










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versão impressa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.16 no.2 Manizales jul./dez. 2012
CARACTERES MORFOLÓGICOS EN ESPORAS DE POLYPODIACEAE Y SU APLICACIÓN EN LA RECONSTRUCCIÓN FILOGENÉTICA*
MORPHOLOGICAL CHARACTERS IN POLYPODIACEAE SPORES AND THEIR APPLICATION IN PHYLOGENETIC RECONSTRUCTION
Luz Amparo Triana-Moreno1
* Fr: 12-IV-2011. FA: 12-II-2012.
1 universidad de Caldas, departamento de Ciencias Biológicas, Manizales, Colombia. luz.triana@ucaldas.edu.co
Resumen
Durante el desarrollo del análisis filogenético preliminar de Pecluma M.G. Price (Polypodiaceae), se estudiaron ocho géneros de la familia, y en el grupo externo se incluyó Oleandra Cav. (Oleandraceae). Se estudió la morfología de las esporas, por ser una fuente de información poco explorada filogenéticamente. Se observó con microscopio electrónico de barrido en alto vacío. Para las esporas se estudiaron los caracteres "esporas clorofílicas", "forma" y "ornamentación". La distribución de los caracteres de las esporas en el cladograma de consenso estricto demuestra que la ornamentación tiende a ser constante dentro de géneros, y permite reconocer diferencias intergenéricas, de modo que los caracteres estudiados se perfilan como un insumo útil para precisar los complejos de especies o géneros que presentan una situación taxonómica conflictiva dentro de la familia (e.g. grupo de Polypodium dulce).
Palabras clave: filogenia, helechos, ornamentación de la espora, Oleandra, Polypodiaceae.
Abstract
During the performance of a preliminary phyogenetic study of Pecluma M.G. Price (Polypodiaceae), eight genera of the family were studied, as well as Oleandra Cav. (Oleandraceae), which was an element of the outgroup. The morphology of the spores was studied, because it is a phylogeneticaly unexplored information source. It was observed using high vacuum scanning electron microscopy. Three spore characters were studied: "chlorophyllous spores", "shape" and "ornamentation". The distribution of the spore characters on the strict consensus cladogram shows that ornamentation tends to be constant within genera, and it allows the recognition of intergeneric differences. Thus, the studied characters could be a useful source to precise the genera or species complexes that present a conflictive taxonomic situation within the family (v.g. Polypodium dulce group).
Key words: phylogeny, ferns, spore ornamentation, Oleandra, Polypodiaceae.
INTRODUCCIÓN
La palinología es una herramienta de importancia destacada en el estudio de la flora vascular, tanto por ser una fuente de información para estudios taxonómicos, sistemáticos y ecológicos de la flora reciente, como por sus aportes al conocimiento de los paleoambientes. Los helechos, como elementos constituyentes de la flora desde hace ca. 345 millones de años, han dejado su huella palinológica en el ambiente de manera tan trascendental, que han sido un elemento clave para explicar la repoblación de la tierra después de la gran extinción ocurrida hace 65 millones de años (MORAN, 1994; 2004). Aunque la mayoría de los esfuerzos de la palinología se han centrado en la flora fanerogámica, los helechos también se han estudiado desde esta perspectiva. Para estas plantas se han generado contribuciones extensas y rigurosas (KREMP & KAWASAKI, 1972; TRYON & LUGARDON, 1991) que han contribuido a la delimitación taxonómica de los géneros y familias, al proporcionar nueva información micromorfológica.
La información que proveen las esporas ha sido aprovechada para la reconstrucción filogenética de helechos en diferentes categorías taxonómicas, como las familias Marsileaceae (SCHNEIDER & PRYER, 2002), Selaginellaceae (KORALL & TAYLOR, 2006) y Lindsaeaceae (LEHTONEN et al., 2010), y para géneros como Elaphoglossum (Dryopteridaceae) (MORAN et al., 2007) y Lomariopsis (Lomariopsidaceae) (ROUHAN et al., 2007), y varias Polypodiaceae grammitidoides como Lellingeria, Melpomene y Terpsichore (Polypodiaceae) (LABIAK et al., 2010a; SUNDUE, 2010; SUNDUE et al., 2010), Leucothrichum (LABIAK et al., 2010b) y Moranopteris (Polypodiaceae) (HIRAI et al., 2011).
Polypodiaceae (sensu SMITH et al., 2006) es una de las familias de helechos más diversas, tanto taxonómica como morfológicamente. Sus estudios palinológicos han abarcado intereses variados, como la descripción de la morfología de la espora en algunos géneros (GIUDICE et al., 2004; RAMOS-GIACOSA et al., 2007), o la identificación de patrones de desarrollo y evolución de la pared de la espora (VAN UFFELEN, 1997). De igual manera, la familia ha sido objeto de estudio para poner a prueba diferentes metodologías de observación palinológica, y determinar sus ventajas y alcances (VAN UFFELEN, 2000).
Desde el punto de vista sistemático, los grupos altamente diversos como Polypodiaceae, suelen presentar problemas de resolución en las relaciones de parentesco entre algunos de sus elementos. Esta situación induce a la exploración de fuentes de información poco utilizadas filogenéticamente. En este caso se propone estudiar la morfología de las esporas, porque éstas han sido escasamente aprovechadas como fuente de información filogenética en las Polypodiaceae no grammitidoides, a pesar de que se han identificado caracteres que definen claramente algunos géneros, y de los que se presume que podrían ser sus sinapomorfías (GIUDICE et al., 2004). Aún son incipientes los estudios filogenéticos que involucran caracteres de la espora para las Polypodiaceae no grammitidoides; en el neotrópico se conoce el análisis del complejo mesoamericano de Polypodium plesiosorum (TEJERO-DÍEZ, 2005).
En el marco del análisis filogenético preliminar de Pecluma M.G. Price (TRIANAMORENO, 2009), se propuso estudiar la micromorfología de las esporas de éste y otros géneros allegados, con el propósito de explorar su capacidad de resolución filogenética.
METODOLOGÍA
Se estudiaron esporas de los géneros Campyloneurum, Melpomene, Microgramma, Pecluma, Phlebodium, Pleopeltis, Polypodium y Terpsichore. Se tomó el género Oleandra (Oleandraceae) como el elemento más externo del outgroup, pues es el género neotropical más cercanamente relacionado con Polypodiaceae (SMITH et al., 2006).
Las esporas estudiadas fueron tomadas de ejemplares de herbario y se observaron con microscopía electrónica de barrido (MEB). Las esporas clorofílicas fueron tratadas previamente con una solución de CO3Na2 3% al calor por dos minutos, lavadas, deshidratadas y suspendidas en etanol 96% (GIUDICE et al., 2004; RAMOSGIACOSA et al., 2007); para su observación se depositaron directamente sobre los stubs. Las esporas preservadas en seco fueron montadas en cintas de carbono o en cintas doble faz, se metalizaron con oro-paladio, y se fotografiaron en alto vacío. Los caracteres y estados de carácter estudiados son: "esporas clorofílicas" (si / no), "forma de las esporas" (Fig. 1) (elipsoide / globosa / tetraédrica) y "ornamentación de la espora" (Fig. 1) (coliculada / tuberculada / verrugosa / retiado-perforada / papilada). Esta información fue codificada y analizada por TRIANA-MORENO (2009), junto con 46 caracteres más, en su mayoría vegetativos. La terminología palinológica corresponde a la propuesta por LELLINGER & TAYLOR (1997) y por LELLINGER (2002).
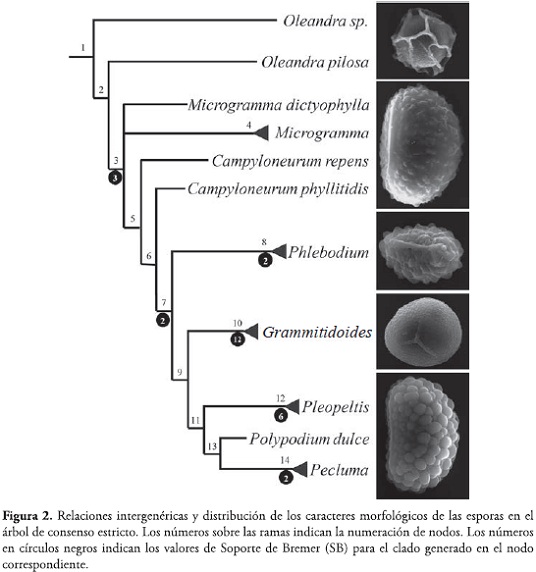
Para interpretar el comportamiento de los caracteres de las esporas se mapeó la distribución de dichos caracteres en el árbol de consenso estricto obtenido por TRIANA-MORENO (2009). Este análisis filogenético se logró por medio de una búsqueda heurística ejecutada por NONA ver. 2.0 (GOLOBOFF, 1993) a través de ASADO ver. 1,50 (NIXON, 2004). Para el cálculo de los índices de Soporte de Bremer (SB) se usó el programa Paup* ver. 4.0b10 para Macintosh (SWOFFORD, 2002).
RESULTADOS Y DISCUSIÓN
El análisis cladístico desarrollado por TRIANA-MORENO (2009) produjo dos árboles más parsimoniosos (L=181, IC=0.35, IR=0.69). Al mapear la distribución de los caracteres de las esporas en el cladograma de consenso estricto (Fig. 2) es posible observar que la ornamentación tiende a ser constante dentro géneros, pero permite establecer diferencias intergenéricas. La monofilia de Polypodiaceae aparece sustentada por la diferenciación en la ornamentación de la espora, la cual deja de presentar una perispora gruesa que forma grandes crestas, como ocurre en Oleandra (Fig. 1d), y en cambio empieza a manifestar novedades evolutivas que contribuyen a definir algunos géneros. Es así como en el nodo 3 aparece la ornamentación verrugosa, característica de Microgramma y Campyloneurum. La ornamentación tuberculada identifica a Phlebodium (nodo 8), mientras que la papilada aparece en los helechos grammitidoides, Melpomene y Terpsichore (nodo 10), que aparecen anidados en los polypodioides, confirmando la posición filogenética y la clasificación propuesta por SMITH et al. (2006).
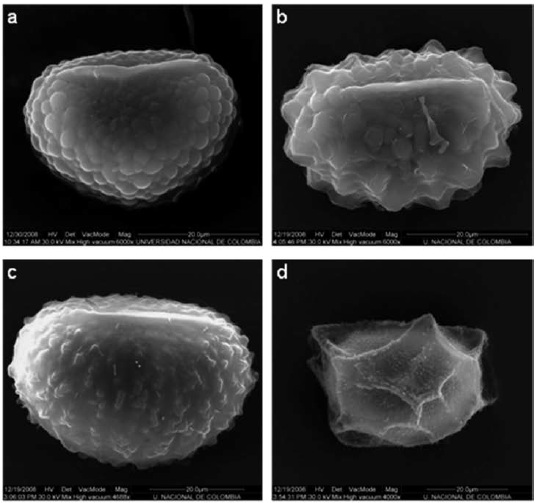
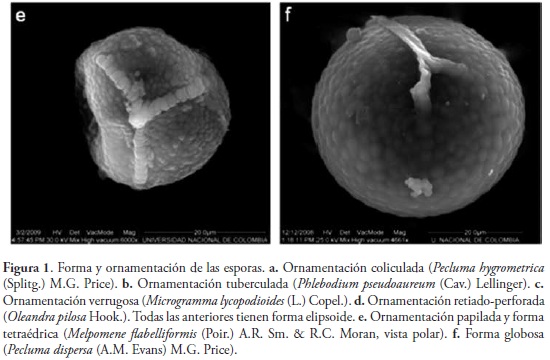
Más adelante, en el nodo 11, aparece la ornamentación coliculada, relacionado a los elementos de Pleopeltis, Polypodium y Pecluma; por presentar el mismo tipo de ornamentación, TRYON &LUGARDON (1991) opinaron que la evidencia palinológica era argumento suficiente para mantener a estos tres géneros dentro de Polypodium, en donde estuvieron circunscritos años atrás, a pesar de que presentaban diferencias macromorfológicas que sustentan su categoría de género. posteriormente, GIUDICE et al. (2004) advirtieron la escasa utilización de los caracteres de la espora para establecer relaciones filogéneticas de Polypodiaceae, pese a su significancia, pues muestran que, aunque los caracteres de la espora pueden resultar conservativos dentro d elos géneros, comtribuyen a diferenciar tanto complejos de especies como a los géneros entre sí, como lo verifican los resultados de este estudio.
Acerca de los demás caracteres, las esporas grammitidoides son clorofílicas y tetraédricas, mientras que las demás son amarillentas y elipsoides, excepto en Pecluma dispersa, donde son globosas (Fig. 1f), lo cual se asocia a su particular ciclo de vida apogámico ameiótico, en el cual se generan 32 diplosporas viables por esporangio, mientras que en otras especies del género, con un ciclo sexual normal, cada esporangio contiene 64 esporas (EVANS, 1968).
Aunque los caracteres observados contribuyeron a resolver relaciones intergenéricas, conviene verificar si la codificación de caracteres más finos, como algunas inclusiones esféricas o glomérulos intercalados con la ornamentación gruesa, podría aportar resolución a nivel intragenérico. De igual manera, es importante extender la observación de estos caracteres, en un contexto filogenético, hacia aquellos complejos de especies o géneros de la familia que se encuentran en una situación taxonómica imprecisa, como sucede, por ejemplo, con el "grupo de Polypodium dulce" (MORAN, 1995; TRIANA-MORENO, 2009), para el cual aún son escasos los argumentos que contribuyan a definir su situación, debido a que los estudios que lo incluyen no abarcan todo el complejo ni consideran detalles tan minuciosos como los estudiados aquí. Al respecto, TRIANA-MORENO (2009) considera que, si bien se verifica una relación cercana entre Pecluma y el "grupo de Polypodium dulce", la escasa representatividad de dicho grupo en el estudio no permite aseverar que sea monofilético, ni expresar con certeza su parentesco con Pecluma, su aparente grupo hermano.
AGRADECIMIENTOS
Esta investigación fue financiada por la Universidad Nacional de Colombia, a través de la Dirección de Investigación, sede Bogotá (proyecto DIB-8003162 y plan de acción COL0044434), y del departamento de Biología, la Facultad de Ciencias y la Dirección de Bienestar de la sede Bogotá. Agradezco a la Universidad Nacional de Colombia por la infraestructura puesta a disposición de esta investigación: el Herbario Nacional Colombiano (COL), el laboratorio de investigación en Fisiología y Bioquímica Vegetal del Departamento de Biología, y el laboratorio de Microscopía Electrónica de Barrido. A los herbarios COL y NY, que permitieron la toma de esporas de sus especímenes. A Jefferson Prado por su valiosa orientación. A Kevin Nixon por autorizarme para usar su programa ASADO, a Gabriela Giudice por su orientación metodológica para el estudio de las esporas. A Robbin Moran y George Yatskievych, quienes me proporcionaron valioso material bibliográfico. A Julián Mostacero por sus acertados aportes para mejorar este documento.
BIBLIOGRAFÍA
EVANS, A.M. 1968. - Interspecific relationships in the Polypodium pectinatum-plumula complex. Annals of the Missouri Botanical Garden 55: 193-293. [ Links ]
GIUDICE, G. E., M. A. MORBELLI, M. R. PIÑEIRO, M. COPELLO & G. ERRA. 2004. - Spore morphology of the Polypodiaceae from northwestern Argentina. American Fern Journal 94: 9-27. [ Links ]
GOLOBOFF, P. A. 1993. - NONA. Noname (a bastard son of Pee-Wee) ver. 2.0 (for Windows). Programa y documentación distribuidos por el autor. Fundación e Instituto Miguel Lilló, Tucumán. [ Links ]
HIRAI, R. Y., G. ROUHAN, P. H. LABIAK, T. A. RANKER & J. PRADO. 2011. - Moranopteris: A new Neotropical genus of grammitid ferns (Polypodiaceae) segregated from Asian Micropolypodium. Taxon 60(4): 1123-1137. [ Links ]
KREMP, G. O. W. & T. KAWASAKI. 1972. - The spores of the pteridophytes. Hirokawa Publishing Company, Tokio. [ Links ]
KORALL, P. & W. A. TAYLOR. 2006. - Megaspore morphology in the Selaginellaceae in a phylogenetic context: A study of the megaspore surface and wall structure using scanning electron microscopy. Grana 45: 22-60. [ Links ]
LABIAK, P. H., M. SUNDUE & G. ROUHAN. 2010a. - Molecular phylogeny, character evolution, and biogeography of the grammitid fern genus Lellingeria (Polypodiaceae). American Journal of Botany 97(8): 1354–1364. [ Links ]
LABIAK, P. H., G. ROUHAN & M. SUNDUE. 2010b. - Phylogeny and taxonomy of Leucotrichum (Polypodiaceae): A new genus of grammitid ferns from the Neotropics. Taxon 59(3): 911-921. [ Links ]
LEHTONEN, S., H. TUOMISTO, G. ROUHAN & M. J. M. CHRISTENHUSZ. 2010. - Phylogenetics and classification of the pantropical fern family Lindsaeaceae. Botanical Journal of the Linnean Society 163: 305-359. [ Links ]
LELLINGER, D. B. 2002. - A modern multilingual glossary for taxonomic pteridology. Pteridologia 3: 1-263. [ Links ]
LELLINGER, D. B. & W. C. TAYLOR. 1997. - Classification of spore ornamentation in the pteridophyta. pp. 33-42. En: JOHNS, R. J. (Ed.), Holtum: memorial volume. Royal Botanic Gardens. Kew. [ Links ]
MORAN, R. C. 1994. - The fern spike. Fiddlehead Forum 21(3): 32–33. [ Links ]
MORAN, R. C. 1995. - Polypodium. En: G. DAVIDSE, M. SOUZA & S. KNAPP (Eds.), Flora Mesoamericana 1: 349-365. Universidad Nacional Autónoma de México, Ciudad Universitaria, México D.F. [ Links ]
MORAN, R. C. 2004. - A Natural History of Ferns. Timber Press, Portland. [ Links ]
MORAN, R. C., J. G. HANKS & G. ROUHAN. 2007. - Spore morphology in relation to phylogeny in the fern genus Elaphoglossum (Dryopteridaceae). International Journal of Plant Sciences 168(6):905–929. [ Links ]
NIXON, K. 2004. - ASADO ver. 1.5. Programa y documentación distribuidos por el autor. Cornell University, Ithaca, Nueva York. [ Links ]
RAMOS-GIACOSA, J. P., M. A.MORBELLI & G. E. GIUDICE. 2007. - Morphology and ultrastructure of the spores of the Grammitidaceae from Argentina. Review of Paleobotany and Palynology 143: 155–166. [ Links ]
ROUHAN, G., J. G. HANKS, D. McCLELLAND & R. C. MORAN. 2007. - Preliminary phylogenetic analysis of the fern genus Lomariopsis (Lomariopsidaceae). Brittona 59(2): 115-128. [ Links ]
SCHNEIDER, H. & K. M. PRYER. 2002. - Structure and function of spores in the aquatic heterosporous fern family Marsileaceae. International Journal of Plant Sciences 163(4):485–505. [ Links ]
SMITH, A. R., K. M. PRYER, E. SCHUETTPELZ, P. KORALL, H. SCHNEIDER & P. G. WOLF. 2006. - A classification for extant ferns. Taxon 55: 705-731. [ Links ]
SUNDUE, M. A. 2010. - A morphological cladistic analysis of Terpsichore (Polypodiaceae). Systematic Botany 35(4): 716-729. [ Links ]
SUNDUE, M. A., M. B. ISLAM & T. A. RANKER. 2010. - Systematics of Grammitid ferns (Polypodiaceae): using morphology and plastid sequence data to resolve the circumscriptions of Melpomene and the polyphyletic genera Lellingeria and Terpsichore. Systematic Botany 35(4): 701-715. [ Links ]
SWOFFORD, D. L. 2002. - Paup* Phylogenetic Analysis Using Parsimony (*and Other Methods). Versión 4.0b10 para Macintosh. Sinauer, Sunderland. [ Links ]
TEJERO-DÍEZ, J. D. 2005. - Revisión taxonómica del complejo Polypodium plesiosorum Kunze (Polypodiaceae, Polypodiophyta). Tesis de Doctorado. Universidad Autónoma Metropolitana, Iztapalapa, México. [ Links ]
TRIANA-MORENO, L. A. 2009. - El género Pecluma (Polypodiaceae) en Colombia. Aproximación filogenética y revisión taxonómica. Tesis de Maestría. Universidad Nacional de Colombia, Bogotá [ Links ].
TRYON, A. F. & B. LUGARDON. 1991. - Spores of the pteridophyta: surface, wall structure, and diversity based on electron microscope studies. Springer Verlaag, Nueva York. [ Links ]
VAN UFFELEN, G. A. 1997. - The spore wall in Polypodiaceae: Development and evolution. Pp. 95-117. En: JOHNS, R. J. (Ed.) Holttum Memorial Volume. Royal Botanic Gardens, Kew. [ Links ]
VAN UFFELEN, G. A. 2000. - Studying spores of the Polypodiaceae: A comparison of SEM with other microscope techniques. pp. 125-131. En: HARLEY, M. M., C. M. MORTON & S. BLACKMORE (Eds.) Pollen and spores: Morphology and biology. Royal Botanic Gardens, Kew. [ Links ]
