










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.15 no.2 Manizales July/Dec. 2011
EFFECTS OF ADITION OF SE MINAL PLASMA ON THE ELIMINATION OF CYTOPLASMIC DROPLETS IN OF SUS SCROFA LINAEUS, 1758 BOAR SEMEN
* FR: 10-VIII-2011. FA: 22-XI-2011.
1 Ph.D. Instituto de Biotecnología Agropecuaria, Universidad de Caldas. Calle 65 No. 26-10, Manizales, Colombia. E-mail: fhenao@ucaldas.edu.co
2 MVZ de la Universidad de Caldas.
3 MVZ de la Universidad de Caldas.
4 MVZ de la Universidad Cooperativa de Colombia seccional Bucaramanga.
Las gotas citoplásmicas (GCs) son remanentes del citoplasma que quedan adheridos al espermatozoide después de la espermatogénesis, constituyen la anormalidad espermática más frecuente en porcinos, y se relacionan claramente con baja fertilidad. Hay serios indicios de que la fructosa y el AMPc del plasma seminal intervienen en la maduración espermática, en el desprendimiento de las GCs, y en la reacción acrosómica. El objetivo del presente estudio fue evaluar el efecto de la adición de plasma seminal, en el desprendimiento de las GCs en machos con diagnóstico de persistencia de las mismas. En el estudio se emplearon tres verracos (dos con persistencia de GCs y uno normal) de tres a cinco años de edad, alojados en la granja Montelindo de la Universidad de Caldas; a los cuales se les realizaron análisis seminales semanales, completos, durante cuatro meses. Se llevó a cabo un arreglo factorial 3x3x2 (adición a la FR de los machos con persistencia de GCs de 0%, 20% de PSMS y 20% de PSMGCs; 0, 60 y 120 minutos de incubación, y 16 y 37°C de temperaturas de incubación) en un diseño de bloques completos al azar, analizado mediante análisis de varianza y prueba de Tukey. La incubación del semen de machos con persistencia de GCs con PSMGCs redujo más del 4% las GCs respecto a la incubación sin PS y con PSMS; igualmente se registró reducción de aproximadamente el 5% en las GCs, al aumentar el tiempo de incubación de 0 a 120 minutos, y alrededor de 2% al llevar la temperatura de incubación de 16 a 37°C.
Palabras clave: semen porcino, gotas citoplasmáticas, plasma seminal, evaluación seminal, calidad seminal.
AbstractThe cytoplasmic droplets (CDs) are remnants of the cytoplasm that are attached to the sperm after spermatogenesis. CDs constitute the most frequent sperm abnormality in pigs and are clearly related to low fertility. There are serious indications that fructose and the seminal plasma AMPc are involved in sperm maturation in the CDs detachment and in the acrosome reaction. The objective of this study was to evaluate the effect of the addition of seminal plasma in the CDs detachment in males with diagnosis of persistence of such detachment. Three boars (two with persistence of CDs and one normal) from three to five years of age, housed in the Montelindo farm at Universidad de Caldas were used in the study. These boars were performed complete seminal analysis weekly during four months. A factorial arrangement 3x3x2 (addition to the males FR with CDs persistence of 0%, 20% of SPHM and 20% of SPMCDs; 0, 60 and 120 minutes incubation, and 16, and 37°C incubation temperature) was carried out in a randomized complete blocks design, analyzed through variance analysis and Tukey's test. Incubation of males semen with persistence of CDs with SPMCDs decreased more than 4% the CDS with regards to incubation without SP and SPHM; similarly, there was reduction of approximately 5% in CDs when increasing incubation time from 0 to 120 minutes, and about 2% when increasing incubation temperature from 16 to 37°C.
Key words: boar semen, cytoplasmic droplets, seminal plasma, semen quality.
Las gotas citoplásmicas (GCs) constituyen la anormalidad espermática más frecuente en porcinos (FISCHER et al., 2005; GÓMEZ, 2010), y se relacionan claramente con baja fertilidad (LOBERCAMP et al., 2007). Además, se asocian con descarte de dosis seminales, de machos, y con baja rentabilidad de los centros de producción de material seminal. Las GCs son pequeños remanentes del citoplasma original del espermatozoide, que quedan adheridos al espermatozoide después de la espermatogénesis (KAPLAN et al., 1984); estas estructuras migran desde la porción proximal del tracto intermedio del espermatozoide a la porción distal, durante el paso por el epidídimo, en el proceso de maduración (FISCHER et al., 2005), para luego eliminarse (KAPLAN et al., 1984). HARAYAMA et al. (1996) y KATO et al. (1996), encontraron que su eliminación ocurre uno a dos minutos después de la eyaculación, cuando los espermatozoides se mezclan con componentes del plasma seminal (PS) como la fructosa y el adenosín monofosfato cíclico (AMPc). Se considera que un macho tiene persistencia de GCs, cuando permanentemente presenta un porcentaje ≥ a 20% de estas, después del proceso natural de eliminación. Es común encontrar defectos estructurales de la cola asociados a GCs, denominados reflejo distal de la pieza media y ovillo-gota; el primero consiste en el plegamiento de la cola entorno a la gota, en un ángulo de 180° (FISCHER et al., 2005); y el segundo se da por el enrollamiento total o parcial de la cola sobre la gota citoplásmica (BONET et al., 1993).
El PS es una mezcla de secreciones provenientes de la red testicular, el epidídimo y las glándulas sexuales accesorias (CENTURIÓN et al., 2003; CABALLERO et al., 2004); funciona como vehículo y medio de inmersión de los espermatozoides después de la eyaculación (GARCÍA, 2007), participa en la regulación de procesos relacionados con nutrición, protección, maduración, movilidad (WABERSKI et al., 1995), y capacitación del espermatozoide (CALVETE et al., 1997). El PS tiene, además, factores que influyen en la respuesta inmune, las contracciones uterinas, la relajación del istmo oviductal (WABERSKI et al., 1995), la ovulación de la cerda (SARAVIA et al., 2009), el transporte de los espermatozoides (ROZEBOOM, 2000) y el reconocimiento y la unión entre gametos (CALVETE et al., 1996). Los componentes del PS varían entre machos (CABALLERO, 2007), y entre las fracciones del eyaculado en razón de su origen. El PS de la fracción pre-espermática está compuesto principalmente por mucina proveniente de las glándulas bulbouretrales y uretrales; el PS de la fracción rica en espermatozoides (FR), particularmente de la primera porción (primeros 10 ml), está compuesto por bicarbonato de origen prostático, y por secreciones proteicas del epidídimo y las vesículas seminales; el PS de la fracción post-espermática o fracción pobre en espermatozoides (FP), está compuesto, también, por bicarbonato de la próstata, y por proteínas, fructosa y AMPc, secretadas en las vesículas seminales (HARAYAMA et al., 1996; RODRÍGUEZMARTÍNEZ et al., 2005; SARAVIA et al., 2009). La fructosa es el azúcar que utilizan normalmente los espermatozoides como fuente de energía, para llevar a cabo los procesos metabólicos (FOLEY et al., 1964), y el AMPc funciona como segundo mensajero, participa en la activación de la tirosina kinasa y la fosforilación proteica, en la capacitación espermática y la fecundación (ADACHI et al., 2008). Tanto la fructosa como el AMPc, intervienen en la maduración espermática, en el desprendimiento de las GCs (HARAYAMA et al., 1996), en la reacción de acrosoma, en la movilidad e hiperactivación espermática y en la fecundación (ADACHI et al., 2008). Se ha observado que a diluciones bajas, la adición de PS al semen refrigerado de porcino puede disminuir la capacitación prematura, mejorar la viabilidad y la capacidad fecundante de los espermatozoides (WABERSKI et al., 1995; GARNER et al., 2001). Las espermadhesinas son proteínas del plasma seminal porcino (PSPs, de su nombre en inglés porcine seminal plasma proteins) y se clasifican como las AQN (alanina-glutamina-asparagina) I-III y AWN (alanina-triptófano-asparagina), que tienen capacidad de unión a la heparina y los heterodímeros PSP I y PSP II, que no tienen esta propiedad (CALVETE et al., 1997; CABALLERO et al., 2004). Ambos grupos se encuentran en mayor proporción en la FP, debido a que aumentan a medida que decrece la concentración de espermatozoides (RODRÍGUEZ-MARTÍNEZ et al., 2005; GARCÍA, 2007). La importancia de las espermadhesinas radica en que se unen a la superficie del espermatozoide durante la eyaculación, y contribuyen a la unión con el ovocito (JONÁKOVÁ & TICHÁ, 2004). Además, se ha encontrado su expresión en la cola del epidídimo (EKHLASI-HUNDRIESER et al., 2002), sin embargo, no está clara aún su función en la maduración espermática. Las proteínas del epidídimo también hacen parte de las proteínas del PS, y aunque participan en la maduración espermática (OKAMURA et al., 1992), no se conoce su efecto en el desprendimiento de las GCs. El objetivo del presente estudio fue evaluar el efecto de la adición de plasma seminal, en el desprendimiento de las GCs en machos con diagnóstico de persistencia de las mismas.
MATERIALES Y MÉTODOSPara este estudio se emplearon tres verracos de tres a cinco años de edad, alojados en la granja Montelindo de la Universidad de Caldas, vereda Santagueda del municipio de Palestina (Caldas) (1024 msnm, 24°C de temperatura media, 2200 mm de precipitación anual y 76% de humedad relativa; corrales individuales de 7 x 3 m, 2 kg/animal/día de un balanceado comercial con 12% de proteína y agua limpia a voluntad). A los machos se les realizaron análisis seminales semanales, durante cuatro meses; los espermogramas de los tres primeros meses se emplearon para establecer la condición de cada animal con respecto a la presencia de GCs. Dos machos fueron diagnosticados como persistentes y uno como normal. Las eyaculaciones se realizaron mediante la técnica de la mano enguantada entre las 04:00 y 06:00 horas; se separó la FR y la FP, y se transportaron inmediatamente al Laboratorio de Control de Calidad Seminal del Instituto de Biotecnología Agropecuaria de la Universidad de Caldas. Se efectuó dilución en BTS (de su nombre en inglés Beltsville Thawing Solution), a una concentración de 30 x 106 espermatozoides/ml. El PS se obtuvo por centrifugación de cada FP a 800 rpm por 15 minutos, y se almacenó a temperatura ambiente hasta su uso; así se consiguió PS del macho sano (PSMS) y PS de los machos con persistencia de GCs (PSMGCs).
Cada espermograma consideró: temperatura, pH, concentración en fotómetro Spermacue-Minitüb y en cámara de Bürker en microscopio de campo claro con objetivo de 40X; morfología mediante fijación con solución salina formolada y evaluación de 300 espermatozoides en microscopio de campo claro con objetivo de 40X. La supervivencia espermática y la integridad acrosómica de la siguiente manera: se incubó 400 ul de una dilución seminal a 30 x 106 espermatozoides/ml de BTS con 100 ul de FITC-PNA y 40 ul de PI a 37°C por 15 min y esta mezcla se fijó en 200 ul de solución salina formolada (CABALLERO et al., 2004); se depositaron 10 ul de la combinación anterior en un portaobjetos, se colocó cubreobjeto e inmediatamente se observó en un microscopio Eclipse 80i Nikon con epifluorescencia, con filtros de 440 nm de longitud de onda. En esta prueba los espermatozoides se clasificaron según su respuesta a los colorantes en 4 categorías celulares: categoría 1 (C1), espermatozoides muertos con acrosoma intacto o PI+/FITC-PNA-; categoría 2 (C2), espermatozoides muertos con acrosoma reaccionado (reacción acrosómica falsa o por degeneración) o PI+/FITC-PNA+; categoría 3 (C3), espermatozoides vivos con acrosoma intacto o PI-/FITC-PNA- (espermatozoides no coloreados); categoría 4 (C4), espermatozoides vivos con acrosoma reaccionado (reacción acrosómica verdadera) o PI-/FITC-PNA+.
La supervivencia espermática se evaluó, también, mediante tinción con Eosina-Nigrosina (E/N), en microscopio de campo claro con objetivo de 40X. La integridad acrosómica, también fue evaluada mediante fijación con glutaraldehído al 2% en BTS en microscopio de contraste de fases (Alphaphot YS2, Nikon) con objetivo de 40X (PURSEL & JOHNSON, 1974). La Integridad bioquímica de la membrana plasmática (IBMP), se evaluó mediante el test corto de hinchazón hipoosmótica (sHOST, de su nombre en inglés short Hypoosmotic Swelling Test) (PÉREZ-LLANO et al., 2001).
El estudio se planteó como un arreglo factorial 3x3x2 usando: 3 niveles de adición de PS a la FR de los machos con persistencia de GCs (0%, 20% de PSMS y 20% de PSMGCs), 3 tiempos de incubación (0, 60 y 120 minutos), y 2 temperaturas de incubación (16 y 37°C), en un diseño de bloques completos al azar (bloqueo por fecha de muestreo). Se realizó un análisis de varianza mediante el programa GLM de SAS y las variables que arrojaron diferencias significativas se compararon por la prueba de Tukey.
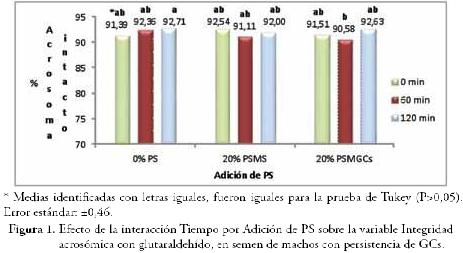
RESULTADOS Y DISCUSIÓNEl efecto de la interacción entre los tres factores (tiempo de incubación, adición de plasma y temperatura de incubación), sobre las variables seminales evaluadas (GCs totales, % de células normales, integridad bioquímica de la membrana plasmática, integridad acrosómica, supervivencia espermática por E/N e integridad acrosómica combinada con supervivencia espermática) no fue significativo (P>0,05). Se registró efecto significativo (P≤0,05) de la interacción Tiempo de incubación por Adición de PS, sobre la integridad acrosómica y sobre el porcentaje de espermatozoides muertos con acrosoma intacto. El porcentaje de espermatozoides con acrosoma normal registrado con la combinación de 0% de PS a 120 minutos (92,71%), fue superior (P≤0,05) al obtenido con 20% de PSMGCs a 60 minutos (90,58%) (Fig. 1), los promedios restantes fueron iguales. Los porcentajes de espermatozoides muertos con acrosoma intacto no registraron diferencias significativas (P>0,05) en la prueba de Tukey. Los valores promedio de integridad acrosómica fluctuaron entre 90,58 y 92,71%, muy por encima de 70%, considerado valor mínimo aceptable.
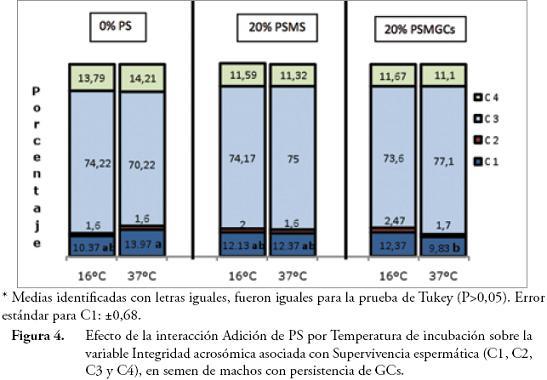
La interacción Adición de PS por Temperatura de incubación tuvo efecto altamente significativo (P≤0,01), sobre la Integridad bioquímica de la membrana plasmática y sobre el porcentaje de espermatozoides muertos con acrosoma intacto. Los porcentajes de espermatozoides positivos a HOST tendieron a bajar con la incubación a 37°C, sin embargo, esta reducción solo fue significativa (P≤0,05) en espermatozoides incubados sin plasma (bajó de 63,76% a 49,74%). En general los valores de HOST+ estuvieron por encima del 50% considerado como valor mínimo normal (50%) (Fig. 3). Estos resultados concuerdan con el trabajo de SALA-ECHAVE et al. (2008), donde se encontró que la incubación de semen a temperaturas mayores de 20°C redujo la funcionalidad bioquímica de la membrana, la movilidad y el porcentaje de acrosomas normales. El porcentaje de células muertas con acrosoma intacto obtenido con 0% de PS a 37°C (13,97%), fue superior (P≤0,05) al obtenido con 20% de PSMGCs (9,83%) a la misma temperatura; los valores restantes para esta variable fueron iguales (P>0,05) (Fig. 4).
La evaluación de la Integridad acrosómica en asociación con la supervivencia celular mediante el empleo de fluorocromos (PI/FITC-PNA), registrada en las Figuras 2 y 4 permite apreciar que los valores de porcentaje de células vivas con acrosoma intacto estuvieron siempre por encima del 70%, valor considerado como el límite inferior aceptable para supervivencia espermática. El porcentaje de espermatozoides vivos con reacción acrosómica verdadera fluctuó entre 9,06% y 14,12%, y el porcentaje de espermatozoides muertos con reacción acrosómica falsa entre 1,15% y 2,47%.
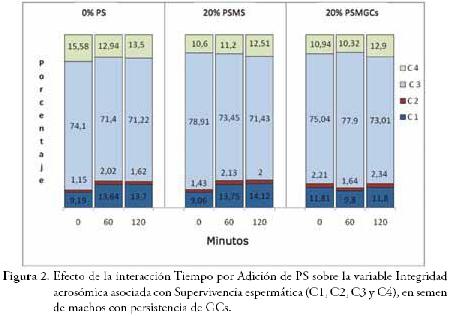

La interacción Tiempo de incubación por Temperatura de incubación, no registró efecto significativo (P>0,05) sobre las variables seminales evaluadas.
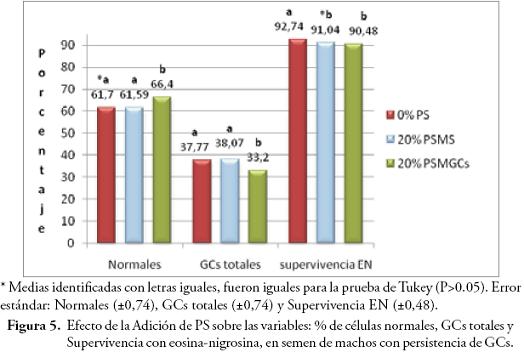
La adición de PS afectó de manera altamente significativa (P≤0,01) las variables: GCs totales, % de células normales, y supervivencia espermática por E/N. El porcentaje de gotas totales obtenido con la adición de 20% de PSMGCs (33,2%), fue inferior (P≤0,05) al obtenido con 20% de PSMS (38,07%) y 0% PS (37,77%), consecuentemente el porcentaje de células normales fue superior al incubar con 20% de PSMGCs (66,4%) (Fig. 5). Este resultado concuerda con el trabajo de HARAYAMA et al. (1996), donde la adición de fructosa y AMPc del PS propicia la liberación de las GCs; y difiere con el trabajo de BERGER & CLEGG (1985), que hallaron mayor porcentaje de GCs retenidas al adicionar PS. La adición de 20% de PSMS no propició el desprendimiento de GCs, debido posiblemente a diferencias en los componentes del PS entre machos (CABALLERO, 2007). La supervivencia espermática presentó porcentaje inferior con la adición de 20% de PSMGCs (90,48%) y 20% PSMS (91,04%), comparado al porcentaje obtenido con 0% PS (92,74%) (Fig. 5). Los valores promedio de supervivencia espermática fluctuaron entre 90,48 y 92,74%, por encima de 70% considerado valor mínimo aceptable.
El factor Tiempo de incubación registró efecto altamente significativo (P≤0,01) sobre GCs totales, % de normales, integridad bioquímica de la membrana y significativo (P≤0,05) sobre la supervivencia espermática por E/N. Los porcentajes de GCs fueron diferentes (P≤0,05) en todos los tiempos de incubación (40,63% a los 0 min; 35,51% a los 60 min y 32,9% a los 120 min), con obtención del porcentaje de GCs más bajo a los 120 minutos, a consecuencia de lo anterior el porcentaje de células normales fue superior al aumentar el tiempo de incubación (66,46%) (Fig. 6).
ZOU & YANG (2000), describieron que la incubación del semen, sin la adición de ningún componente, propicia la liberación de las GCs, que ocurrió en mayor proporción cuando se incubó por un tiempo mayor a 60 min.El porcentaje de espermatozoides positivos a HOST a los 0 min (58,51%) y 60 min (54,48%), fue superior que el tiempo de incubación a 120 min (52,76%), lo cual coincide con los reportes de SALA-ECHAVE et al. (2008); y la supervivencia espermática a los 0 min (92,49%), fue superior que a los 60 min (90,85%) de incubación (Fig. 6), concordante con los resultados de KAWANO et al. (2004) y GARCÍA et al. (2010), con semen congelado-descongelado.
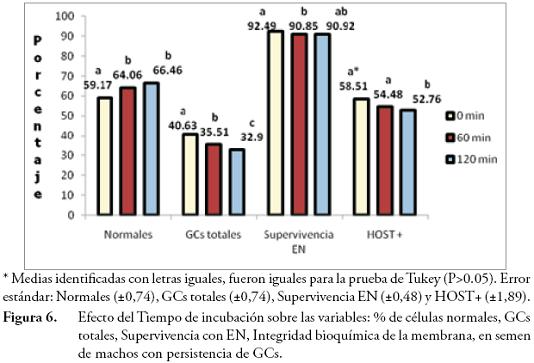
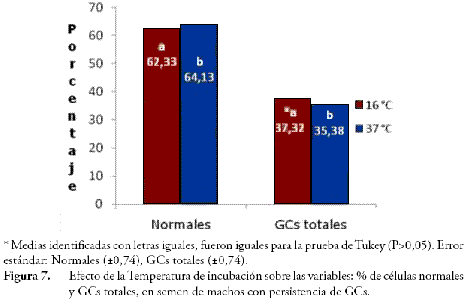
La Temperatura de incubación tuvo efecto significativo (P≤0,05) sobre las GCs totales y sobre el % de células normales, y efecto no significativo (P>0,05) sobre la integridad acrosómica y la supervivencia por E/N. Los porcentajes de GCs fueron inferiores al incubar a 37°C (35,38%), que la incubación a 16°C (37,32%), consecuentemente el % de células normales aumentó a 64,13% en la incubación a 37°C (Fig. 7), esto coincide con lo reportado por ZOU & YANG (2000), donde la liberación de GCs era mayor a temperaturas 4 y 15°C que las temperaturas de incubación a 39 y 20°C.
CONCLUSIONES
• La interacción entre los tres factores (tiempo de incubación, adición de plasma y temperatura de incubación) y la interacción de Tiempo de incubación por Temperatura de incubación no afectaron de manera significativa las variables dependientes medidas (P>0,05).
• A pesar de que se registró efecto por lo menos significativo (P≤0,05) de la interacción Tiempo de incubación por Adición de PS sobre la integridad acrosómica y sobre el % de espermatozoides muertos con acrosoma intacto, y de la interacción Adición de plasma seminal por Temperatura de incubación sobre el % de espermatozoides HOST+ y sobre el % de espermatozoides muertos con acrosoma intacto, estos resultados no trascienden debido a que las variables en mención registraron valores siempre por encima de los mínimos normales.
• La incubación del semen de machos con persistencia de GCs con PSMGCs redujo el % de GCs y por ende aumento el % de células normales.
• Los valores registrados para % de GCs y para recuento de espermatozoides HOST+ se redujeron de manera por menos significativa (P≤0,05), al aumentar el tiempo de incubación a 120 minutos, recíprocamente aumentó el % de células normales. A pesar de que el tiempo de incubación afectó el % de vivos con E/N, estos valores fueron muy superiores al límite normal inferior.
• El porcentaje de GCs en el semen se redujo de manera significativa (P≤0,05) al aumentar la temperatura de incubación de 16 a 37°C, de manera concomitante aumentó el % de células normales.
El PS es un componente importante del eyaculado de los porcinos que en condiciones naturales cumple una función específica que no es muy clara y requiere de más estudios, al igual que la incubación espermática a diferentes temperaturas. Adicional a esto, la variabilidad de criterios de recolección de semen porcino en Colombia hace imprescindible encaminar acciones investigativas hacia este punto, donde se determine con precisión el efecto que tienen las diferentes fracciones del eyaculado sobre la calidad seminal.
ADACHI, J., TATE, S. & MIYAKE, M., 2008.- Effects of protein phosphatase inhibitor calyculin a on the postacrosomal protein serine/threonine phosphorylation state and acrosome reaction in boar spermatozoa incubated with a cAMP analog. J Reprod Dev., 54 (3): 171-6. [ Links ]
BERGER, T. & CLEGG, E.D., 1985.- Effect of male accessory gland secretions on sensitivity of porcine sperm acrosomes to cold shock, initiation of motility and loss of cytoplasmic droplets. J Anim Sci., 60: 1295-1302. [ Links ]
BONET, S., BRIZ, M. & FRADERA, A., 1993.- Ultrastructural abnormalities of boar spermatozoa. Theriogenology, 40: 383-396. [ Links ]
CABALLERO, I., 2007.- Estudio del plasma seminal y la espermadhesina PSP-I/PSP-II sobre la funcionalidad de los espermatozoides de verraco: Tesis, Universidad de Murcia, Departamento de Medicina y Cirugía Animal, Facultad de Veterinaria, España. [ Links ]
CABALLERO, I., VÁZQUEZ, J.M., GIL. M.A., CALVETE, J.J., ROCA, J., SANZ, L., PARRILLA, I., GARCÍA, E.M., RODRÍGUEZ-MARTÍNEZ, H. & MARTÍNEZ, E.A., 2004.- Does seminal plasma psp-i/psp-ii spermadhes in modulate the ability of boar spermatozoa to penetrate homologous oocytes in vitro? J Androl., 25: 1004-1012. [ Links ]
CALVETE, J.J., DOSTÁLOVÁ, Z., SANZ, L., ADERMANN, K., THOLE, H.H., TÖPFER-PETERSEN, E., 1996.- Mapping the heparin-binding domain of boar spermadhesins. FEBS Letters, 379: 207-211. [ Links ]
CALVETE, J.J., ENSSLIN, M., MBURU, J., IBORRA, A., MARTÍNEZ, A., ADERMANN, K., WABERSKI, D., SANZ, L., TÖPFER-PETERSEN, E., WEITZE, K.F., EINARSSON, S. & RODRÍGUEZ-MARTÍNEZ, E., 1997.- Monoclonal antibodies against boar sperm zonapellucida-binding protein awn-1. Characterization of a continuous antigenic determinant and immunolocalization of awn epitopes in inseminated sows. Biol Reprod., 57: 735-742. [ Links ]
CENTURIÓN, F., VÁZQUEZ, J.M., CALVETE, J.J., ROCA, J., SANZ, L., PARRILLA, I., GARCÍA, E.M. & MARTÍNEZ, E.A., 2003.- Influence of porcine spermadhesins on the susceptibility of boar spermatozoa to high dilution. Biol Reprod., 69: 640-646. [ Links ]
EKHLASI-HUNDRIESER, M., SINOWATZ, F., GREISER DE WILKE, I., WABERSKI D. & TÖPFER-PETERSEN, E., 2002.- Expression of spermadhesin genes in porcine male and female reproductive tracts. Mol Reprod Dev., 61 (1): 32-41. [ Links ]
FISCHER, K.A., LEYEN, K.V., LOVERCAMP, K.W., MANANDHAR, G., SUTOVSKY, M., FENG, D., SAFRANSKI, T. & SUTOVSKY, P., 2005.- 15-lipoxygenase is a component of the mammalian sperm cytoplasmic droplet. Reproduction, 130: 213-222. [ Links ]
FOLEY, C.W., HEIDENREICH, C.J., HARRINGTON, R.B., JONES, H.W. & ERB, R.E., 1964.- Changes in fructose, lacticacid, ph, and motility of boar semen during incubation at 37°C. J Anim Sci., 23: 558-561. [ Links ]
GARCÍA, E.M., 2007.- Análisis, función y aplicaciones biotecnológicas de las proteínas del plasma seminal de porcino PSP I y PSP II: Tesis, Universidad de Murcia, Facultad de Veterinaria, Departamento de Medicina y Cirugía Animal, España. [ Links ]
GARCÍA, J.C., DOMÍNGUEZ, J.C., PEÑA, F.J., ALEGRE, B., GONZÁLEZ, R., CASTRO, M.J., HABING, G.G. & KIRKWOOD, R.N., 2010.- Thawing boar semen in the presence of seminal plasma: Effects on sperm quality and fertility. Anim Reprod Sci., 119: 160-165. [ Links ]
GARNER, D.L., THOMAS, C.A., GRAVANCE, C.G., MARSHALL C.E., DEJARNETTE, J.M. & ALLEN, C.H., 2001.- Seminal plasma addition attenuates the diltion effect in bovine sperm. Theriogenology, 56: 31-40. [ Links ]
GÓMEZ, G., 2010.- Dinámica de la calidad seminal de verracos del centro-occidente colombiano: Tesis, Universidad de Caldas, Maestría en Sistemas de Producción Agropecuaria, Manizales, Colombia. [ Links ]
HARAYAMA, H., SHIBUKAWA, T., MIYAKE, M., KANNAN, Y. & KATO, S., 1996.- Fructose stimulates shedding of cytoplasmic droplets from epididymal boar spermatozoa. Reprod Fertil Dev., 8: 1039-1043. [ Links ]
JONÁKOVÁ, V. & TICHÁ, M., 2004.- Boar seminal plasma proteins and their binding properties. A review Collection of Czechoslovak Chemical Communications, 69: 461-475. [ Links ]
KAPLAN, M., RUSSELL, L.D., PETERSON, R.N. & MARTAN, J., 1984.- Boar sperm cytoplasmic droplets: Their ultrastructure, their numbers in the epididymis and at ejaculation and their removal during isolation of sperm plasma membranes. Tissue and Cell, 16: 455-468. [ Links ]
KATO, S., SHIBUKA, W.A.T., HARAYAMA, H., et al., 1996.- Timing of shedding and dissentegration of cytoplasmic droplet from boar and goat spermatozoa. J. Reprod. Dev., 42: 237-241. [ Links ]
KAWANO, N., SHIMADA, M. & TERADA, T., 2004.- Motility and penetration competence of frozen-thawed miniature pig spermatozoa are substantially altered by exposure to seminal plasma before freezing. Theriogenology, 61: 351-364. [ Links ]
LOVERCAMP, K.W., SAFRANSKY, T.J. & FISCHER, K.A., 2007.- High resolution light microscopic evaluation of boar semen quality sperm cytoplasmic droplet retention in relationship with boar fertility parameters. Arch Androl, 53: 219-228. [ Links ]
OKAMURA, N., DACHEUX, F., VENIEN, A., ONOE, S., HUET, J.C. & DACHEUX, J.L., 1992.- Localization of a Maturation-Dependent Epididymal Sperm Surface Antigen Recognized by a Monoclonal Antibody Raised against a 135-Kilodalton Protein in Porcine Epididymal Fluid. Biol Reprod., 47: 1040-1052. [ Links ]
PÉREZ-LLANO, B., LORENZO, J.L., YENES, P., TREJO, A. & GARCÍA-CASADO, P. 2001.- Short hypoosmotic swelling test for the prediction of boar sperm fertility. Theriogenology, 56: 387-398. [ Links ]
PURSEL, V.G. & JOHNSON, L.A., 1974.- Glutaraldehyde fixation of boar spermatozoa for acrosome evaluation. Theriogenology, 1: 63-68. [ Links ]
RODRÍGUEZ-MARTÍNEZ, H., SARAVIA, F., WALLGREN, M., TIENTHAI, P., JOHANNISSON, A., VÁZQUEZ, J.M., MARTÍNEZ, E., ROCA, J., SANZ, L. & CALVETE, J.J., 2005.- Boar spermatozoa in the oviduct. Theriogenology, 63:514-535. [ Links ]
ROZEBOOM, K.J., 2000.- Evaluating of boar semen quality (en) N. C. S. UNIVERSITY (ed.) Animal Science Facts. Extension Swine Husbandry, North Carolina State, 1-8. [ Links ]
SALA-ECHAVE, R., REGUERA, G., GARCÍA-CASADO, P. & PÉREZ-LLANO, B., 2008.- Influencia del calentamiento de las dosis seminales porcinas sobre la calidad espermática durante la I.A. Av. Tecnol. porc., 5 (3): 34-41. [ Links ]
SARAVIA, F., WALLGREN, M., JOHANNISSON, A., CALVETE, J.J., SANZ, L., PEÑA, F.J., ROCA, J. & RODRÍGUEZ-MARTÍNEZ, H., 2009.- Exposure to the seminal plasma of different portions of the boar ejaculate modulates the survival of spermatozoa cryopreserved in MiniFlatPacks. Theriogenology, 71: 662-675. [ Links ]
WABERSKI, D., SÜDHOFF, H., HAHN, T., JUNGBLUT, P.W., KALLWEIT, E., CALVETE, J.J., ENSSLIN, M., HOPPEN, H.O., WINTERGALEN, N. & WEITZE, K.F., 1995.- Advanced ovulation in gilts by the intrauterine application of a low molecular mass pronase-sensitive fraction of boar seminal plasma. J Reprod Fertil., 105: 247-252. [ Links ]
ZOU, C.X. & YANG, Z.M., 2000.- Evaluation on sperm quality of freshly ejaculated boar semen during in vitro storage under different temperatures. Theriogenology, 53: 1477-1488. [ Links ]

