










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.15 no.1 Manizales Jan./June 2011
PARTE II: MUSCULATURA , SANGRE Y SISTEMA NERVIOSO*
APIS MELLIFERA LINAEUS, 1758 (HYMENOPTERA) HISTOLOGY AS AN ENTOMOLOGIC CONTRIBUTION
PART II: MUSCULATURE , BLOOD AND NERVOUS SYSTEM
* FR: 26-II-2011. FA: 24-V-2011.
1 Director Grupo de Investigaciones Entomológicas y Ambientales – GENA. Profesor Asociado Universidad Industrial de Santander. Calle 9 con carrera 27. Coordinador Grupo Biodiversidad Corporación CTAS. Consultor FUSDECOL. E-mail: alfvillalmo@gmail.com
2 Biólogo UIS. Estudiante de Maestría en Entomología, Universidad Nacional de Colombia. E-mail: jcagudelo@gmail.com
3 Profesor Asociado Universidad Industrial de Santander. Director Laboratorio de Histotecnia, UIS. Calle 9 con carrera 27. E-mail: darrietadap@gmail.com
Se presenta la segunda parte de los resultados de los cortes de cabeza y protórax de Apis mellifera. Se exponen las observaciones realizadas al sistema muscular, células sanguíneas y sistema nervioso.
Palabras clave: docencia, cerebro, hemocitos, insectos, ojos.
AbstractThe second part of the results of the head and prothorax cuts of Apis mellifera are presented. Observations about the muscular system, blood cells and nervous system are exposed.
Palabras clave: teaching, brain, hemocytes, insects, eyes.
En este documento se amplían los resultados de los cortes de A. mellifera, cuyo objetivo fue establecer una metodología para realizar montajes permanentes de insectos bajo las condiciones del Laboratorio de Histotecnia-UIS, generando material didáctico para facilitar el estudio de anatomía e histología de insectos en asignaturas relacionadas con este grupo.
MATERIALES Y MÉTODOSSe realizaron cortes histológicos seriados de cabeza y protórax de A. mellifera, considerando los planos vertical, horizontal y sagital. A los mejores cortes se les hicieron fotos digitales con una cámara SONY CyberShot adaptada a un microscopio Olympus, las cuales fueron analizadas y procesadas con los programas Microsoft Office Picture Manager, CorelDRAW Graphics Suite X3 y Adobe Photoshop CS2.
RESULTADOSSe realizaron más de mil cortes, que fueron organizados en 310 montajes permanentes, de los cuales se seleccionaron 152 por la calidad del montaje, y se hicieron un total de 191 fotos digitales. Con el propósito de comprender e interpretar mejor los resultados, la información está organizada en siete secciones, en este documento se presentan las secciones IV a VII; las secciones anteriores se encuentran en VILLALOBOS-M. et al. (2011). Se muestran algunas de las imágenes obtenidas, se hacen descripciones de los montajes y se señalan las estructuras más evidentes.
IV. MUSCULATURASegún RESH & CARDÉ (2003), el sistema muscular de los insectos presenta tres tipos de fibras: esqueléticas, responsables de postura y locomoción; viscerales, que son las que causan el movimiento de tubo digestivo, túbulos de Malpighi y parte del sistema reproductivo; y cardiacas, que provocan contracción en masa del tejido que forma la aorta dorsal para impulsar su contenido asociado, la sangre. Las células musculares de las fibras viscerales y cardiacas son pequeñas, filiformes y mononucleadas. Según BLUM (1995), si bien la nomenclatura es similar a la utilizada en vertebrados, se usa sólo por la comodidad de su estudio, puesto que la estructura, fisiología y control son diferentes en ambos grupos.
En insectos, las fibras musculares esqueléticas se insertan en el exoesqueleto y son estriadas, polinucleadas y con núcleos en posición central (Fig. 13A); estos núcleos se encuentran parcial o totalmente fusionados, evidenciando el posible origen sincitial del aparato nuclear (PESSACQ, 1998). Con base en el grosor, el tamaño y la forma de las fibras y los paquetes de fibras, los músculos esqueléticos de los insectos se clasifican en fibrilares, microfibrilares y tubulares (BLUM, 1995). En la Figura 13B se observan las dos primeras: las fibrilares que corresponden a los músculos longitudinales dorsales observados en el protórax, y la microfibrilar que corresponde al músculo tergocoxal posterior o remotor tergal.

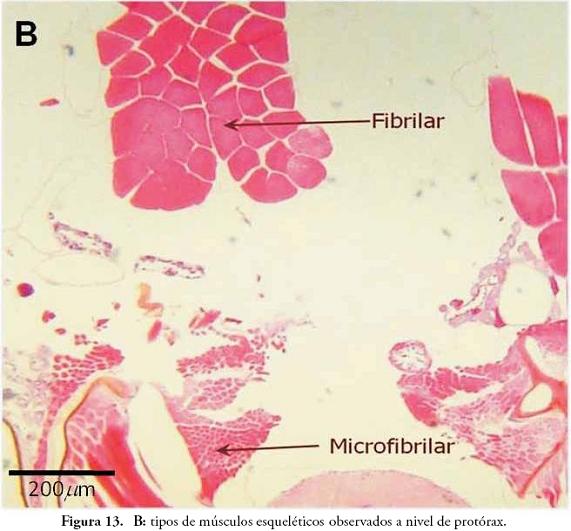
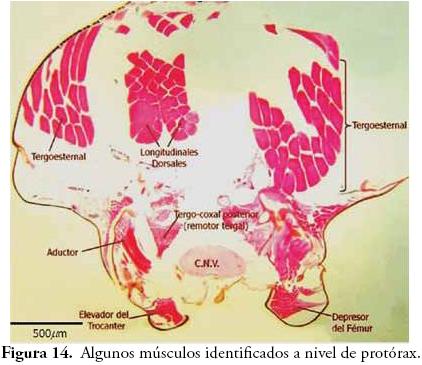
En la Figura 14 se identifican algunos músculos a nivel de protórax. En posición dorsal, se observan cuatro paquetes musculares muy evidentes y de gran volumen: los dos paquetes ubicados hacia el centro y prácticamente fusionados, corresponden a los músculos longitudinales dorsales, y los dos paquetes localizados hacia los lados y claramente insertados en el tergo, son los músculos tergoesternales. En posición ventral se observan varios músculos asociados a las patas, entre los cuales se pudieron identificar: aductores, depresores del fémur, elevadores del trocanter y tergocoxal posterior.
V. CÉLULAS SANGUÍNEAS
Las células sanguíneas de los insectos reciben el nombre de hemocitos, y según BLUM (1995) se reconocen unos cinco tipos básicos llamados: Prohemocitos, Plasmocitos, Granulocitos, Esferulocitos y Coagulocitos. Sin embargo, CHAPMAN (1998) describe 8 clases: Plasmatocitos, Esferulocitos (células de Spherule), Oenocitoides, Granulocitos, Cistocitos, Prohemocitos, Coagulocitos y Adipohemocitos.
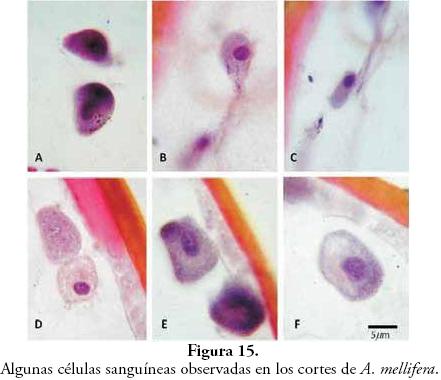
Si bien el conocimiento sobre las células sanguíneas de los insectos aún no está totalmente dilucidado, es posible encontrar algunos esquemas que sirven de herramienta para identificarlas y que permiten observar su gran diversidad. En el presente trabajo se lograron varias imágenes (Fig. 15) de hemocitos similares a los descritos por BLUM (1995), CASELÍN-C. et al. (2008) y CHAPMAN (1998).
Las células sanguíneas de los insectos y de los artrópodos en general, no son responsables del transporte de gases, pues existe toda una compleja red de tubos (troncos traqueales, tráqueas, sacos aéreos, etc.) que cumplen con dicha función. De esta forma, las funciones de los hemocitos incluyen procesos de almacenamiento de varios tipos de sustancias, así como defensa del organismo basada en mecanismos de coagulación, fagocitosis, encapsulamiento, melanización, respuesta inmune, etc. (CHAPMAN, 1998).
Basados en las descripciones de CHAPMAN (1998), se realizaron las identificaciones de las células sanguíneas encontradas. En la Figura 15-A se observan dos Plasmatocitos, con proyecciones citoplasmáticas bastante notorias. Desde B hasta F se observan Granulocitos, aunque D posiblemente es un Adipohemocito.
VI. SISTEMA NERVIOSOEn la mayoría de los montajes permanentes obtenidos en los diferentes planos de corte en protórax y cabeza de A. mellifera, se observan numerosos elementos del sistema nervioso, como cordón nervioso ventral, ganglios, nervios óptico y ocelar, componentes del cerebro y cuerpos pedunculados. Para su fácil comprensión, la información se organizó en subtemas de la siguiente manera:
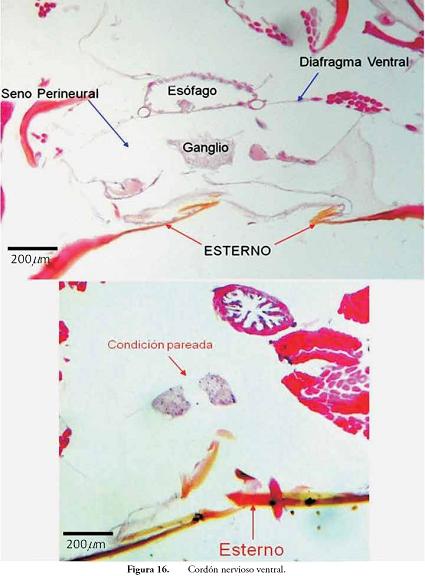
Cordón nervioso ventral: es la estructura de los insectos para "conectar" el ganglio supraesofágico o cerebro con el resto de los ganglios y nervios de los segmentos corporales. La naturaleza, posición y componentes se comparten con otros grupos de artrópodos y anélidos, los cuales también tienen el cordón en posición ventral. La naturaleza pareada es muy evidente en algunos grupos, como en los anélidos; sin embargo, en insectos estas estructuras se acercan y se fusionan hasta hacerse bastante difícil observar dicha característica. En la Figura 16, se aprecia el Cordón Nervioso en posición ventral y su condición ganglionar, además se aprecia su ubicación dentro del espacio llamado Seno Perineural, el cual se encuentra limitado por un delicado tejido conectivo que corresponde al Diafragma Ventral. En la foto inferior de la Figura 16, se aprecia la condición pareada del Cordón Nervioso.
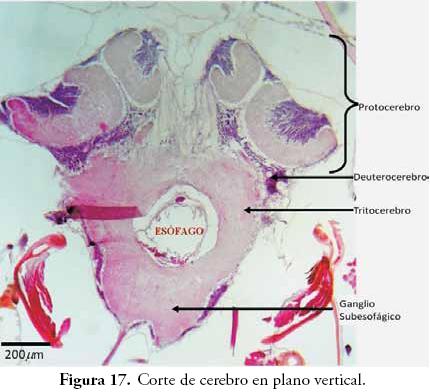
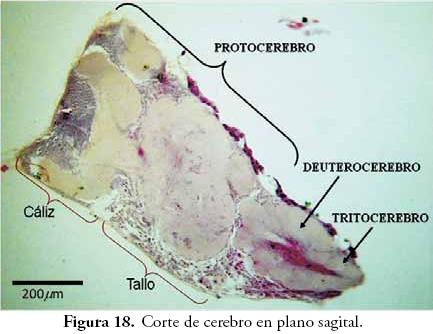
Cerebro: los cortes de cerebro de A. mellifera (Figs. 17 y 18), dieron los resultados más complejos pero también las imágenes fotográficas más espectaculares tomadas durante el presente trabajo. El cerebro de los insectos puede llegar a estar conformado por más de un millón de neuronas (RESH & CARDÉ, 2003); corresponde al centro del control motor, de la recepción de estímulos y de las numerosas respuestas fisiológicas. La mayoría de los entomólogos acepta que esta compleja estructura está formada por la fusión de tres pares de ganglios preorales que se encuentran en posición supraesofágica, sin embargo, existen otras interpretaciones con leves modificaciones como las encontradas en GOODRICH (1918), DUPORTE (1946), MANTON (1960) y MATSUDA (1965).
En la Figura 17 se observa cómo el cerebro está conformado por tres pares de ganglios, que forman tres zonas bien definidas; en esta misma figura, se aprecia su posición supraesofágica. En el cerebro de los insectos, cada par de ganglios forma las tres regiones del cerebro llamadas: Protocerebro, Deuterocerebro y Tritocerebro. El Protocerebro está encargado de inervar ojos, ocelos, vertex y frente, el Deuterocerebro inerva las antenas, y el Tritocerebro inerva componentes del aparato bucal, especialmente musculatura labral.
En algunos grupos, como se observa en los cortes de A. mellifera, el Protocerebro presenta estructuras complejas llamadas cuerpos pedunculados (mushroom body). Según RESH & CARDÉ (2003), el cerebro de la mayoría de los Hymenoptera puede estar formado por más de un millón de neuronas, de las cuales unas 300 mil estarían conformando los cuerpos pedunculados, lo cual se puede comprender mejor al observar el espacio que ocupan estas estructuras en el cerebro (Figs. 17 y 18).
Cuerpos pedunculados: en los cortes de cerebro de A. mellifera (Figs. 17 y 18) se aprecian un par de estructuras complejas dentro del protocerebro. Según RESH & CARDÉ (2003), estas estructuras fueron descubiertas por Félix Dujardin en 1850, quien las asoció con memoria e inteligencia. Varios estudios señalan que los cuerpos pedunculados pueden estar relacionados con el aprendizaje y la memoria, sobre todo de los olores. Además, estarían vinculados con la memoria asociativa, con el registro de los estímulos sensoriales, con el control motor y con la memoria espacial, todos ellos, importantes factores en grupos con ciertos niveles de sociabilidad (STRAUSFELD & LI, 1999; STRAUSFELD, 2002).
En la Figura 19 se observa un corte vertical de los cuerpos pedunculados señalándose sus componentes, donde se ve que su estructura corresponde a las descripciones de MOBBS (1982) e ITO et al. (1997), quienes afirman que se observa un tallo y un cáliz, y que este último se divide en dos mitades. ITO et al. (1997) demostraron que cada mitad del cáliz proviene de una cuarta parte del neuroblasto embriológico correspondiente.
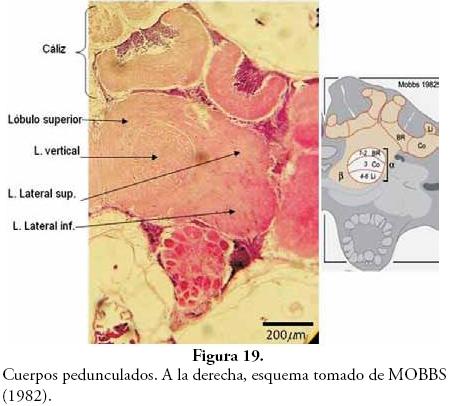
Trabajos realizados por FARRIS & STRAUSFELD (2001), han permitido comparar los cuerpos pedunculados de Periplaneta americana y Apis mellifera con las descripciones de LEE et al. (1999) hechas para Drosophila. Además, según RESH & CARDÉ (2003) un trabajo similar fue realizado por STRAUSFELD, y corresponde a un estudio comparativo de cerebros de varios grupos de insectos.
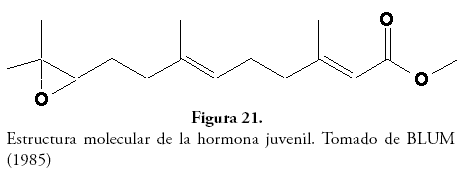
Glándulas neurosecretoras: en algunos cortes se encontraron pequeñas masas basófilas (coloración que sugiere funciones de secreción), las cuales fueron comparadas con los esquemas y descripciones propuestos por HARTFELDER (2000), AYALI et al. (2002), RESH & CARDÉ (2003) y NUSS et al. (2008), identificándose como corpora allata y corpora cardiaca (Fig. 20). Las corpora allata están involucradas con la producción de un sesquiterpenoide, terpeno de 15 carbonos y derivado de tres isoprenos, llamado hormona juvenil (Fig. 21), que se encuentra presente en todas las ninfas y larvas de los insectos, y cuya acción primaria es prevenir en avance del proceso de metamorfosis, el cual responde a niveles de ecdisona producida en glándula protorácica.
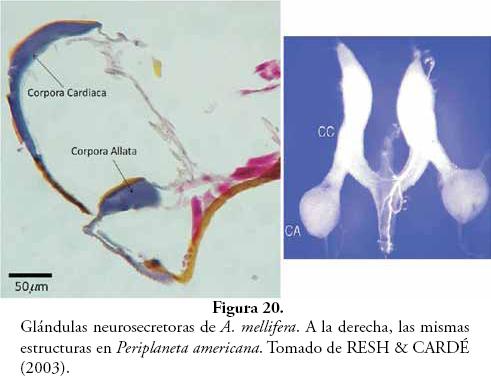
Las corpora cardiaca, junto con las corpora allata, el ganglio subesofágico y el cerebro, forman un complejo neurosecretor que se encarga de producir varios grupos de neuropéptidos (Nps) que actúan como mensajeros químicos extracelulares. Los Nps son pequeñas moléculas de menos de 50 aminoácidos, que se comportan como importantes reguladores de procesos fisiológicos. Se han identificado más de 150 Nps, los cuales pueden actuar como neurotransmisores, neurohormonas o neuromoduladores de amplio rango (RESH & CARDÉ, 2003).
VII. ÓRGANOS FOTORRE CEPTORESLos insectos presentan varios tipos de receptores de estímulos, todos ellos corresponden a modificaciones y especializaciones de componentes tegumentarios, que son inervados por terminaciones nerviosas que llevan el estímulo a centros nerviosos que analizan la señal y regulan la producción de algún tipo de respuesta. Algunos de estos receptores están relacionados con la captación de estímulos químicos, como los palpos, el ovipositor y las antenas, son los llamados quimiorreceptores. Otros se especializan en recibir estímulos mecánicos (incluyendo los sonoros), como palpos, tarsómeros y tímpanos, y son llamados mecanorreceptores. También existen órganos especializados en captar estímulos térmicos, estímulos magnéticos y niveles de humedad: termorreceptores, magnetorreceptores e higrorreceptores, respectivamente. Finalmente, se encuentran los propiorreceptores que establecen la posición y ubicación de su propio cuerpo, y los fotorreceptores que se especializan en captar estímulos lumínicos, en este último grupo se incluyen los ojos y ocelos. En las siguientes dos secciones, se exponen los resultados de los cortes de ojos y ocelos de A. mellifera, señalando sus componentes básicos.
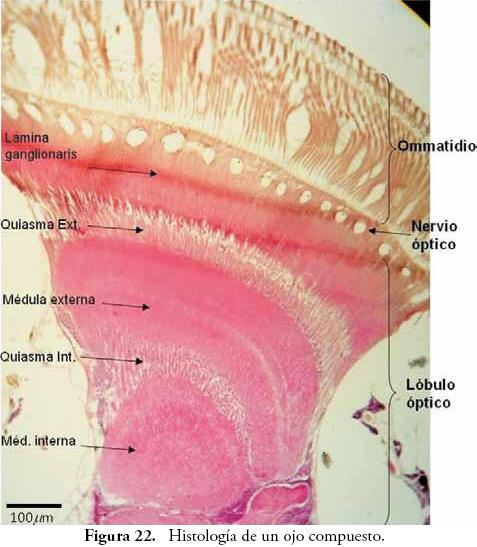
Ojos: en los insectos en general, este órgano es llamado ojo compuesto, pues está conformado por numerosas unidades independientes: los omatidios. Estas unidades captan el estímulo lumínico que se encuentra delante de ellos, y al final conforman una imagen en mosaico que el cerebro interpreta como un todo. Algunos grupos, como Collembola, Thysanura, Thysanoptera, áfidos, chiches aleyródidos y hormigas presentan unas pocas unidades que hacen del ojo compuesto una estructura relativamente simple. Sin embargo, la mayor parte de los insectos presentan cientos o miles de omatidios que ocupan gran parte de la superficie de la cabeza. Por ejemplo, en libélulas se han calculado cerca de 25.000 unidades. En la Figura 22, se observa cómo el ojo de A. mellifera cuenta con numerosas unidades, tan solo en ese fragmento de corte es posible contar varias docenas de omatidios.
En la Figura 22, se observa un segmento de un corte transversal de ojo, allí se observan los componentes principales de su estructura. Inicialmente se aprecian los omatidios, los cuales se encuentran conectados al lóbulo óptico por medio de una región estrecha que corresponde al nervio óptico o retina. El lóbulo óptico se aprecia como una estructura grande y compleja con varias zonas muy bien definidas, y cuya función general es llevar los mensajes lumínicos desde los omatidios y la retina, hasta las zonas del Protocerebro que están especializadas en la interpretación de formas y colores.
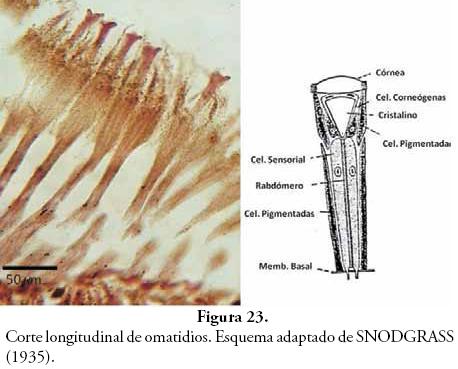
En la Figura 23, se observa un detalle del corte longitudinal de un grupo de omatidios, en donde se aprecia la ausencia de la córnea (seguramente fragmentada al momento del corte), pero apicalmente se observan estructuras eosinófilas que corresponden a los cristalinos, a lado y lado de estas estructuras se advierten zonas con abundantes gránulos, que corresponden a los pigmentos de las células corneógenas y células pigmentadas, cuya función es crear el efecto de cámara oscura para direccionar los haces de luz hacia la zona sensorial debajo de ellas, y que se observa como una zona larga y estrecha donde se ubican las células sensoriales que también se encuentran acompañadas por más células pigmentadas, que se observan como pequeñas manchas oscuras.
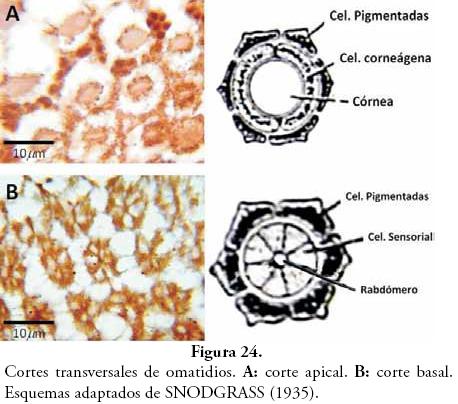
En la Figura 24, se observan cortes transversales de los omatidios, los cuales fueron realizados a diferentes niveles. La foto superior es a un corte bastante apical, donde se aprecia una región central eosinófila que corresponde al cristalino, y rodeándolos se identifican las células pigmentadas. En la foto inferior es un corte basal, donde se identifican entre cinco y seis células sensoriales (claras) rodeadas por un número similar de células pigmentadas (oscuras). En el centro de las células sensoriales se encuentra un espacio bien definido que corresponde al rabdómero.
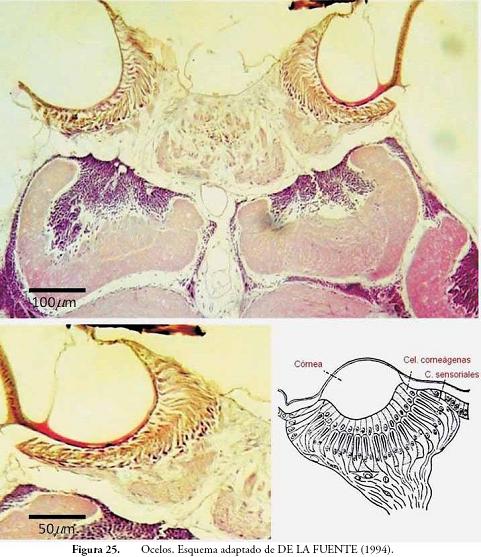
Ocelos: estas estructuras están conformadas por un único lente o córnea, asociado a numerosas células sensoriales (Fig. 25). Los ocelos son órganos fotorreceptores con limitaciones en su capacidad para definir colores y formas. En A. mellifera se observan tres ocelos, dos laterales y uno medio. En la Figura 25, se observa el corte transversal de los ocelos laterales, el central no se observa por no estar en el mismo plano. En esta figura, se aprecian los componentes de los ocelos, así como el nervio ocelar que es el que lleva los estímulos lumínicos a zonas especializadas del Protocerebro.
AGRADECIMIENTOS
Los autores desean agradecer a todas y cada una de las personas que hicieron posible este artículo, especialmente a nuestras familias y amigos por su paciencia en los momentos en que no estuvimos presentes por estar realizando esta investigación. A nuestros amigos Wilson Medina (q.e.p.d.), Jorge Villamizar Cobos, Arnaldo Báez, Jorge Niño Rondón, Adolfo Hernández García e Inés Johanna Gómez por su colaboración y acompañamiento permanente. A los Doctores Jaime Alberto Camacho, Sergio Isnardo Muñoz y Óscar Gualdrón por su apoyo para asistir al XXXV Congreso de la Sociedad Colombiana de Entomología en la ciudad de Cali, donde se presentaron avances de esta investigación.
AYALI, A.; ZILBERSTEIN, Y. & COHEN, N., 2002.- The locust frontal ganglion: a central pattern generator network controlling foregut rhythmic motor patterns. Journal of Experimental Biology, 205: 2825-2832. [ Links ]
BLUM, M.-S., 1985.- Fundamentals of insect physiology. Athens, Georgia, USA: John Wiley Publications. 598p. [ Links ]
CASELÍN-C., S.; LLANDERAL-C., C.; RAMÍREZ-C., A.; SOTO-H., M. & MÉNDEZ-M., J.T., 2008.- Caracterización morfológica de hemocitos de la hembra de Dactylopius coccus (Hemiptera: Coccoidea). Agrociencia, 42: 349-355. [ Links ]
CHAPMAN, R.F., 1998.- The insects: structure and function. 4th ed. USA: Cambridge University Press. 788p. [ Links ]
DE LA FUENTE, J.A., 1994.- Zoología de Artrópodos. Madrid, España: Interamericana-McGraw Hill. 805p. [ Links ]
DUPORTE, E.M., 1946.- Observations on the morphology of the face in insects. Journal of Morphology, 79: 371-417. [ Links ]
FARRIS, S.M. & STRAUSFELD, N.J., 2001.- Development of laminar organization in the mushroom bodies of the cockroach: Kenyon cell proliferation, outgrowth, and maturation. J. Comp. Neurol., 439: 331-351. [ Links ]
GOODRICH, E.S., 1918.- On the development of the segments of the head in Scyllium. Q. J. Microsc. Sci, 63: 1-30. [ Links ]
HARTFELDER, K., 2000.- Insect juvenile hormone: from "status quo" to high society. Braz. J. Med. Biol. Res., 33 (2): 157-177. [ Links ]
ITO, K.; AWANO, W.; SUZUKI, K.; HIROMI, Y. & YAMAMOTO, D., 1997.- The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development, 124: 761-771. [ Links ]
LEE, T.; LEE, A. & LUO, L., 1999.- Development of the Drosophila mushroom bodies: Sequential generation of three distinct types of neurons from a neuroblast. Development, 126: 4065-4076. [ Links ]
MANTON, S.M., 1960.- Concerning head development in the arthropods. Biol. Rev., 35: 265-282. [ Links ]
MATSUDA, R., 1965.- Morfology and evolution of the insect head. Memoirs of the American Entomological Institute, 4: 1-334. [ Links ]
MOBBS, P.G., 1982.- The brain of the honeybee Apis mellifera. I. The connections and spatial organization of the mushroom bodies. Philos. Trans. R. Soc. Lond. Biol. Sci., 298: 309-354. [ Links ]
NUSS, A.B.; FORSCHLER, B.T.; CRIM, J.W. & BROWN, M.R., 2008.- Distribution of neuropeptide F-like immunoreactivity in the eastern subterranean termite, Reticulitermes flavipes. Journal of Insect Science, 8: 1-18. [ Links ]
PESSACQ, T.P., 1998.- Algunas notas sobre aspectos histológicos del tejido muscular de Himenóptero. I. El aparato nuclear. Rev. Colombiana Entomología, 24 (1-2): 71-74. [ Links ]
RESH, V. & CARDÉ, R., 2003.- Enclyclopedia of Insects. USA: Elsevier Science, 1295p. [ Links ]
SNODGRASS, R.E., 1935.- Principles of Insect Morphology. London, England: McGraw-Hill, 667p. [ Links ]
STRAUSFELD, N.J., 2002.- Organization of the honey bee mushroom body: Representation of the calyx within vertical and gamma lobes. J. Comp. Neuro., 450: 4-33. [ Links ]
STRAUSFELD, N.J. & LI, Y., 1999.- Representation of the calyces in the medial and vertical lobes of cockroach mushroom bodies. J. Comp. Neuro., 409: 626-646. [ Links ]
VILLALOBOS-M., A.; AGUDELO, J.C. & ARRIETA-P., D., 2011.- Histología de Apis mellifera Linnaeus, 1758 (Hymenoptera) como aporte entomológico, Parte I: Regiones corporales, organización anatómica e integumento. Bol.cient.mus.hist.nat., 14 (2): 201-214. [ Links ]

