










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.16 no.1 Manizales Jan./June 2012
STUDY OF SPIDERS' DIVERSITY IN A TROPICAL DRY FOREST (BS-T) IN SABANALARGA, ATLÁNTICO, COLOMBIA
* FR: 14-II-2012. FA: 22-VI-2012.
1 Bióloga. Grupo Biodiversidad del Caribe Colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia. Email: ry.escorciag@gmail.com.
2 M.Sc. Docente investigador. Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia. Email: neyjosemartinez@gmail.com.
3 Estudiante de pregrado. Semillero de Investigación Insectos (NEOPTERA) del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia.
El presente trabajo, muestra como se evaluó la composición y estructura, de la araneofauna en la Reserva Campesina La Sierra (RCS), municipio de Sabanalarga, departamento del Atlántico, Colombia. Se realizaron cinco muestreos, entre agosto y diciembre de 2009. Se escogieron tres hábitats: Cultivo (Cul), Interior de Bosque (I. B) y Borde de bosque (Bor). Se recolectaron 879 individuos, agrupados en 29 familias y 68 morfoespecies. La familia Araneidae, presentó mayor riqueza (23) y abundancia (433); destacándose, la morfoespecie Araneus sp2, con 30 individuos. El hábitat de I. B, presentó la mayor riqueza (55) y abundancia (334); mientras que, en diciembre, se capturó el mayor número de morfoespecies (24) y de individuos (300). El gremio de las arañas Cursoriales en Suelo (CS), presentó mayor riqueza (9) y Constructoras de telas orbiculares (CTO), la mayor abundancia (504). A pesar de comprender un área relativamente pequeña, la RCS, arrojó un alto porcentaje de las familias reportadas para el país, demostrando, que este remanente de bosque, aún mantiene una alta diversidad de arañas.
Palabras clave: Araneidae, Araneus sp1, composición, constructoras de telas orbiculares, gremios, RCS.
AbstractThis work shows how the composition and structure of arachnofauna in the Rural Reserve la Sierra (RCS in Spanish), Sabanalarga municipality, department of Atlántico, Colombia was evaluated. Five samplings were collected between August and December 2009. Three habitats were selected: Farming (Farm), Interior Forest (I.F) and Forest Edge (FE). A total of 879 individuals grouped in 29 families and 68 morphoespecies were collected. The Araneidae family presented the greatest richness (23) and abundance (433). The IF presented the greatest richness (55) and abundance (334), while in December, the greatest number of morphospecies (24) and individuals (300) were captured. The group of Cursorial Spiders (CS) presented greater richness (9) and the greatest abundance in Orbicular Web Construction (OWC), 504. In spite of covering a relatively small area, the RCS produced a high percentage of the families reported for the country, demonstrating that this forest still keeps a high diversity of spiders.
Key words: Araneidae, Araneus sp1, composition, OWC, guilds, RCS.
En Colombia, la mayor parte del bosque seco tropical (Bs-T), se encuentra en la planicie costera de la región Caribe (IAVH, 1998). Esta formación boscosa, se define como aquella formación vegetal, que presenta una cobertura boscosa continua y que se distribuye, entre los 0-1000 m de altitud; presenta temperaturas superiores a los 240 ºC (piso térmico cálido) y precipitaciones, entre los 700 y 2000 mm anuales, con uno o dos periodos marcados de sequía al año (MURPHY & LUGO, 1986; IAVH, 1997). En la actualidad, esta formación boscosa en la región, está aislada en fragmentos inmersos, en una matriz compuesta de cultivos, potreros y cercas vivas, lo que genera variaciones en la composición y estructura de la vegetación y altera, la disponibilidad de alimento (AGUILERA & CASANUEVA, 2005), generando cambios en la composición de las comunidades de arañas entre diferentes hábitats y microhábitats.
Las arañas (orden Araneae), comprenden uno de los grupos más comunes de animales, debido a que, cuenta con la capacidad de prosperar en diversos hábitats (MANI, 1968; TURNBULL, 1973) siendo considerados, como uno de los grupos más diversos de la tierra, ubicándose en el séptimo lugar de los órdenes de animales (CODDINGTON & LEVI, 1991) con 42.751 especies, agrupadas en: 3.859 géneros y 110 familias (PLATNICK, 2012). Además, este grupo de artrópodos, es un grupo focal que puede aportar información ecológica, sobre la dinámica de los fragmentos de bosque seco y los hábitats adyacentes, debido a la sensibilidad, a pequeños cambios en la estructura de los hábitats (RICO et al., 2005).
En Colombia, han sido reportadas 680 especies, agrupadas en: 249 géneros y 49 familias (FLÓREZ, 1995). Sin embargo, se espera que este número, sea mayor ante la escasez de estudios en algunas regiones biogeográficas del territorio colombiano, como es el caso, del Cinturón Árido Pericaribeño, donde predomina la zona de vida de Bs-T (HOLDRIDGE, 1967).
Teniendo en cuenta lo anterior, se evaluó la composición y estructura de la araneofauna, en un fragmento de Bs-T, en la Reserva Campesina La Sierra (RCS), del departamento del Atlántico, Colombia, describiendo la diversidad, abundancia y composición faunística en ésta, realizando comparaciones entre hábitats y épocas de muestreo y clasificando las arañas por gremios; todo esto, con el fin de contribuir al conocimiento de las arañas en la región Caribe.
MATERIALES Y MÉTODOSÁrea de estudio
La Reserva Campesina La Sierra (RCS), está localizada en el corregimiento La Peña, en el municipio de Sabanalarga, departamento del Atlántico, Colombia (Figura 1). Está ubicada a 10º35'43,7"N y 74º59'81,5"O, a una altitud de 172 m. La temperatura promedio anual en la zona, es de 27,8 ºC, con una precipitación anual de 1.021 mm y una humedad relativa, que oscila entre 62 y 92 % por mes.
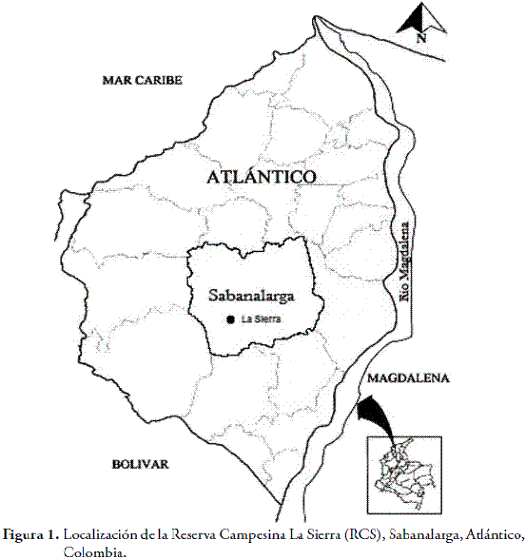
El área de estudio forma parte de la región biogeográfica del Cinturón Árido Pericaribeño (HERNÁNDEZ et al., 1992). Por sus características bioclimáticas, la región de estudio, corresponde al bosque seco tropical (HOLDRIDGE, 1967). La RCS, posee una extensión de 34 ha, de las cuales, un tercio ha sido deforestada para las actividades antrópicas de siembra de cultivos, ganadería y extracción de madera, principalmente.
Diseño de muestreoEl presente estudio, se realizó entre agosto y diciembre de 2010, con salidas mensuales de tres días. Los muestreos se realizaron en jornadas diurnas, entre las 8:00-12:00 y las 13:00-16:00 horas. En la RCS, se escogieron tres hábitats, que estaban distanciados, aproximadamente 150 m, uno del otro, presentando, condiciones vegetales y de conservación, muy contrastantes. El primero, fue el Interior de Bosque (I. B), con zonas de vegetación original y acumulación, de demasiada hojarasca; con parches, donde se evidencia, los procesos de tala selectiva con pérdida moderada de dosel, originando rastrojos con vegetación secundaria, que confiere un paisaje actual en mosaico. El fragmento de Borde de bosque (BOR), localizado en un sendero que recorría el borde del fragmento, donde se observó, el tránsito de campesinos y ganado vacuno. Este lugar presentaba muchos arbustos, enredaderas y especies vegetales propias del fragmento de bosque. El tercero, se ubicó en la parte de Cultivo (CUL), donde se observó, la siembra de plátano (Musa sp.) y arboles dispersos de guayaba (Psidium guajava L.), mango (Manguifera indica L.) y papaya (Carica papaya L.). Los tres hábitats, se encuentran atravesados por una quebrada, que influye en la estructura y composición vegetal.
En cada hábitat, se delimitó un transecto de 100 x 4 m, donde se marcaron cinco estaciones de muestreo equidistantes 20 m. En cada estación, se realizaron las siguientes técnicas de captura: i) agitación de follaje, ii) cernido de hojarasca y iii) captura manual. Además, los muestreos se complementaron con 10 trampas de caída, distribuidas uniformemente, a lo largo del transecto, que permanecieron en campo, durante 24 horas. Cada trampa, consistió en un vaso plástico con alcohol, al 70 %, a un tercio de su volumen. En cada muestreo, se tomaron los datos de precipitación, con la ayuda de un pluviómetro.
Los ejemplares recolectados, se preservaron en alcohol etílico al 70 % y se trasladaron al laboratorio de Zoología de la Universidad del Atlántico, donde se identificaron a nivel de familia, con las claves de JÓQUE & DIPPENAAR (2007) y las claves, a nivel de género o especie de OPEL (1979), CODDINGTON (1986), LEVI (2002) y AGUILERA & CASANUEVA (2005). Para el caso colombiano, los inventarios de especies, son aún incipientes y están basados, principalmente, en la literatura (GILEDE- MONCAYO & BELLO-SILVA, 2000), por esta razón, cuando no fue posible la identificación, a nivel de especie, se emplearon unidades taxonómicas reconocibles (morfoespecies), siguiendo las recomendaciones de OLIVER & BEATTIE (1996). Una vez identificados los especímenes, se incorporaron a la colección de arañas en húmedo, del Museo de la Universidad del Atlántico (UARC) y al Instituto de Ciencias Naturales (ICN), de la Universidad Nacional de Colombia.
Análisis de datosSe estimó la riqueza y abundancia de familias y morfoespecies, de arañas por hábitat y muestreo. Con el fin de observar, la efectividad de los métodos de captura y el esfuerzo muestral en la RCS, se realizó una curva de acumulación para las morfoespecies encontradas, con los estimadores de riqueza no paramétricos ICE, Chao 2 y Bootstrap con ayuda del programa EstimateS 8.2 (COLWELL, 2009). La diversidad alfa, en cada hábitat, se estimó calculando los números de la serie de Hill (N1 y N2) y el índice de equidad de Pielou (J). La diversidad beta (o tasa de recambio de especies), entre hábitats, se calculó con el Índice de complementariedad (I. C), propuesto por COLWELL & CODDINGTON (1994). La descripción de estos índices, se encuentran en VILLAREAL et al. (2004). Para determinar, si existen diferencias significativas en la riqueza y abundancia de arañas y en la estructura de la comunidad, entre los tres hábitats, se aplicó la técnica de ANOSIM. La matriz de similitud, se calculó utilizando el índice de Bray-Curtis. Considerando a las especies, que aportaron más del 10 % de la abundancia total, se determinó las morfoespecies, que tipificaron o caracterizaron los hábitats (análisis SIMPER). Adicionalmente, con el fin de estimar la riqueza y abundancia de los gremios de arañas en la RCS, estas fueron agrupadas en gremios, escogidos según la conducta adoptada por el depredador, en relación con la presa (LILJESTHRÖM et al., 2002).
RESULTADOS Y DISCUSIÓNConsideraciones generales sobre la araneofauna de la Reserva Campesina de La Sierra
La presente contribución, constituye el primer estudio sobre las comunidades de arañas en fragmentos de bosque seco tropical (Bs-T), en las zonas bajas del Caribe colombiano. En la RCS, se capturaron 879 individuos, distribuidos en 68 morfoespecies y 29 familias (Tabla 1). El esfuerzo de muestreo realizado, fue suficiente para inventariar el 90,72 %, de la riqueza teórica máxima esperada, según el estimador ICE; el 90,42 %, según Chao 2 y el 84,21 %, según el estimador Bootstrap (Figura 2). Además, se observa que, la distancia entre los estimadores y la curva que representa el conjunto de los valores de riqueza observada es pequeña, lo que indica, que las estimaciones de riqueza, son consistentes (CODDINGTON, 2001). Sin embargo, el hecho de que, aproximadamente, un tercio de la riqueza observada, corresponda a especies raras (12 encontradas únicamente, en un sitio y 11, sólo en dos sitios), hace que los inventarios no sean completos, lo cual es de esperarse, en ecosistemas tropicales, donde las especies dominantes, son pocas en comparación con la gran cantidad de especies raras (DEZA & ANDÍA, 2009).
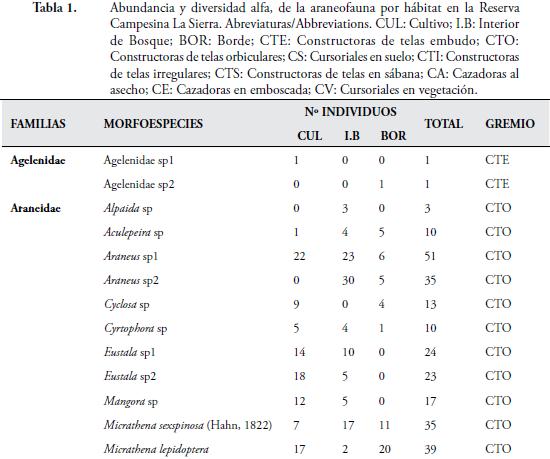

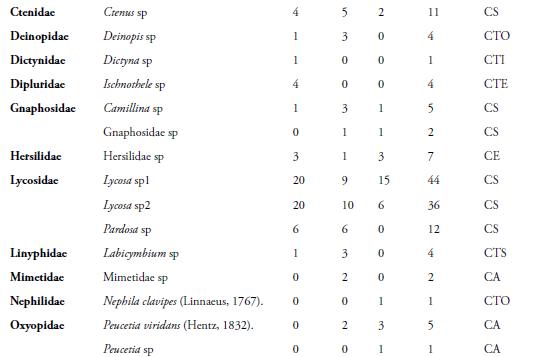
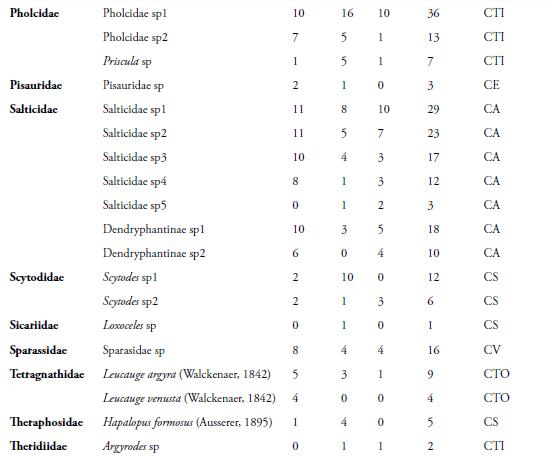

Diversos trabajos realizados en bosques tropicales, han demostrado también, un alto número de especies raras en términos de abundancia, con sólo un individuo (FLÓREZ, 1998; SILVA & CODDINGTON, 1996; DEZA & ANDÍA, 2009). A nivel de familia, en este estudio, se capturaron el 59,2 %, del total registradas en Colombia por FLÓREZ (1995). El número de familias de arañas, es mayor que el reportado por FLÓREZ (1999), quien capturó 20 familias, en un bosque muy seco en la localidad de Loboguerrero, Valle del Cauca, Colombia. Lo anterior indica, que este remanente de Bs-T, mantiene una fauna representativa de la araneofauna presente en las tierras bajas del Caribe colombiano, lo que demuestra, la importancia de estos fragmentos como reservorio de la biodiversidad en un agropaisaje.
De las 29 familias registradas, tres pertenecen al suborden Mygalomorphae (Barychelidae Dipluridae y Theraphosidae) y 26, al suborden Araneomorphae. El suborden Mygalomorphae, estuvo representado por sólo 10 individuos, en los tres hábitats, lo cual puede atribuirse, al corto tiempo de muestreo. Adicional a esto, los efectos de la intervención humana en la zona, pueden estar incidiendo, en la supervivencia de las especies de tarántulas, disminuyendo las poblaciones de este grupo de arañas.
La familia Araneidae, fue la más diversa (23 morfoespecies) y abundante (433 individuos), seguida de Salticidae, con siete morfoespecies y 112 individuos (Tabla 1). Los araneidos, son una familia muy diversa y ocupa, casi todos los estratos arbóreos y una gran cantidad de nichos ecológicos. Además, esta familia, es considerada una de las más cosmopolitas y de alta riqueza genérica (CODDINGTON & LEVI, 1991). Estos datos concuerdan, ampliamente, con los resultados obtenidos por distintos autores en el neotrópico, como: FLÓREZ (1998), RICO et al. (2005), CABRA et al. (2010) y por FERREIRA-OJEDA et al. (2009), en un bosque húmedo subtropical de La Sierra Nevada de Santa Marta, Colombia.
Por otro lado, las familias Barychelidae, Caponiidae, Dictynidae, Nephilidae, Philodromidae y Sicariidae, presentaron la menor riqueza, con una sola morfoespecie y la menor abundancia, con tan solo un individuo (Tabla 1). Este hecho, se debe posiblemente, a que las arañas de suelo, como las pertenecientes a la familia Barychelidae, Caponiidae y Sicariidae, aprovechan a manera de microhábitat, los troncos podridos de los árboles (JIMÉNEZ et al., 2010), por lo cual, es muy difícil su visualización y captura.
La morfoespecie con mayor abundancia fue Araneus sp1 (Araneidae), con 51 individuos, seguida por Lycosa sp1 (Lycosidae) y Micrathena sp (Araneidae), con 44 y 42 individuos, respectivamente (Tabla 1). Es posible, que la abundancia de Araneus sp1 y Micrathena sp, se deba, a que los juveniles o los huevos, estén latentes, durante la época de lluvias y luego eclosionen, al llegar la época seca. La gran abundancia de Lycosa sp1, se debe posiblemente, a las técnicas de muestreo utilizadas, debido a que las trampas de caída y el cernido de hojarasca, son métodos eficientes para este tipo de araña, que presenta una actividad epígea (RUBIO, 2007).
Debido al bajo nivel de exploración, realizado a nivel del Bs-T, del departamento del Atlántico y a que este estudio, es una primera aproximación a la araneofauna de esta región, las posibilidades de comparación y comprensión de estos patrones de diversidad de géneros y morfoespecies, se ven limitadas, sin embargo, en este trabajo, es posible discutir y establecer relaciones, con otros lugares de Colombia, donde existe este tipo de bosque.
Diferencias entre hábitats y épocas de muestreoEntre los tres hábitats estudiados, la mayor riqueza específica, a nivel de familia (23) y morfoespecie (55), se capturó en I. B; mientras que, el menor número de familias (20) y morfoespecies (44), se presentó en BOR (Tabla 1). Con respecto a la abundancia, el mayor valor, se presentó en el I. B, con 334 individuos (38,0 %) y en BOR, se capturó la menor con 218 individuos (24,8 %), del total recolectado (Tabla 1, Figura 2). Esto se debe, a que el I. B, posee una alta complejidad estructural en cuanto a la vegetación, además por su alta densidad boscosa y gran cantidad de microhábitats, es el hábitat propicio para muchas especies de arañas. Asimismo, durante los muestreos, se observó en este hábitat, una gran proporción de hojarasca que proporciona una cantidad de microclimas, que sirven para que muchas arañas, se protejan de las fluctuaciones de temperatura externa y la desecación (FOELIX, 1982). Estos factores y la provisión de presas, reducen la depredación interespecífica y permiten, la persistencia de especies raras (UETZ, 1976). El BOR, a pesar de ser un hábitat abierto para tejer telas, corresponde a un sendero por el cual, transita habitualmente, ganado vacuno, convirtiéndolo, en un hábitat altamente antropizado. Esto demuestra, que las arañas poseen una alta sensibilidad a los cambios en la estructura del hábitat (NEW, 1999). La morfoespecie mas abundante en I. B, fue Araneus sp2, con 30 individuos (Tabla 1). Esto se debe posiblemente, a que ubica su tela de gran tamaño y adherencia, en sitios abiertos del bosque, donde puede capturar un mayor número de presas. En BOR, Micrathena lepidóptera, con 20 individuos y Micrathena clypeata, con 17 individuos, fueron las morfoespecies más abundantes (Tabla 1). Las telas orbiculares construidas por estas especies, son ubicadas en áreas abiertas con bastante flujo del viento, como es el hábitat BOR. Araneus sp1 (22), fue la morfoespecie más abundante en el hábitat CUL (Tabla 1); la cual puede estar aprovechando la distancia, entre las plantas de Musa sp, sembradas en este hábitat para construir sus redes en lugares frescos, por donde circulan muchos insectos fitófagos, que le sirven como presas. Además, las hojas anchas de esta especie vegetal, pueden ser utilizadas por estas arañas como refugio (ARMENDANO et al., 2009) para protegerse de posibles depredadores y de los cambios bruscos de temperatura, que se presentan en la zona durante el día.
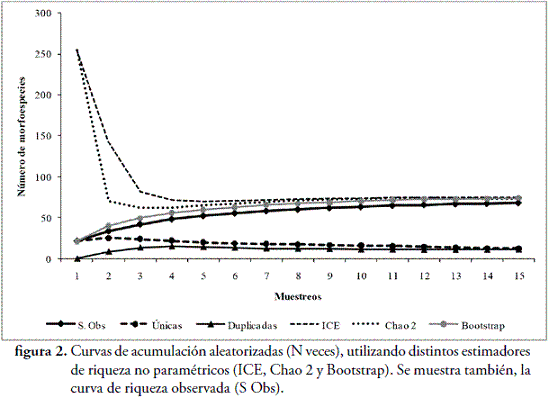
Con respecto a la diversidad, el mayor valor del número de Hill N1 (36,59), se presentó en I. B y el menor (30,19), n BOR. Para N2, el mayor valor (27,69), se calculó en CUL y el menor (23,6), en BOR (Tabla 1). Estos valores de diversidad, son bastante altos, demostrándose, que el número de especies comunes (N1) y muy comunes (N2), contribuyen de manera efectiva, a la diversidad del área. El índice de equidad de Pielou (J'), fue muy similar entre los tres hábitats, presentándose el mayor valor (0,9067), en CUL y el menor (0,8983), en I. B (Tabla 1). Los resultados de este índice, son altos, lo que puede estar indicando, que la composición de arañas, es bastante diversa en los tres hábitats escogidos, por esto, de ninguna manera, no hay una morfoespecie dominante y la mayoría de especies exclusivas, están representadas por pocos individuos.
Con respecto a los meses de muestreo, diciembre, presentó el mayor número de familias (24) y la mayor abundancia (300), mientras que, en noviembre, se capturó el menor número de familias (10) y la menor abundancia, con 112 individuos (Figura 3). Esto se debe, a que en diciembre, había finalizado el periodo de lluvias y la artropofauna tropical, varía sus densidades, debido a la disponibilidad de las presas, según la fenología del bosque (LEVINGS & WINDSOR, 1984; FLÓREZ, 1997). Además, en los meses de lluvia, las hembras de arañas, inician su reproducción y en diciembre, finaliza la puesta de sus huevos, lo cual incrementa su abundancia (Obs. per). A nivel de morfoespecies, la mayor riqueza (44) y abundancia (141) para CUL, se presentó en diciembre. Para este mismo mes, se capturó la mayor riqueza (29) y abundancia (88), en el BOR (Figura 3). En el I. B, la mayor riqueza, se presentó en diciembre (28), mientras que la mayor abundancia (82), se presentó en septiembre. Este hecho, puede estar relacionado, con la fenología del bosque y con los niveles de hojarasca, factores determinantes, en la incidencia de los artrópodos (LEVINGS & WINDSOR, 1984).
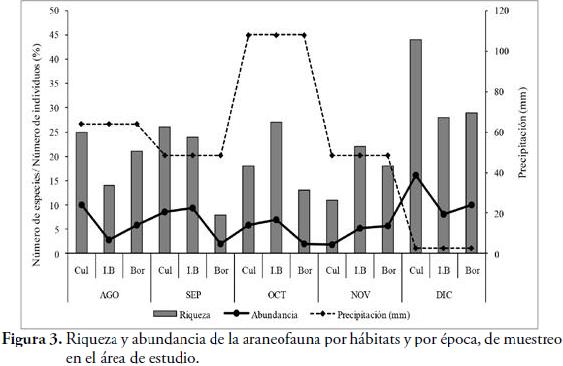
Los tres hábitats, comparten 28 morfoespecies (41,2 %) de las 68 capturadas. El Índice de Complementariedad (I. C), presentó el mayor valor entre CUL y BOR (0,97), compartiendo tres morfoespecies (Cyclosa sp, Parawixia sp1 y Dendryphantinae sp2). El segundo valor, lo presentó I. B, con BOR (0,91), con ocho morfoespecies compartidas (Araneus sp2, Micrathena schreibersii, Micrathena clypeata, Gnaphosidae sp, Peucetia viridans, Salticidae sp5, Cryptachaea sp). El menor valor de I. C (0,86), se presentó entre I. B, con CUL, con 13 morfoespecies compartidas (Eustala sp1, Eustala sp2, Mangora sp, Pronous sp, Verrucosa sp, Clubionidae sp2, Deinopis sp, Pardosa sp, Pisauridae sp, Scytodes sp1, Hapalopus formosus, Uloborus sp2). Esto demuestra, que la tasa de recambio, en la composición de especies de la zona, es bastante alta y que existe, una fauna de arañas propias para cada hábitat. A pesar de las distintas técnicas de captura empleadas y de los tres hábitats muestreados, existe un alto número de Singletons y especies únicas, debido a la alta diversidad de artrópodos, que posee un bosque seco tropical. Estos resultados, se corroboran con los reportados por FLÓREZ (1998), SILVA & CODDINGTON (1996) y AVALOS et al. (2009b).
Con el test ANOSIM, se determinó que la estructura de la comunidad de arañas en los tres hábitats escogidos en la RCS, no presentaron diferencias significativas (R = -0,088; n = p= 0,85). Esto probablemente, se debe a que la distancia existente, entre un hábitat y otro, era muy pequeña por el tamaño reducido de la reserva.
La similitud de los hábitats, se debe, a que el aporte realizado, a la abundancia por medio de las morfoespecies Eustala sp2 (10,42 %) y Araneus sp1 (9,98 %) en el CUL y Lycosa sp1 en el I. B (10,45 %) y en BOR (13,93 %), es muy bajo, tal como lo demuestra el análisis SIMPER (Tabla 2). Los porcentajes de similitud promedio, demuestran que, las morfoespecies encontradas, no tipifican cada hábitat, por lo que la estructura de la comunidad de arañas en la zona, es la misma.
GremiosDesde el punto de vista trófico, se capturaron familias pertenecientes a ocho gremios (Tabla 1, Figura 4), lo que demuestra, la alta complejidad del ensamblaje de este grupo de artrópodos en la zona. La riqueza de gremios parece causar un efecto sobre el emplazamiento de las telas y el aumento en la disponibilidad de sustratos donde fijarlas (SAMU & SZINETÁR, 2002), mostrando, la diversidad de formas que adquieren las arañas para obtener sus recursos, causando una disminución en la competencia entre ellas. LILJESTRHRÖM et al. (2002), señalan que, la diversidad de gremios presentes en el área, demuestra el efecto de las arañas en el control de las poblaciones de insectos y otros artrópodos; lo cual es importante, debido al aumento de insectos plaga, en los cultivos y es posible, que algunas de las morfoespecies presentes en la zona, puedan ser utilizadas como biocontroladoras.

El gremio de las arañas Cursoriales en Suelo (CS), fue el que presentó, una mayor riqueza en los tres hábitats (Figura 4A), debido probablemente, a que en la zona, había una gran cantidad de hojarasca y pasto seco, lo cual puede proveer, una mayor cantidad de microhábitats para las arañas (JIMÉNEZ et al., 2010). Por otro lado, los gremios Constructoras de Telas Sábana (CTS) y Cursoriales en Vegetación (CV), presentaron menor riqueza, con solo una familia de las 29 totales (3,4 %, cada uno). El gremio de Constructoras de Tela Embudo (CTE), no aparece en I. B, debido posiblemente, a que las arañas, no encuentran un refugio seco; esta zona, se encontraba influenciada por una quebrada ubicada más cerca del transecto, que en las otras dos zonas, que inunda los espacios, donde pueden habitar algunas migalomorfas, que pertenecen a este gremio.
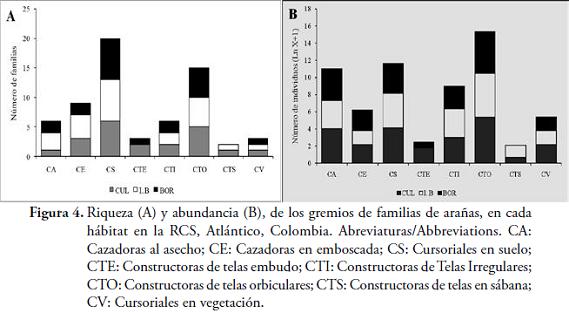
En cuanto a la abundancia, el gremio de Constructoras de Telas Orbiculares (CTO), presentó el mayor valor (504 individuos), con una distribución casi equitativa para los tres hábitats (Figura 4B). Estos resultados evidencian la tendencia general, en los bosques neotropicales (FLÓREZ, 1998; SILVA & CODDINGTON, 1996; RICO et al., 2005; CABRA et al., 2010), donde este gremio, exhibe una estrecha relación con la estructura de la vegetación. Adicional a esto, en el gremio de la CTO, se encuentra la familia Araneidae, la cual contribuyó, con 433 individuos, a la abundancia de este gremio. Estos resultados, no coinciden con los reportados por FLÓREZ (1999), quien realizó un trabajo de estructura y composición de arañas, en un bosque muy seco tropical de Colombia, donde predominaron los gremios de arañas tejedoras de telas irregulares (CTI) y CV, relegando a un tercer plano a las CTO. Posiblemente, la cantidad de arbustos de baja altura y mucha frondosidad, ofrecen una disponibilidad adecuada de sustratos, que favorezcan el tejido de telas orbiculares.
AGRADECIMIENTOSA toda la comunidad de la Reserva Campesina La Sierra (RCS), por su colaboración con la fase de campo. A Humberto Junior Maldonado Santana y Zuley Isabel Turizo Vargas, quienes nos colaboraron en la fase de campo y al resto de los miembros del semillero: NEÓPTERA, de la Universidad del Atlántico. Al profesor Eduardo Flórez, por su colaboración, con la identificación de algunos especímenes y por facilitarnos su bibliografía. A la Facultad de Ciencias Básicas de la Universidad del Atlántico, por facilitar los permisos de recolecta y por permitirnos utilizar, el Laboratorio de Zoología para la determinación taxonómica de las arañas.
AGUILERA, M. A. & CASANUEVA, M. E., 2005.- Arañas chilenas: estado actual del conocimiento y clave para las familias de Araneomorphae. Gayana, 69 (2): 201-224. [ Links ] [ Links ]
AVALOS, G., DAMBORSKY, M. P., BAR, M., OSCGHEROY, E. & PORCEL, E., 2009b.- Composición de la fauna de Araneae (Arachnida) de la Reserva provincial Iberá, Corrientes, Argentina. Rev. Biol. Trop., 57 (1-2): 339-351. [ Links ] [ Links ]
CODDINGTON, J., 1986.- The genera of the spider family Theridiosomathidae. Smithsonian contributions to zoology. Number 422. City of Washington. [ Links ] [ Links ]
CODDINGTON, J. A., 2001.- Estimación de la biodiversidad desde la escala local a la filogenética. Memorias. Primer Congreso Colombiano de Zoología. ICN. Bogotá [ Links ]. [ Links ]
COLWELL, R. K. & CODDINGTON, A., 1994.- Estimating the extent of terrestrial biodiversity througli extrapolation. Phil. Trans. R. Soc. London, (Ser. 6). 345: 101-118. [ Links ] [ Links ]
FERREIRA-OJEDA, L., FLÓREZ, E. & SABOGAL-GONZÁLEZ, A., 2009.- Arañas orbitelares de un Bosque Húmedo Subtropical de la Sierra Nevada de Santa Marta (Magdalena, Colombia). Caldasia, 31 (2): 381-391. [ Links ] [ Links ]
FLÓREZ, E., 1997.- Estudio de la comunidad de arañas del bosque seco tropical en la estación Biológica El Vínculo. Cespedesia, 22 (69): 37-57. [ Links ] [ Links ]
FLÓREZ, E., 1999.- Estructura y composición de una comunidad de arañas de un bosque muy seco tropical de Colombia. Boletín de Entomología Venezolana, 14 (1): 37-51. [ Links ] [ Links ]
GILEDE-MONCAYO, O. & BELLO-SILVA, J. C., 2000.- La familia Araneidae (Araneoidea: Orbicularie) en el departamento del Meta, Colombia. Biota Colombiana 1, (1): 125-130. [ Links ] [ Links ]
HOLDRIDGE, L. R., 1967.- Life Zone Ecology. Tropical Science Center, San José, Costa Rica. 206 p. [ Links ]
INSTITUTO ALEXANDER VON HUMBOLDT, IAVH., 1997.- Caracterización ecológica de cuatro remanentes de Bosque Seco tropical de la región Caribe colombiana. Grupo de Exploraciones Ecológicas Rápidas, IAVH, Villa de Leyva. 76 p. [ Links ] [ Links ]
JIMÉNEZ, M. & NAVARRETE, J. G., 2010.- Fauna de arañas del suelo de una comunidad árida-tropical en Baja California. Sur, México. Revista Mexicana de Biodiversidad, 81: 417-426. [ Links ] [ Links ]
LEVI, H., 2002.- Keys to the genera of araneid orbweavers (Araneae, Araneidae) of the Americas. The Journal of Arachnology, 30: 527-562. [ Links ] [ Links ]
LEVINGS, S. C. & WINDSOR, D. M., 1985.- Litter arthropod populations in a tropical deciduous forest: relationships between years and arthropod groups. J. Anim. Ecol., 54: 61-69. [ Links ] [ Links ]
MANI, M. S., 1968.- Ecology and biogeography of high altitude insects. Dr. Junk N. V. Publishers, The Hague. [ Links ]
MURPHY, P. G. & LUGO, A. E., 1986.- Ecology of tropical dry forest. Annals Review of Ecology and Systematics, 17: 67-68. [ Links ] [ Links ]
PLATNICK, N., s. f.- The World Spider Catalog, Version 11.0. [En línea]. Disponible en: http://research.amnh.org. [ Links ] [ Links ]
OPELL, B. D., 1979.- Revision of genera and tropical American species of the spider family Uloboridae. Bull. Mus. Comp. Zool., 148 (10): 443-549. [ Links ] [ Links ]
RIECHERT, S. E., 1974.- Thoughts on the ecological significance of spiders. Bioscience, 24: 352-356. [ Links ] [ Links ]
RUZICKA, V., 1987.- Biodiagnostic evaluation of epigeic spider communities. Ekológia (CSSR), 6: 345-357. [ Links ] [ Links ]
SAMU, F. & SZINETAR, C., 2002.- On the nature of agrobiont spiders. J. Arahnol., 30: 389-402. [ Links ] [ Links ]
UETZ, G. W., 1976.- Gradient analysis of spider communities in stream-side forest. Oecologia, 22: 373-385. [ Links ] [ Links ]
