Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas v.15 n.2 Manizales jul./dic. 2011
BUTTERFLY DIVERSITY AND RELATIONSHIP WITH INGA ORNATA KUNTH (MIMOSOIDAE) REPRODUCTIVE PHENOLOGY IN A FOREST SHEPHERDING AGROECOSYSTEM OF QUINDÍO, COLOMBIA
* FR: 23-VII-2011. FA: 15-X-2011.
1 Autor para correspondencia. Licenciado en Biología y Educación Ambiental. Grupo de Investigación en Biodiversidad y Educación, Universidad del Quindío (BIOEDUQ), A.A. 460. E-mail: oschumar@gmail.com
2 Bióloga. MSc. Centro de Investigaciones en Biodiversidad (CIBUQ). Universidad del Quindío, A.A. 460. E-mail: rociogarcia06@yahoo.es
3 Licenciado en Biología y Educación Ambiental. Universidad del Quindío. E-mail: wgomezmarin@yahoo.es
4 Licenciado en Biología y Educación Ambiental. Universidad del Quindío. E-mail: larcencord@gmail.com
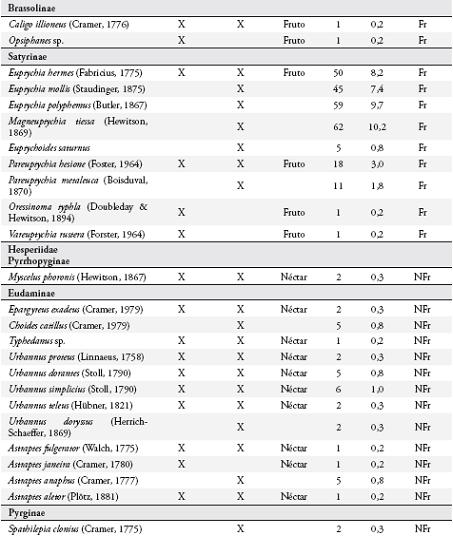
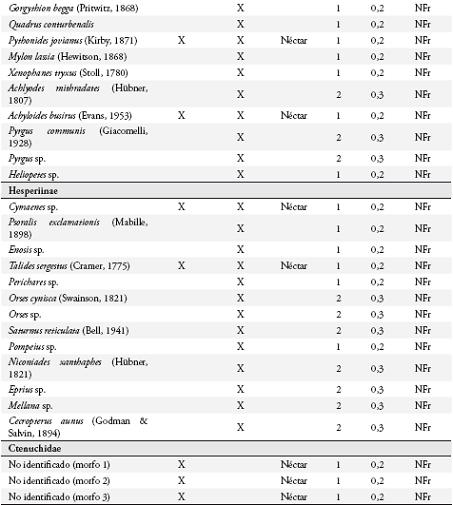
Se estudió la diversidad de mariposas diurnas y su relación con la fenología reproductiva del guamo, I. ornata, en un agroecosistema ganadero en Quindío, Colombia. Se hicieron muestreos mensuales en dos transectos de 50 m x 2 m utilizando redes entomológicas, trampas cebadas con fruta en descomposición y observaciones de las mariposas visitantes de I. ornata. En estos transectos se contabilizó mensualmente el número de flores y frutos disponibles de I. ornata. Se registró un total de 142 especies de mariposas, 116 en el potrero y 59 asociadas a los guamos. Se encontró una mayor representación del gremio de mariposas no frugívoras en el área de estudio. La riqueza, abundancia y diversidad de mariposas mostraron una tendencia a aumentar durante los meses con mayor disponibilidad de flores y frutos, aunque no se encontró una correlación significativa entre estas variables. Los resultados de este estudio destacan la importancia del uso de I. ornata como recurso alimentario por parte de las comunidades de mariposas adultas, principalmente por las no frugívoras.
Palabras clave: mariposas diurnas, gremios, uso recursos, Inga ornata, agroecosistemas.
AbstractThe diversity of diurnal butterflies and their relationship with the guamo, Inga ornata, reproductive phenology in a forest shepherding agroecosystem in Quindío, Colombia was studied. Monthly samples of two transects of 50 x 2 m were carried out using entomological nets, traps baited with decomposed fruit and direct observations of l. ornate visiting butterflies were performed. The number of l. ornate flowers and fruits available in these transects was also counted monthly. A total of 142 butterfly species was recorded, 116 in pastures and 59 associated with I. ornata trees. A higer representation of the nectar feeding guild was found in the area of study. The richness, abundance and diversity of butterflies showed a tendency to increase during the months with higher availability of flowers and fruits, but there was not a significant correlation between these variables. The results of this study highlight the importance of I. ornata as a food resource for adult butterflies, especially for nectar feeding guild.
Key words: diurnal butterflies, guilds, resource use, Inga ornata, agroecosystems.
Los bosques andinos premontanos de la Cordillera Central de Colombia han sido fuertemente fragmentados y deforestados durante los siglos XIX y XX (ETTER, 1998), esta situación ha sido particularmente acentuada en la región del valle geográfico del río Cauca, donde el incremento de la agricultura y la ganadería han generado diferentes mosaicos de vegetación dominados por sistemas productivos y bosques en diferentes estados de regeneración que se encuentran altamente fragmentados (ALZATE, 1994). A pesar de su transformación, estos sistemas presentan una alta diversidad de insectos debido a su heterogeneidad espacial (GARCÍA et al., 2001; HERNÁNDEZ et al., 2003; HORNER et al., 2003).
Los lepidópteros son componentes conspicuos de la mayoría de ecosistemas, donde presentan una alta especificidad hacia las plantas de las cuales se alimentan (FAGUA, 1996). En el trópico, las comunidades de mariposas adultas se agrupan en dos gremios de alimentación, aquellas que obtienen la mayoría de sus requerimientos nutricionales del néctar floral y las que se alimentan de jugos de frutos en descomposición o de savia fermentada de los árboles (DEVRIES et al., 1997). Sin embargo, estos organismos presentan una gran variedad de hábitos alimenticios entre los que se destacan el consumo de polen, sales minerales y excretas de aves o mamíferos (RAY & ANDREWS, 1980; ADLER & PEARSON, 1982; VALENCIA et al., 2005). Los patrones de diversidad y riqueza de las mariposas neotropicales han sido ampliamente estudiados (DEVRIES et al., 1997; DEVRIES & WALLA, 2001), sin embargo algunos aspectos de su ecología, como el forrajeo de los adultos, han sido pobremente documentados (HALL & WILLMOTT, 2000).
Inga ornata es una especie que se distribuye en la cordillera de los Andes de Colombia, Ecuador y Perú entre los 800 a 2200 m, y es utilizado como árbol de sombra en plantaciones de café y cacao (PENNINGTON, 1997), por su significativo aporte de nutrientes al suelo y prolongación de la vida productiva de los cafetales (FARFÁN & MESTRE, 2004). Esta especie presenta una floración masiva que atrae a una gran variedad de visitantes, principalmente insectos y aves que se alimentan de las flores, néctar, polen, nectarios extraflorales y de frutos (MARÍN-GÓMEZ, obs. pers.). El objetivo de este trabajo fue determinar la diversidad de mariposas que habitan en un agroecosistema ganadero y su relación con la fenología reproductiva de I. ornata.
MATERIAL Y MÉTODOSEste estudio se realizó en la Reserva Natural "La Montaña del Ocaso", ubicada en la Cordillera Central de los Andes, Colombia, departamento del Quindío, en la confluencia del río Roble con el río La Vieja (4°34'818'' N - 75°51'870'' W), a una elevación entre los 975 y los 1100 msnm. El paisaje se caracteriza por parches de Guadua angustifolia, un fragmento de bosque secundario de 106 ha y potreros con dominancia de I. ornata.
Se realizó un seguimiento de la fenología reproductiva (floración y fructificación) de I. ornata en un transecto de 100 m x 20 m ubicado en el potrero. Las observaciones se realizaron con binoculares 10x42 cada 15 días entre septiembre de 2005 a marzo de 2006. Durante cada observación se contó el número de flores adultas y de frutos maduros en los árboles.
Los muestreos de mariposas se realizaron durante los meses de septiembre de 2005 y marzo de 2006, con una intensidad de dos salidas de campo mensuales de cuatro días cada una. En el potrero se ubicaron dos transectos de 50 m. En uno de estos se ubicaron seis trampas Van Someren Rydon a una distancia de 10 m y a una altura de 4 m del suelo. Para cada trampa se usó un cebo elaborado con banano y cerveza, las cuales se revisaron cada 24 h. Adicionalmente, se hicieron colectas utilizando redes entomológicas con un esfuerzo de muestreo de 16 horas día/2 personas, desde las 09:00 h hasta las 17:30 h. Con binoculares 10x42 se realizaron observaciones de la actividad de forrajeo de las mariposas que visitaron las flores y los frutos de árboles de I. ornata ubicados en el transecto, entre las 06:00-18:00 h, durante los periodos de mayor oferta de flores y frutos.
Las mariposas colectadas en campo se sacrificaron por medio de presión digital. Los especímenes fueron determinados y depositados en el Museo de Artrópodos de la Universidad del Quindío (MAUQ) y Museo de Historia de Natural de la Universidad Caldas (MHN-UC). La identificación se realizó mediante el uso de las guías de campo GARCÍA et al. (2002) y VALENCIA et al. (2005), y por comparación con la colección de referencia del Museo de Historia Natural de la Universidad de Caldas (MHN-UC) con la revisión de Julián Salazar.
Se realizó una prueba de chi-cuadrado para determinar si hubo diferencias entre la producción de flores, de frutos, la riqueza, la abundancia y la diversidad de especies (índice de Shannon-Wiener) de mariposas entre los meses de estudio. La prueba de correlación de Spearman se usó para evaluar la relación entre la fenología reproductiva de I. ornata con la riqueza, la abundancia y la diversidad de mariposas. Los análisis se corrieron con el programa Statistica 5.1 (STATSOFT INC., 1998).
RESULTADOS Fenología reproductiva de Inga ornata
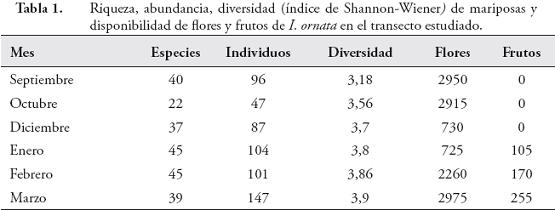
Los árboles de I. ornata presentaron dos picos de producción de flores durante los meses de septiembre-octubre (2005) y marzo (2006), con una baja producción de flores en los meses de noviembre a enero (Tabla 1). La fructificación solo ocurrió en los meses de enero a marzo con un pico de producción durante este último mes. La cantidad de flores disponibles por árbol varió entre 0 y 960 (× = 101,4±153,6) y la cantidad de frutos disponibles entre 0 y 100 (× = 4,7±8,6). La producción mensual de flores y de frutos presentó diferencias significativas entre meses (flores: X² = 2841,165 g.l = 5, p < 0,0001; frutos: X² = 658,362 g.l = 5, p < 0,0001).
Las flores de I. ornata son blancas con numerosos estambres que se unen en la base formando un tubo donde se acumula el néctar (Figura 1a). Las flores abren en la tarde y duran 24 horas, durante este periodo de tiempo secretan entre 150 y 850 ul de néctar con una concentración de azucares entre 16% y 17,6% y un contenido energético entre 139,68 y 1313,8 julios (MARÍN-GÓMEZ, obs. pers). El fruto maduro es una legumbre tomentosa de color ferruginoso de 19,5 a 51 cm de largo que cuando alcanza su máximo desarrollo empieza a abrir debido al crecimiento de la semilla y la presión del arilo (Figura 1b).

Riqueza y abundancia de mariposas en el potrero
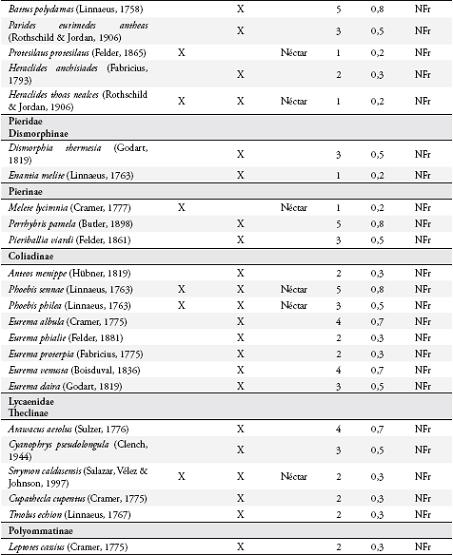
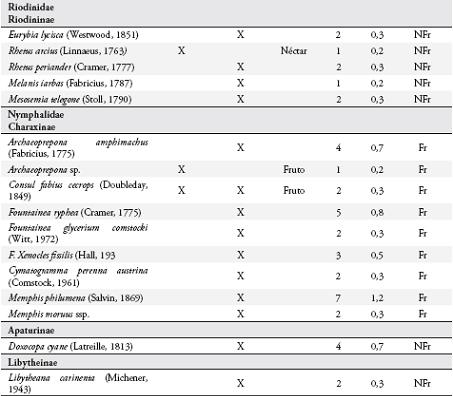
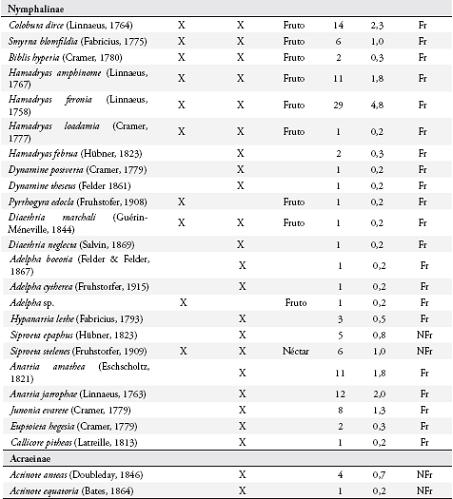
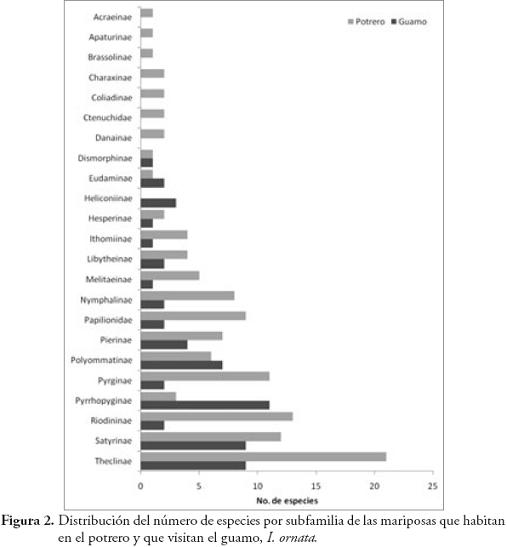
Se colectaron 585 individuos correspondientes a 116 especies (Anexo 1), con dominancia de la familia Nymphalidae (73 especies). En general, las subfamilias más representativas fueron Nymphalinae (20 especies y 159 individuos), Hesperiinae (13 especies y 20 individuos), Eudaminae (12 especies y 33 individuos), Pyrginae (11 especies y 15 individuos), Charaxinae (8 especies y 26 individuos) y Satyrinae (7 especies y 208 individuos) (Figura 2). Las especies dominantes fueron Magneuptychia tiessa (62 individuos), Euptychia polyphemus (59), E. hermes (49), E. mollis (45), Hamadryas feronia (29), Pareuptychia hesione (18) y Colobura dirce (14). Se colectó un mayor número de especies (96) con redes entomológicas y un mayor número de individuos (364) con trampas.
Mariposas asociadas al guamo
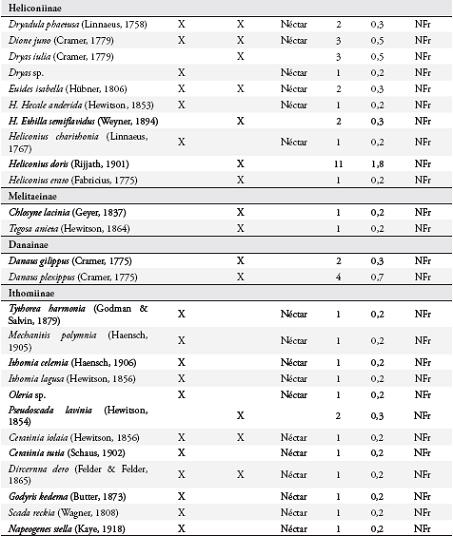
Se observaron 59 especies de mariposas visitando los árboles de I. ornata, 34 de estas reportadas también para el potrero. Catorce especies fueron atraídas desde hábitats circundantes al guamo: Ceratinia tutia, Godyris kedema, Ithomia lagusa, Scada reckia y Oleria sp., desde el bosque, guadual y guadual mixto; Pyrrhogyra edocla desde el bosque y guadual; Ithomia celemia desde el guadual mixto y guadual; Astraptes janeira, Hecale anderida, Heliconius charithonius, Mechanitis polymnia y Napeogenes stella desde el guadual mixto; Melete lycimnia y Protesilaus protesilaus desde el guadual. Además 11 especies no se reportan en el listado de la reserva, tres especies de Ctenuchidae y nueve de Nymphalidae: Dryas sp., Archaeoprepona sp., Adelpha sp., Vareuptychia austera, Tythorea harmonia, Rhetus arcius, Oressinoma typha y Dyrcernna dero.
Nymphalidae y Hesperiidae fueron grupos dominantes en el potrero y en los guamos, sin embargo hay diferencias notorias al comparar el número de especies por subfamilias entre estos dos elementos (Figura 2); Ithomiinae y Heliconinae presentaron un mayor número de especies en los guamos, pero estuvieron poco representadas en el potrero. Lo contrario ocurrió con el resto de las subfamilias que presentaron una mayor riqueza en el potrero (Figura 2).
Uso de recursos alimenticios por las mariposasSe observaron 42 especies consumiendo el néctar floral (Anexo 1), con dominancia de Nymphalidae (17) y Hesperiidae (14). Las subfamilias más representadas fueron: Ithomiinae (11), Eudaminae (9), Nymphalinae (8), y Heliconiinae (6). Los visitantes más comunes fueron Astraptes fulgerator, Urbanus proteus y Siproeta stelenes.
Las mariposas que visitaron las flores presentaron diferencias en su actividad de forrajeo diurno. Durante las primeras horas de la mañana (06:00-08:00 h) fueron más activos los Ithomiinae, a partir de este periodo y hasta el medio día aumentó la actividad de los Pieridae (Phoebis spp.), Papilionidae, Heliconiinae y Nymphalidae (Siproeta stelenes). Después de las 14:00 h la actividad de visitas a las flores fue poca y estuvo representada solo por Hesperiidae, que forrajearon durante todo el día con una mayor actividad en las ultimas horas de la tarde (16:00 a 18:00 h), periodo que coincide con la antesis floral.
Se registraron 17 especies de la familia Nymphalidae consumiendo la pulpa de los frutos (Anexo 1), principalmente al medio día, pertenecientes a las subfamilias: Nymphalinae, Satyrinae, Brasolinae y Charaxinae. Estas mariposas visitaron solo los frutos maduros abiertos, en donde forrajearon durante más de 2 minutos si no se perturbaban. Todos los Satyrinae y tres especies de Nymphalinae (Colobura dirce, Hamadryas feronia y H. amphione) fueron los visitantes más frecuentes. Cada especie forrajeó en forma independiente sobre un fruto, aunque se observó forrajeo simultáneo con algunas especies de dípteros (Sarcophagidae) y varios véspidos.
Relación entre la oferta de alimento y la diversidad de mariposasLa riqueza, la abundancia y la diversidad de especies aumentaron durante los últimos tres meses de estudio, coincidiendo con los picos de producción de frutos de I. ornata (Tabla 1), aunque solo se encontraron diferencias significativas en la abundancia de mariposas entre meses (X² = 53,25773 g.l = 5, p < 0,0001). Se encontró una relación positiva entre el número de frutos y la abundancia de mariposas (rs = 0,881, p = 0,021, n = 6) y entre el número de frutos y la diversidad de especies (rs = 0,95, p = 0,0048, n = 6). No se encontró relación al comparar el número de flores con la diversidad, la abundancia y la riqueza de especies. Tampoco se encontró una correlación positiva al comparar la diversidad, riqueza y abundancia de mariposas frugívoras y no frugívoras con la disponibilidad de frutos y flores, respectivamente.
DISCUSIÓNNymphalidae fue la familia con mayor riqueza en el potrero, resultado esperado ya que esta familia, junto con Lycaenidae y Hesperiidae, son los grupos de mariposas más ricos en el Neotrópico (BROWN, 1996). Se encontró una mayor representatividad de Nymphalidae, que la registrada por HERNÁNDEZ et al. (2003) en un sistema ganadero en Nicaragua. La riqueza de mariposas en el potrero estudiado puede estar relacionada a factores como el clima, altitud, estado de conservación, distancia a hábitat adyacentes (BROWN & FREITAS, 2000), o disponibilidad de recursos tales como la abundancia de flores y frutos de I. ornata, una especie común en la zona de estudio (MARÍN-GÓMEZ, obs. pers.), y que fue utilizada por el 50% de la comunidad de mariposas del potrero con mayor representación de mariposas no frugívoras.
Dentro del gremio de mariposas no frugívoras, Ithomiinae fue la subfamilia con mayor número de especies asociadas a I. ornata, mientras que en la parcela del potrero solo se reportaron tres especies. Este grupo se alimenta principalmente del néctar floral, están asociados a bosques poco intervenidos (DEVRIES et al., 1997; BROWN & FREITAS, 2000) y se desplazan a zonas abiertas en busca de néctar y otros recursos como heces de aves o plantas hospederas para el caso de la hembras (DRUMMOND & BROWN, 1987 RAY & ANDREWS, 1980; BECCALONI & GASTON, 1995; BECCALONI, 1997) y alcaloides como las pirrolizidinas, usadas como precursores de feromonas en los machos (BROWN, 1984; TRIGO et al., 1996). Aunque no se encontró una correlación entre el número de flores disponibles de I. ornata y la diversidad de Ithomiinae, sí se observó una clara tendencia al aumento de especies en la época de floración de esta planta.
Los Heliconiinae mostraron una tendencia similar a la observada en Ithomiinae, con un mayor número de especies asociadas a I. ornata y un menor número en el potrero. Al igual que muchos Ithomiinae, los Heliconiinae se desplazan desde el interior del bosque a zonas de crecimiento secundario en busca de flores (MALLET & GILBERT, 1995). Hesperiidae fue el segundo grupo con mayor número de especies visitando I. ornata, aunque fueron poco abundantes en el potrero. En general, se colectaron con jama y pocas especies con trampa, por lo que su abundancia puede estar sub-estimada, dado que son mariposas no frugívoras, de vuelo rápido y difíciles de capturar con jama. Las especies de Hesperiidae son altamente móviles y realizan migraciones altitudinales en busca de recursos florales y plantas hospederas (BROWN & FREITAS, 2000), y son importantes polinizadores de varias especies de plantas (KOPTUR, 1983; VALENCIA et al., 2005). Tampoco se encontró una correlación positiva entre el número de flores de I. ornata y la diversidad de esta familia en el potrero, aunque sí se observó una tendencia al aumento del número de especies, principalmente de la subfamilia Eudaminae.
Menos del 50% de los Nymphalinae registrados en el potrero, fueron observados en I. ornata, la mayoría pertenecientes al gremio de los frugívoros (sensu DEVRIES et al., 1997), a excepción de Siproeta epaphus y S. stelenes que se alimentaron exclusivamente de néctar y no fueron capturados en trampas. Nymphalinae y Charaxinae fueron los frugívoros con mayor número de especies y baja abundancia, Brassolinae estuvo pobremente representado y Satyrinae fue la subfamilia más abundante en el potrero, lo que podría relacionarse con la oferta de frutos. Estas cuatro subfamilias mostraron una tendencia al incremento de su diversidad durante los meses de mayor disponibilidad de frutos de I. ornata, aunque esta correlación no fue significativa. Esta tendencia también podría estar relacionada con la explotación de recursos alimenticios alternativos como carroña, charcas o suelos húmedos con alta concentración de minerales, abundancia de excretas de ganado y aves (ADLER & PEARSON, 1982; BECK et al., 1999; VALENCIA et al., 2005; HAMER et al., 2006).
A pesar de la alta riqueza de mariposas no frugívoras que visitaron las flores de I. ornata, tan solo tres especies fueron visitantes frecuentes (A. fulgerator, U. proteus y S. stelenes). En las especies de Inga estudiadas por KOPTUR (1983) en Costa Rica, los principales polinizadores fueron Sphingidae y Hesperiidae, mientras que en la Amazonia brasileña lo fue Heliconinae (FALCÃO & CLEMENT, 2000), estos datos concuerdan con los de este estudio a excepción de Sphingidae que no fue estudiado. Un resultado novedoso de este estudio es el consumo de frutos de árboles del género Inga por mariposas, algo no reportado en literatura. Dentro de este grupo los visitantes frecuentes fueron todos los Satyrinae y tres especies de Nymphalinae (C. dirce, H. feronia y H. amphione), habitantes frecuentes de bosques y no de potreros (DEVRIES et al., 1997; DEVRIES & WALLA, 2001), al parecer la oferta de frutos de los guamos es un atractivo para especies que utilizan este recurso. I. ornata es un árbol de floración continua que produce grandes cantidades de néctar y frutos, además su floración en masa atrae a una gran diversidad de mariposas y parece ser un recurso importante para estos organismos, principalmente para las no frugívoras.
AGRADECIMIENTOSA Germán Darío Gómez y al Centro de Investigaciones en Biodiversidad de la Universidad del Quindío por su apoyo logístico, y a Julián Salazar del Museo de Historia Natural de la Universidad de Caldas por su invaluable colaboración en la determinación de los ejemplares y revisión al manuscrito.
ADLER, P.H. & PEARSON, D.L., 1982.- Why do male butterflies visit mud puddles? Canadian Journal of Zoology, 60: 322-325. [ Links ]
ALZATE, H., 1994.- Patrimonio y deterioro ambiental. Informe del estado de los recursos naturales y medio ambiente en el Valle del Cauca. Contraloría del Valle, División de Recursos Naturales y Medio Ambiente, Cali, Colombia. 136p. [ Links ]
BECCALONI, G.W., 1997.- Vertical stratification of Ithomiine (Nymphalidae: Ithomiinae) mimicry complexes: the relationship between adult flight height and larval host-plant height. Biological Journal of the Linnean Society, 62: 313-341. [ Links ]
BECCALONI, G.W. & GASTON, K.J., 1995.- Predicting the species richness of Neotropical forest butterflies: Ithominae (Lepidoptera: Nymphalidae) as indicators. Biological Conservation, 71: 77-86. [ Links ]
BECK, J., MÜHLENBERG, E. & FIEDLER, K., 1999.- Mud-puddling behaviour in tropical butterflies. Tropical Lepidoptera, 6: 5-9. [ Links ]
BROWN, K.S.JR., 1984.- Chemical ecology of dehydropyrrolizidine alkaloids in adult Ithomiinae (Lepidoptera: Nymphalidae). Rev. Bras. Biol., 44: 435-460. [ Links ]
________., 1996.- Conservation of threatened species of Brazilian butterflies. International symposium on Butterfly Conservation III: 45-62. Japan. [ Links ]
BROWN, K.S.JR. & FREITAS, A.V., 2000.- Atlantic forest butterflies: Indicators for landscape conservation. Biotropica, 32 (4b): 934-956. [ Links ]
DEVRIES, P.J., MURRAY, D. & LUNDE, R., 1997.- Species diversity in vertical, horizontal and temporal dimensions of a fruit-feeding butterfly community in an Ecuatorian Rainforest. Biological Journal of the Linnean Society, 62: 313-341. [ Links ]
DEVRIES, P.J. & WALLA, T.R., 2001.- Species diversity and community structure in Neotropical fruitfeeding butterflies. Biological Journal of the Linnean Society, 74: 1-15. [ Links ]
DRUMMOND III, B.A. & BROWN, K.S.JR., 1987.- Ithomiinae (Lepidoptera: Nymphalidae): summary of known larval foodplants. Annals of Missouri Botanical Garden, 74: 341-358. [ Links ]
ETTER, A., 1998.- Mapa general de ecosistemas de Colombia (1:11.500.000) (en) CHÁVEZ, M.E., ARANGO, N. (eds.) Informe Nacional sobre el Estado de la Biodiversidad. Vol. 3. Instituto Alexander von Humboldt, Programa de las Naciones Unidas para el Medio Ambiente y Ministerio de Medio Ambiente, Bogotá, Colombia. [ Links ]
FALCÃO, M.A. & CLEMENT, C.R., 2000.- Fenologia e produtividade do ingá cipó (Inga edulis) na Amazônia Central. Acta Amazónica, 30 (2): 173-180. [ Links ]
FAGUA, G., 1996.- Comunidades de mariposas y artropofauna asociada al suelo de tres tipos de vegetación de la serranía de Taraira (Vaupes, Colombia): Una prueba del uso de mariposas como bioindicadores. Revista Colombiana de Entomología, 22 (3): 143-151. [ Links ]
Farfán, V.F. & Mestre, A.M., 2004.- Manejo del sombrío y fertilización del café en la zona central Colombiana. Avances técnicos Cenicafé, 330: 1-8. [ Links ]
GARCÍA, R.C., ARMBRETCHT, I. & ULLOA-CHACÓN, P., 2001.- Staphylinidae (Coleoptera): composición y mirmecofilia en bosques secos relictuales de Colombia. Folia Entomológica Mexicana, 40 (1): 1-10. [ Links ]
GARCÍA, C., CONSTANTINO, L.M., HEREDIA, M.D. & KATTAN, G., 2002.- Mariposas Comunes de la Cordillera Central de Colombia. Wildlife conservation society. Cali, Colombia. [ Links ]
HALL, J.P. & WILLMOTT, K.R., 2000.- Patterns of feeding behaviour in adult male riodinid butterflies and their relationship to morphology and ecology. Biological Journal of the Linnean Society, 69: 1-23. [ Links ]
HAMER, K.C., HILL, J.K., BENEDICK, S., MUSTAFFA, N., CHEY, V.K. & MARYATI, M., 2006.- Diversity and ecology of carrion and fruit-feeding butterflies in Bornean rain forest. Journal of Tropical Ecology, 22: 25-35. [ Links ]
HERNÁNDEZ, B., MAES, J.M., HARVEY, C.A., VÍLCHEZ, S., MEDINA, A. & SÁNCHEZ, D., 2003.- Abundancia y diversidad de escarabajos coprófagos y mariposas diurnas en un paisaje ganadero en el departamento de Rivas, Nicaragua. Agroforestería en las Américas, 10 (39-40): 1-10. [ Links ]
HORNER, M.C., DAILY, G.C., EHRLICH, P.R. & BOGGS, C.L., 2003.- Countryside Biogeography of Tropical Butterflies. Conservation Biology, 17 (1): 168-177. [ Links ]
Koptur, S., 1983.- Flowering phenology and floral biology of Inga (Fabaceae: Mimosoideae). Systematic Botany, 8 (4): 354-368. [ Links ]
MALLET, J. & GILBERT, L.E., 1995.- Why are there so many mimicry rings? Correlations between habitat, behavior and mimicry in Heliconius butterflies. Biological Journal of the Linnean Society, 55: 159-180. [ Links ]
PENNINGTON, T.D., 1997.- The genus Inga Botany. The Royal Botanical Gardens. London, England. 490p. [ Links ]
RAY, T.S. & ANDREWS, C.C., 1980.- Antbutterflies: butterflies that follow army ants to feed on ant-bird droppings. Science, 210: 1147-1148. [ Links ]
STATSOFT, INC., 1998.- Statistica for Windows. Tulsa. http://www.statsoft.com [ Links ]
TRIGO, J.R., BROWN, K.S.JR., SILVANA, A.H. & SOARES, L.E., 1996.- Qualitative patterns of pyrrolizidine alkaloids in Ithomiinae butterflies. Biochemical Systematics and Ecology, 24 (3): 181-188. [ Links ]
VALENCIA, C.A., GIL, Z.N. & CONSTANTINO, L.M., 2005.- Mariposas diurnas de la zona central cafetera de Colombia. Guía de Campo. Cenicafé, Chinchiná, Colombia. 244p. [ Links ]