










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.15 no.1 Manizales Jan./June 2011
MICROBIOLOGICAL AND MOLECULAR EVALUATION OF MONILIOPHTHORA PERNICIOSA (AGARICALES: MARASMIACEAE)
* FR: 3-IX-2010. FA: 28-V-2010.
1 Grupo de Investigación Genética, Biodiversidad y Fitomejoramiento –GEBIOME–, Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Caldas, Manizales, Caldas, Colombia. E-mail: german.lopez@ucaldas.edu.co; fredy.rivera@ucaldas.edu.co
Moniliophthora perniciosa presenta discrepancias en su taxonomía y es el agente causal de la escoba de bruja del cacao, que en Colombia es la segunda enfermedad después de la moniliasis (Moniliophthora roreri). En este estudio se analizaron 12 muestras del patógeno, obtenidas de diferentes materiales de cacao de la Granja Casa Luker S.A. (Palestina, Caldas, Colombia). El ADN fue obtenido de cultivos monospóricos y de fructificaciones del hongo. Cada muestra se evaluó a partir de cultivos microbiológicos utilizando marcadores moleculares tipo ITS (genes ITS1, ITS2 y 5.8S). La caracterización molecular del ADN obtenido de la extracción directa de fructificaciones del hongo, evita la selección artificial en los medios de cultivo y garantiza un diagnóstico más real de la diversidad del patógeno en las plantaciones. La caracterización molecular permitió diferenciar entre M. perniciosa y M. roreri.
Palabras clave: ITS, ADN ribosomal, escoba de bruja, moniliasis del cacao.
AbstractMoniliophthora perniciosa presents discrepancies in its taxonomy and is the causal agent of cocoa witches' broom, which in Colombia is the second disease after moniliasis (Moniliophthora roreri). In this study 12 samples of the pathogen obtained from different cocoa materials from the Casa Luker S.A. Farm (Palestina, Caldas, Colombia) were analyzed. The DNA was obtained from monosporic cultures and from the fungus fructification. Each sample was evaluated from microbiological cultures using molecular markers ITS (ITS1, ITS2 and 5.8S genes). The Molecular ADN characterization obtained from direct extraction from the fungus fructification avoids artificial selection in the culture means and ensures a more realistic diagnosis of the pathogen diversity in the plantations. The molecular characterization allowed differentiation between M. pernicious and M. roreri.
Key words: ITS, ribosomal DNA, cocoa witches' broom disease, moniliasis of the cocoa.
El cacao presenta dos patógenos de alta incidencia, los hongos M. perniciosa y M. roreri, agentes causales de la enfermedad de la escoba de bruja y la moniliasis del cacao, respectivamente. Estos dos patógenos son los causantes de las mayores pérdidas en producción a nivel mundial. El estado fitosanitario del cultivo asociado a problemas de tecnología, capacitación técnica del recurso humano y la pérdida de la variabilidad genética de las plantaciones, han producido un decreciente interés por parte de los agricultores en producir cacao en Colombia (MINISTERIO DE AGRICULTURA, 2004).
El agente causal de la escoba de bruja del cacao fue clasificado inicialmente por STAHEL en 1915 como Marasmius perniciosa, más tarde fue transferido al género Crinipellis por SINGER (1942), y luego fue denominado Moniliophthora perniciosa por AIME & PHILLIPS-MORA (2005). Este patógeno se ha visto infectando brotes, inflorescencias y frutos de Theobroma cacao y es endémico para muchas otras especies del género Theobroma, Herrania y de las familias Solanaceae, Bignoniaceae y Malpighiaceae (RESENDE et al., 2000). Entre sus características microbiológicas se destacan un micelio morfológicamente variable y generalmente dependiente del biotipo, que le brinda el hospedero o incluso el medio de cultivo. Sus esporas son blancas, laminillas no separables, micelio tabicado y poseen esporas externas de origen sexual (basidiosporas) que se forman sobre órganos especiales llamados basidios; las basidiosporas son liberadas mediante un mecanismo de explosión, estas tienen vida corta, son fotosensibles y si no llegan a caer en un lugar apropiado para su desarrollo mueren en menos de una hora (GARCÉS, 2001).
La caracterización molecular de M. perniciosa y M. roreri, tradicionalmente ha partido de la extracción de ADN obtenido de cultivos monospóricos, lo cual garantiza un ADN de muy buena calidad, pureza y exclusivo del organismo estudiado (VILLEGAS, 2005), sumado a que tal como lo afirman RODRÍGUEZ & SAAVEDRA (2005), un factor limitante para investigar este patógeno es la escasez en campo de inóculo infectivo, debido a la estacionalidad y la fuerte influencia de la condiciones climáticas en la producción de los basidiocarpos. Pero la obtención de un ADN puro y de buena calidad proveniente de cultivos, también puede presentar una limitante en los estudios de patogenicidad y variabilidad genética, conferida por la selección artificial de esporas en los medios de cultivo, así se pueden caracterizar molecularmente esporas o biotipos que se presentan minoritariamente en el cultivo y que de ninguna manera presentan impacto en la producción.
Los estudios del ADN ribosomal en numerosos basidiomicetos, han encontrado que los espaciadores transcritos internos (ITS), muestran una variación que se convierte en un marcador taxonómico muy útil para distinguir entre especies (VILGALYS & GONZÁLEZ, 1990; WHITE et al., 1990; MILLER et al., 1999; CHEN et al., 2000). En este estudio se utilizaron herramientas microbiológicas y moleculares, encaminadas a fortalecer el conocimiento taxonómico de este patógeno, y además se propone una metodología rápida y confiable para su diagnóstico.
MATERIALES Y MÉTODOS Recolección, aislamiento y determinación de aislados
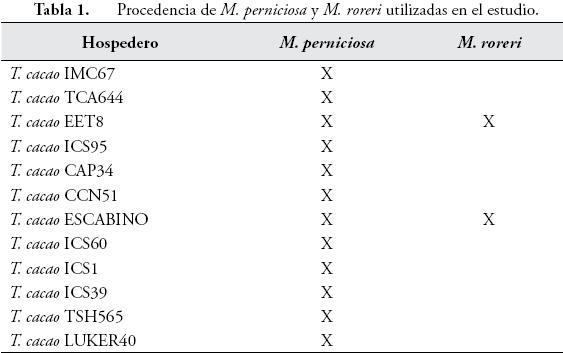
La recolección del material se realizó en la Granja Casa Luker S.A. (Palestina, Caldas, Colombia), ubicada a 1010 msnm, temperatura promedio de 22,5ºC y una precipitación promedio anual de 2128 mm. Se colectaron doce muestras de basidiocarpos de tallos para M. perniciosa y dos muestras de fruto para M. roreri, provenientes de diferentes materiales de cacao (Tabla 1). Estas muestras se rotularon y almacenaron en papel aluminio y luego en bolsas de papel, para después llevarlas a neveras de icopor con pilas refrigerantes. Las muestras permanecieron un máximo de 7 horas antes de su posterior tratamiento en el laboratorio.
Las escobas secas con presencia de basidiocarpos en diferentes estados de desarrollo, se llevaron al laboratorio. Las escobas que presentaron basidios en estadios tempranos de crecimiento, se almacenaron en cámara húmeda, y las que presentaron basidiocarpos en estadio óptimo de desarrollo, se almacenaron en bolsas y se guardaron a 4ºC.
El basidiocarpo seleccionado se adhirió del píleo a la tapa de una caja de petri, con ayuda de vaselina y posteriormente se descargaron las esporas en la base de la caja, la cual contenía Agar Agua al 3%. Con un bisturí se cortó la parte del medio de cultivo donde las esporas se localizaban y se montó sobre un portaobjetos, ubicando a 4X las esporas y traspasando a 10X una sola espora a cada uno de los medios de cultivo (PDA, Agar Malta y Agar Sabouraud). Se evaluó el crecimiento miceliar en los medios de cultivo, las cajas de petri se incubaron a 26ºC, por un periodo de ocho días. Para M. roreri se realizó la descarga de las esporas directamente desde los frutos infectados y se efectuó el mismo procedimiento hasta la obtención de cultivo monospórico (VILLEGAS, 2005).
Extracción del ADN
Para la extracción del ADN de cultivos monospóricos, se tomó una porción de micelio del hongo y se maceró con 200 micro-l de buffer de lisis (EDTA 0,5 M, NaCl 5 M, SDS 10 mM), se llevó al baño maría por 5 minutos a 96ºC. La extracción se realizó por el método fenol cloroformo alcohol isoamílico y precipitación con etanol y acetato de sodio, el precipitado se resuspendió en 30 micro-l de Tris-EDTA pH: 8,0 (Tris HCl 10 mM, EDTA 0,1 M) y se almacenó a 4ºC (modificado de GOODWIN & LEE, 1993.) La extracción directa del ADN de fructificaciones, correspondió al corte de basidiocarpos y lavado con hipoclorito al 3% durante 1 minuto y agua destilada por 5 minutos, se maceraron los basidiocarpos correspondientes a cada material de cacao con 200 micro-l de buffer de lisis (EDTA 0,5 M, NaCl 5 M, SDS 10 mM) e incubación a 56ºC por una hora, extracción por el método fenol-cloroformoalcohol isoamílico (adicionando 0,3 g de glass beads al inicio de la extracción), precipitación con etanol y acetato de sodio. El precipitado se resuspendió en 30 micro-l de Tris-EDTA pH: 8,0 y se conservó a 4ºC (modificado de NIH, 2005). La calidad y cantidad se determinó por comparaciones con concentraciones conocidas en geles de agarosa al 1%.
Amplificación por PCR
La amplificación de secuencias repetidas del rRNA que contienen las regiones espaciadoras ITS1, ITS2 y el gen 5.8S, que se han convertido en marcadores moleculares para numerosas especies de basidiomicetos, se llevó a cabo con los iniciadores ITS-1 (5'TCC GTA GGT GAA CCT GCG G3'), ITS-4 (5'TCC TCC GCT TAT TGA TAT GC3') propuestos por (WHITE et al., 1990; SARTORATO, et al., 2006). La mezcla de reacción se llevó a cabo en 20 micro-l que contenían de 100 a 200 ng de ADN, 800 microM de la mezcla dNTP's, 1,5 unidades de Taq polimerasa (invitrogen), 1,5 mM de MgCl2, 0,4 microM de cada iniciador y 2 micro-l del buffer 10X. La amplificación se realizó en un termociclador (Bio-Rad PTC-200) con el siguiente perfil térmico: desnaturalización inicial de 94ºC x 2,5 min, seguido de 35 ciclos de: Anillamiento: 60ºC x 40 s, Extensión: 72ºC x 30 s, Denaturación: 94ºC x 30 s, y una elongación final de 72ºC por 5 minutos. Los productos de amplificación se separaron en geles denaturantes de acrilamida al 4%, en cámara de secuenciamiento C.B.S. Scientific corp., revelados con nitrato de plata (SANGUINETTI et al., 1994) y analizados fotográficamente por medio del programa Corel Photo-Paint versión 11.633.
Dos muestras de M. perniciosa y dos de M. roreri (hospederos EET8 y Escabino), se evaluaron microbiológicamente en los diferentes medios de cultivo. Los medios que permitieron el mayor crecimiento del hongo fueron PDA (Papa - Dextrosa Agar) y Malta, por lo tanto se decidió producir el inóculo de los patógenos en estos dos medios de cultivo y realizar la extracción de ADN.
La amplificación de secuencias repetidas del rRNA con los iniciadores ITS1 - ITS4, presentaron un producto de 750 pb de M. perniciosa y de 710 pb para M. roreri, para todos los aislados monospóricos y basidiocarpos de los dos patógenos (Figura 1), confirmando el diagnóstico rápido y preciso de los dos patógenos. Además, tenemos que los resultados de nuestro estudio, confirman que el ADN obtenido del método de extracción directa es de buena calidad, lo cual se evidenció por su fácil amplificación a través del PCR, utilizando diferentes concentraciones de ADN y por la realización de amplificaciones comparativas con el ADN del cultivo monospórico. Ambos métodos de obtención de ADN presentaron buena sensibilidad, reproducibilidad y estabilidad en cuanto a su capacidad de amplificación por PCR con los marcadores ITS seleccionados.

DISCUSIÓN
La amplificación de secuencias repetidas del rRNA, específicamente de la región ITS, permitió corroborar rápida y eficientemente a M. perniciosa y diferenciarlo de M. roreri. En este trabajo no se encontró polimorfismo intraespecífico y, por lo tanto, no se pueden discriminar biotipos usando este marcador molecular, lo cual concuerda a lo reportado por VILGALYS & GONZÁLEZ (1990), WHITE et al. (1990), MILLER et al. (1999), CHEN et al. (2000) y ARRUDA et al. (2003), donde encontraron que los ITS en numerosos basidiomicetos es considerado un marcador taxonómico para diferenciar especies.
Los productos del PCR obtenidos de la amplificación con los iniciadores ITS1 - ITS4, para M. perniciosa presentaron un producto único de peso molecular aproximado correspondiente a 750 pb, lo que concuerda con lo encontrado por ARRUDA et al., (2003), en 120 aislados monospóricos de M. perniciosa provenientes de diferentes localidades y hospederos de cacao, y los trabajos de LANA (2004) realizados en tres regiones geográficas de Brasilia, que muestran que la variabilidad genética en esta región parece ser reducida para esta especie. Así tenemos que los iniciadores ITS1 e ITS4, para M. roreri, presentaron productos únicos de 710 pb, lo cual concuerda con lo reportado por PHILLIPS-MORA et al. (2007), quienes encontraron una banda diagnóstica aproximada de 741 pb para 94 aislados de M. roreri provenientes de Brasilia, utilizando los iniciadores ITS4 e ITS5.
Los estudios de M. perniciosa y M. roreri, presentan resultados interesantes y algunas veces contradictorios, así PHILLIPS-MORA et al. (2007) afirman que M. perniciosa y M. roreri, están íntimamente relacionados y probablemente comparten un ancestro común, evidenciando su relación, incluso proponen la hipótesis que M. roreri sea la fase asexual de M. perniciosa. Esta hipótesis es discordante con los resultados encontrados en este estudio, ya que si estos dos hongos fueran una misma especie, el diagnóstico molecular debería arrojar un perfil molecular similar para ambos aislamientos patogénicos.
La caracterización molecular de hongos fitopatógenos por lo general utiliza cultivos monospóricos realizados a partir de aislamientos en campo, disminuyendo las dificultades que representa la obtención de material de partida en cantidad suficiente bajo condiciones climáticas o estacionales desfavorables a la producción de basidiocarpos. Sin embargo, la desventaja evidente de la utilización de cultivos de hongos en el laboratorio es la selección artificial de esporas que ocurre en los medios artificiales de cultivo, privilegiando la proliferación de ciertos genotipos que no necesariamente representan adecuadamente la diversidad genética en campo.
En el presente estudio, los medios de cultivo PDA y Malta presentaron los mejores resultados para el crecimiento del patógeno, lo que concuerda con RODRÍGUEZ & SAAVEDRA (2005), quienes encontraron un buen crecimiento en PDA, así como un crecimiento del micelio más denso que los otros medios de cultivo. La extracción de ADN directamente de basidiocarpos del hongo puede ser un limitante para los análisis moleculares, dependiendo de la herramienta usada, así tenemos que marcadores moleculares aleatorios como polimorfismos de fragmentos amplificados al azar (RAPD), pueden presentar problemas a la hora de utilizar ADN obtenido bajo estas condiciones. Sin embargo, en este trabajo se demuestra que los marcadores de tipo específico ITS (especie específico) presentan ventajas en cuanto a la reducción de tiempo y costos de análisis, evitando además el riesgo de contaminación que representan los medios de cultivo.
Otro aspecto importante de la extracción directa, es que cuando se utilizan marcadores moleculares que determinan biotipos y diversidad genética (IGS y SSR), el uso de medios de cultivo favorece la amplificación del biotipo mayoritario o seleccionado en el cultivo, que posiblemente no sobrevivirían en el entorno natural, y no del basidiocarpo que se encuentra infectando la planta. La importancia de un método diagnóstico certero, radica en el nivel de confianza que ofrece a quien esté interesado en conocer el estado real de la situación fitosanitaria de un cultivo. Una vez que se tiene certeza del diagnóstico del patógeno, se pueden tomar medidas eficientes para el control que ahorren tiempo y dinero.
AGRADECIMIENTOSA la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas, por la financiación de este proyecto. Al Grupo de Investigación GEBIOME (Genética, Biodiversidad y Fitomejoramiento), por su soporte logístico, académico, técnico y humano. A Casa Luker y Granja Luker, por permitirnos colectar el material vegetal. Al laboratorio de Marcadores Moleculares de Cenicafé, dirigido por la Dra. Pilar Moncada, por su entrenamiento técnico y apoyo. A Luis Eduardo Zuluaga, por su asesoría en la etapa de campo.
AIME, M. & PHILLIPS-MORA, W. 2005. The causal agents of witches' broom and pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia 97(5):1012-1022. [ Links ]
ARANZAZU, F., 2000.- Escoba de bruja en Colombia su impacto económico y manejo, Tecnología para el mejoramiento del sistema de producción del cacao. Revta. Corpoica, 85-90. [ Links ]
ARRUDA, C.; FERREIRA, M. & FELIPE, M., 2003.- Nuclear and Mitochondrial rDNA variability in Crinipellis perniciosa from different geographic places. Origins and hosts, 25-37. [ Links ]
CHEN, W.; GRAU, R.; ADEE, E. & MENG, X., 2000.- A molecular marker identifying subspecific populations of the soybean brown stem rot pathogen, Phialophora gregata. Ecology and Population Biology, 90: 875-883. [ Links ]
CORREA, A., 1983.- Prueba de medios de cultivos artificiales para la fructificación de Crinipellis perniciosa: Tesis, Universidad de Caldas, Facultad de Ciencias Agropecuarias, Manizales. [ Links ]
GARCÉS, D., 2001.- Escoba de bruja del cacao. Revta. Agronomía, 24: 329-369. [ Links ]
GOODWIN, D.C. & LEE, S.B., 1993.- Microwave Miniprep of Total Genomic DNA from Fungi, Plants, Protists, and Animals for PCR. Biotechniques, 15: 438-444. [ Links ]
LANA, T., 2004. Caracterização genética e fisiológica de Crinipellis perniciosa. 91f. Tese (Doutorado em Fitopatologia) - Escola Superior de Agricultura 'Luiz de Queiroz', Universidade de São Paulo, Piracicaba, [ Links ]
MILLER, R.; SOARES, A. & LOPES, C.A., 1999.- Molecular comparison of Fusarium populations causing eumartii wilt and dry rot of potato in Brazil. Fitopatologia Brasileira, 24: 149-155. [ Links ]
MINISTERIO DE AGRICULTURA., 2004.- Manual del cultivo del Cacao: 132-139. Programa para el Desarrollo de la Amazonia, Proamazonia. [ Links ]
NIH (NATIONAL INSTITUTE OF HEALTH, USA), 2005.- DNA preparation from blood. Maryland, US. Disponible en: http://www.riedlab.nci.nih.gov/index.php/selected-publications (última consulta en línea 12 de enero de 2009). [ Links ]
PHILLIPS-MORA, W. & WILKINSON, M.J., 2000.- Biodiversity and Biogeography of the cacao (Theobroma cacao L.) pathogen Moniliophthora roreri (Cif.) Evans. Plant Pathology, 56: 911-922. [ Links ]
PHILLIPS-MORA, W; AIME, M. & WILKINSON, M., 2007.- Biodiversity and biogeography of the cacao (Theobroma cacao) pathogen Moniliophthora roreri in tropical America. Plant Pathology 56: 911-922. [ Links ]
RESENDE, M.; GUTEMBERG, B.; SILVA, L.; NIELLA, G.; CARVALHO, G.; SANTIAGO, D. & BEZERRA, J., 2000.- Crinipellis perniciosa proveniente de um novo hospedeiro, Heteropteys acutifolia, e'patogenicoao T. cacao. Fitopatologia Brasileira, 25: 88-91. [ Links ]
RODRÍGUEZ, E. & SAAVEDRA, J., 2005.- Ajuste de metodologías para la producción de basidiocarpos de escoba de bruja Crinipellis perniciosa (Stahel) Singer en medios artificiales. Ascolfi Informa, 31: 1-3. [ Links ]
SANGUINETTI, C.J; DIAS, N. & SIMPSON, A.J., 1994.- Rapid silver staning and recovery of PCR products separate on polycrylamide gels. Biotechniques, 17: 914-921. [ Links ]
SARTORATO, A.; NECHET, K.L. & HALFELD-VIEIRA, B.A., 2006.- Diversidad e genética de isolados de Rhizoctonia solani coletados em feijão-caupi no Estado de Roraima. Fitopatología Brasileira, 31: 24-33. [ Links ]
SINGER, R., 1942.- Monographic study of the genera Crinipellis and Chaetocalathus. Lilloa Tucuman, 8: 441-514. [ Links ]
STAHEL, G., 1915.- Bull. Dep. Landb. Suriname 33, 26pp. (description sp. nov) [ Links ]
VILGALYS, R. & GONZÁLEZ, D., 1990.- Organization of ribosomal DNA in the basidiomycete Thanatephorus praticola. Current Genetics, 18: 277-280. [ Links ]
VILLEGAS, R.L., 2005.- Aislamiento monospórico de Moniliophthora perniciosa. Instituto Colombiano Agropecuario. [ Links ]
WHITE, T.; BRUNS, T.; LEE, S. & TAYLOR, J., 1990.- Amplification and direct sequencing of fungal RNA genes for phylogenetics: 315-322 (in) M.A. INNIS, D.H. GELGARD, J.J. SNINSKY & T.J. WHITE (eds.) PCR Protocols: a Guide to Methods and Applications. San Diego, CA: Academic Press. [ Links ]

