con apariencia sincitial, y el segundo se encuentra formado por quitina, varios tipos de proteínas, componentes fenólicos y pigmentos, en combinaciones que varían según la región corporal y capa cuticular (epicutícula, exocutícula y endocutícula). La cutícula tiene un grosor entre 100 y 300 micrometros, y es secretada por la epidermis siguiendo una disposición en lamelas según el modelo parabólico o perfil de equilibrio (Figura 7) (RESH & CARDÉ, 2003).
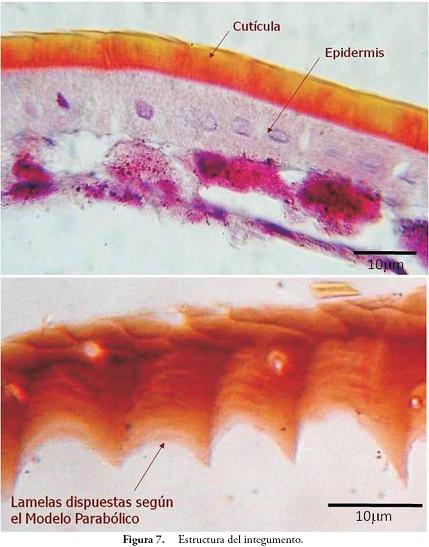
Al integumento se le considera un tejido dinámico con responsabilidades diversas como la reelaboración de la cutícula cada vez que un insecto hace ecdisis, la participación en procesos glandulares y la conformación de varios órganos de los sentidos (RESH & CARDÉ, 2003). La importancia del integumento también radica en ser el límite protector, evitando la entrada de agentes patógenos, e impidiendo la salida de agua. Además, presenta numerosas modificaciones que le aportan al insecto sitios de inserción muscular, puntos de articulación, gran rigidez y flexibilidad, etc.
III. Modificaciones integumentarias
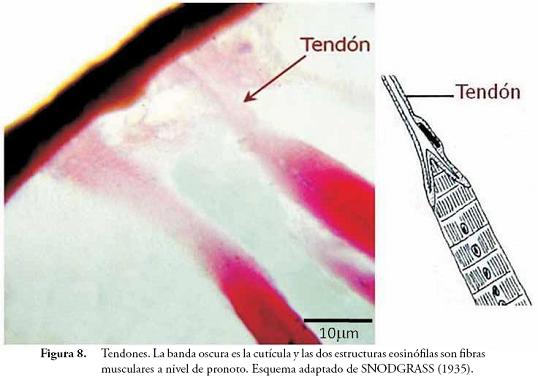
Dentro de las modificaciones integumentarias de los insectos existen las siguientes clases: apodemas (fragmas y tendones), procesos externos (cuernos, setas, espinas, nódulos, estrías, etc.), suturas (líneas, pseudosuturas y suturas verdaderas) y articulaciones (membranas articulares y articulaciones específicas). La mayoría de estas estructuras son observadas con facilidad realizando disecciones muy sencillas, sin embargo otras, como los tendones, sólo se aprecian si se hacen cortes microscópicos. En las láminas permanentes de A. mellifera, se pudieron observar algunas de estas modificaciones.
Tendones (Figura 8): corresponde a una clase de modificación interna que consiste en células epidérmicas que se alargan para unirse a cada una de las fibrillas musculares, las cuales también presentan sus propias modificaciones para acoplarse, llamadas tonofibrillas.
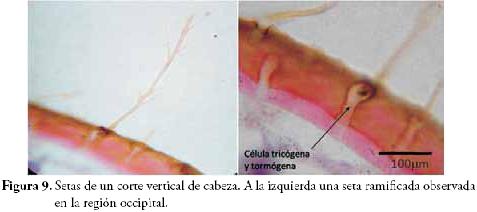
Setas (Figura 9): son modificaciones externas donde intervienen dos células epidérmicas. La tricógena, forma el componente filiforme que sobresale, y la tormógena, crea el espacio para que la primera se mueva libremente y cumpla con sus funciones sensitivas sin riesgo de fracturarse. En algunos casos, toda la estructura es ocupada por una célula secretora de origen mesodérmico, y se convierte en una seta glandular.
Sensilas (Figura 10): en los cortes de antenas y palpos de A. mellifera se identificaron dos tipos de sensilas. Las setáceas que tienen forma de pelo corto y se encuentran relacionadas con la recepción de estímulos mecánicos y químicos, y las placódeas que tienen forma de copa y cumplen funciones de recepción de estímulos térmicos y químicos.
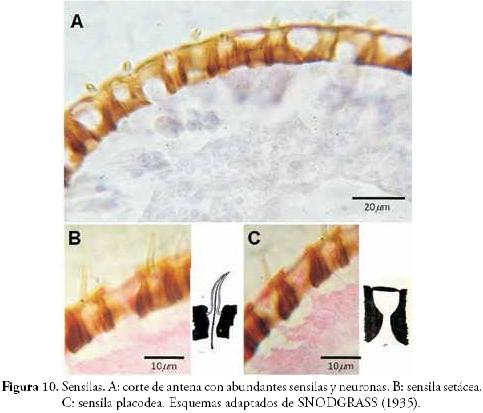
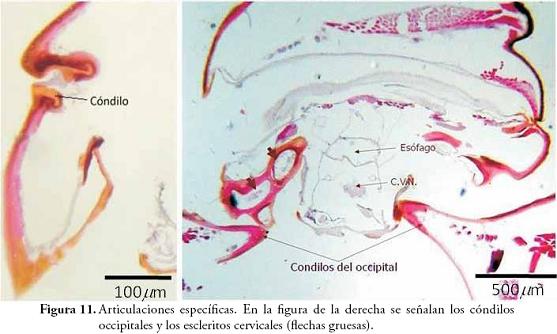
Articulaciones (Figura 11): se definen como modificaciones externas que se convierten en espacios de inflexión y apoyo. Se clasifican en dos grupos, las membranas articulares que corresponden a zonas originalmente no esclerotizadas (por ejemplo: las membranas intersegmentarias y las pleuras), y las articulaciones específicas (monocondílicas y dicondílicas) que se caracterizan por ser puntos de contacto entre dos escleritos. En la Figura 11 se observa el detalle de una articulación específica (a la izquierda) donde se hace evidente la presencia de un cóndilo mandibular, y un corte a nivel de cuello (a la derecha) mostrando la articulación dicondílica que une la cabeza con el protórax.
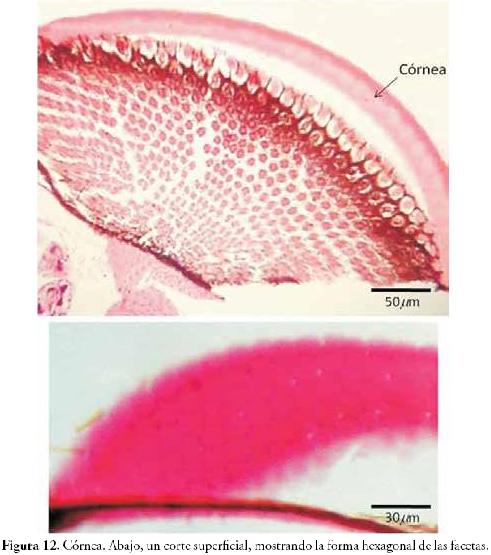
Córnea (Figura 12): modificación netamente cuticular que corresponde a la parte más externa del ojo compuesto. Se observa al microscopio como una capa gruesa y eosinófila. La modificación forma un lente hexagonal de superficie convexa llamado faceta (Figura 12, foto de abajo). Las células epidérmicas (debajo de la córnea) se modifican y especializan para formar complejas estructuras fotorreceptoras llamadas ommatidas.
AGRADECIMIENTOS
Los autores desean agradecer a todas y cada una de las personas que hicieron posible este artículo, especialmente a nuestras familias y amigos por su paciencia en los momentos en que no estuvimos presentes por estar realizando esta investigación. A nuestros amigos Wilson Medina (q.e.p.d.), Jorge Villamizar Cobos, Arnaldo Báez, Jorge Niño Rondón, Adolfo Hernández García e Inés Johanna Gómez por su colaboración y acompañamiento permanente. A los doctores Jaime Alberto Camacho Pico, Sergio Isnardo Muñoz Villareal y Óscar Gualdrón por su apoyo para asistir al XXXV Congreso de la Sociedad Colombiana de Entomología en la ciudad de Cali, donde se presentaron avances de este trabajo.
AYALI, A.; ZILBERSTEIN, Y. & COHEN, N., 2002.- The locust frontal ganglion: a central pattern generator network controlling foregut rhythmic motor patterns. Journal of Experimental Biology, 205: 2825-2832. [ Links ]
BAHILLO DE LA PUEBLA, P., 1996.- Sobre la recolección, tratamiento y conservación de los cerambícidos. Bol. SEA, 14: 55-62. [ Links ]
BORROR, D.J.; TRIPLEHORN, C.A. & JOHNSON, C.M., 1989.- An Introduction to the Study of Insects. Saunders College Publishing, USA. 875p. [ Links ]
BRUN, L.O.; STUART, J.; GAUDICHON, V.; ARONSTEIN, K. & FRENCH-C., R.H., 1995.- Functional haplodiploidy: A mechanism for the spread of insecticide resistance in an important international insect pest. Proc. Natl. Acad. Sci., 92: 9861-9865. [ Links ]
CASELÍN-C., S.; LLANDERAL-C., C.; RAMÍREZ-C., A.; SOTO-H., M. & MÉNDEZ-M., J.T., 2008.- Caracterización morfológica de hemocitos de la hembra de Dactylopius coccus (Hemiptera: Coccoidea). Agrociencia, 42: 349-355. [ Links ]
CASTAÑEDA-V., A.; VALDEZ-C., J.; EQUIHUA-M., A.; GONZÁLEZ-H., H.; ROMERO-N., J.; SOLÍS-A., J.F. & RAMÍREZ-A., S., 2007.- Genitalia de tres especies de Heilipus (Coleoptera: Curculionidae) que dañan frutos de aguacate (Persea americana) en México y Costa Rica. Neotropical Entomology, 36 (6): 914-918. [ Links ]
CORONADO, P. & MÁRQUEZ, A., 1976.- Introducción a la Entomología, Morfología y Taxonomía de los insectos. Limusa-Wiley, México. 282p. [ Links ]
CURRIE, C.R.; POULSEN, M.; MENDENHALL, J.; BOOMSMA, J.J. & BILLEN J., 2006.- Coevolved crypts and exocrine glands support mutualistic bacteria in fungus-growing ants. Science, 311: 81-83. [ Links ]
DE LA FUENTE, J.A., 1994.- Zoología de Artrópodos. Interamericana / McGraw-Hill, Madrid, España. 805p. [ Links ]
GUTIÉRREZ-S., F., 1999.- Valor de la genitalia como carácter taxonómico en la identificación de géneros de Apiomerini (Hemiptera: Reduviidae). Bioagro, 11 (1): 29-39. [ Links ]
HARTFELDER, K., 2000.- Insect juvenile hormone: from "status quo" to high society. Braz. J. Med. Biol. Res., 33 (2): 157-177. [ Links ]
HERNÁNDEZ, J.M. & ORTUÑO, V.M., 1992.- Estudio de la genitalia femenina en Iberodorcadzon (Breuning, 1943) y comentarios sobre su valor taxonómico (Coleoptera: Cerambycidae). Graellsia, 48: 91-97. [ Links ]
MURILLO, L.R., 2007.- Un método para la identificación de tres especies crípticas de Protesilaus (Lepidóptera: Papilionidae) del sur de Brasil, basado en su morfología genital. Rev. Biol. Trop., 55 (2): 665-671. [ Links ]
NICULITA, H.; BILLEN, J. & KELLER, L., 2007.- Comparative morphology of cephalic exocrine glands among castes of the black ant Lasius niger. Arthropod Structure y Development, 36: 135-141. [ Links ]
OSUNA, E., 1995.- Morfología del exoesqueleto de los insectos. Volumen I: Origen y evolución, el exoesqueleto y Volumen II: Trabajo de laboratorio. Anauco Editores, Universidad Central de Venezuela, Caracas, Venezuela. 475p. [ Links ]
RESH, V. & CARDÉ, R., 2003.- Enclyclopedia of Insects. Elsevier Science, USA. 1295p. [ Links ]
ROSS, H., 1967.- A text book of Entomology. John Wiley, New York, USA. 539p. [ Links ]
SÁNCHEZ, P.A.; SÁNCHEZ, F.; CAETANO, F.H. & JAFFÉ, K., 2000.- El tubo digestivo en adultos de Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae): Morfología y ultraestructura. Bol. Entomol. Venez., 15 (2): 195-216. [ Links ]
SNODGRASS, R.E., 1935.- Principles of Insect Morphology. McGraw-Hill, London, England. 667p. [ Links ]
VILLALOBOS-M., A.; AGUDELO-M., J.C.; BARAJAS-S. A.F.; HERRERA-Q., S. & CÉSPEDES, J.C., 2009.- Cambios morfológicos durante la ontogenia del estado pupal de Melanoloma viatrix (Diptera: Richardiidae). Rev. Col. Entomología, 35 (1): 98-100. [ Links ]
WIGGLESWORTH, V.B., 1956.- Insect physiology. 5th Ed. Methuen & Co., London, England. 134p. [ Links ]
WOODRUF, R.E., 1973.- Arthropods of Florida and Neighboring Lands Areas. Volume 8: The Scarab beetles of Florida (Coleoptera: Scarabaeidae Part I). Florida Departament of Agriculture and Consumer Services, Gainsville, Florida, USA. 220p. [ Links ]
YÉLAMOS, T., 1990.- Preparación de genitalias de insectos. Bol. SEA, 8: 35-36. [ Links ]

