










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.16 no.2 Manizales July/Dec. 2012
MORPHOMETRIC RELATIONSHIPS OF COMMERCIALLY IMPORTANT DEEP-SEA CRUSTACEANS IN THE COLOMBIAN CARIBBEAN SEA
* FR: 19-VIII-2010. FA: 20-V-2012.
1 Estudiante Programa de Ingeniería Pesquera, Universidad del Magdalena, Grupo de Investigación Ciencia y Tecnología Pesquera Tropical (CITEPT), Cra. 32 No. 22-08 Avenida del Ferrocarril, Santa Marta, Colombia. Email: ingp.eyneletrodriguez@gmail.com.
2 Estudiante Programa de Ingeniería Pesquera, Universidad del Magdalena, Grupo de Investigación Ciencia y Tecnología Pesquera Tropical (CITEPT), Cra. 32 No. 22-08 Avenida del Ferrocarril, Santa Marta, Colombia. Email: ing.gustavoardila@gmail.com
3 Profesor Asociado Programa de Ingeniería Pesquera, Universidad del Magdalena, Grupo de Investigación Ciencia y Tecnología Pesquera Tropical (CITEPT), Cra. 32 No. 22-08 Avenida del Ferrocarril, Santa Marta, Colombia. Email: jparamo@unimagdalena.edu.co
Con el propósito de suministrar información biológica de los crustáceos de profundidad de importancia comercial para una nueva pesquería en el Mar Caribe colombiano, en el presente estudio se describen las relaciones morfométricas de la gamba española (Aristaeomorpha foliacea), el camarón rojo real (Pleoticus robustus), el camarón rosado manchado (Penaeopsis serrata) y la langosta de aguas profundas del Caribe (Metanephrops binghami). Las muestras fueron colectadas en el Caribe colombiano con pesca de arrastre en profundidades entre 200 y 550 m (estratos de profundidad de 100 m). Se registró el peso total, sexo y número de individuos por especie en cada estación. La longitud total (LT) vs longitud cefalotórax (LC) y el peso total (PT) vs LT se determinó con una regresión lineal usando un modelo linealizado (logaritmo) alométrico. Se encontraron diferencias estadísticamente significativas en las tallas entre sexos para las especies de camarones de aguas profundas, revelando dimorfismo sexual. Aunque, no se hallaron diferencias estadísticamente significativas entre las tallas de M. binghami en los sexos, revelando homogeneidad en las tallas. Las hembras de A. foliacea y P. robustus incrementan el PT alométricamente 3,27 y 3,32 veces cada incremento en LT, respectivamente, pero los machos de estas dos especies tienen un crecimiento isométrico del PT con relación a la LT. La especie P. serrata mostró un crecimiento alométrico negativo en hembras y machos. Las hembras y machos de M. binghami mostraron un crecimiento alométrico positivo, es decir, ésta especie incrementa más en PT respecto a la LT.
Palabras clave: morfometría, Caribe colombiano, crustáceos de aguas profundas.
AbstractWith the purpose of providing biological information about commercially important deep-sea crustaceans for a new fishery in the Colombian Caribbean Sea, in this study the morphometric relationships of the giant red shrimp (Aristaeomorpha foliacea), the royal red shrimp (Pleoticus robustus), the pink speckled shrimp (Penaeopsis serrata) and the Caribbean lobster (Metanephrops binghami) were described in this study. The samples were collected in the Colombian Caribbean Sea using trawling nets in depths between 200 and 550 m (100 m strata intervals). The total weight, sex and number of individuals per species from each station were recorded. Total length (TL) vs cephalothorax length (CL) and total weight (TW) vs TL were determined with a linear regression using a linear allometric model (logarithm). Statistical significant differences were found in the size between sexes for deep-sea shrimp species, revealing sexual dimorphism. Even though, significant differences between the size of sexes of M. binghami were not found, revealing size homogeneity. The A. foliacea and P. robustus females increased the WT allometrically 3.27 and 3.32 times each increase in TL, respectively, but the males of these two species had an isometric growth of TW in relation to TL. The P. serrata species showed a negative allometric growth in the female and male. M. bighami females and males showed a positive allometric growth, that is to say, this species increased more in TW regarding TL.
Key words: morphometry, Colombian Caribbean Sea, deep-sea crustaceans.
En el Caribe colombiano la pesquería de camarón de aguas someras está dirigida principalmente a Farfantepenaeus notialis (PÉREZ-FARFANTE, 1967), mientras las especies de camarones F. brasiliensis (LATREILLE, 1817), F. subtilis (PÉREZ- FARFANTE, 1967) y Litopenaeus schmitii (BURKENROAD, 1936) aportan un porcentaje significativamente pequeño de las capturas. No obstante, éste es un caso típico de una pesquería colapsada debido a un manejo pesquero inadecuado (PARAMO & SAINT-PAUL, 2010). Por lo tanto, el futuro desarrollo del sector pesquero en el Caribe colombiano debe enfocarse en la búsqueda de nuevos recursos que sean económicamente atractivos, pero al mismo tiempo la extracción no debe poner en riesgo la sostenibilidad. Actualmente, no existe una pesquería de aguas profundas desarrollada en el Caribe colombiano. Si bien, estudios previos han identificado el potencial de nuevas pesquerías de crustáceos de profundidad con alto valor comercial en el Caribe colombiano, tales como, la gamba española -Aristaeomorpha foliacea- (RISSO, 1827), el camarón rojo real -Pleoticus robustus- (SMITH, 1885) (PARAMO & SAINT-PAUL, 2012a), el camarón rosado manchado -Penaeopsis serrata- (BATE, 1881) (PARAMO & SAINT-PAUL, 2012b) y la langosta de aguas profundas del Caribe -Metanephrops binghami- (BOONE 1927) (PARAMO & SAINT-PAUL, 2012c) (Tabla 1).

Desde el punto de vista pesquero es relevante conocer la estructura de las poblaciones para el éxito en el manejo pesquero y, más aun en pesquerías de crustáceos, ya que al analizar las tallas se pueden describir los parámetros demográficos de una población con relación a la presión pesquera (LIZÁRRAGA-CUBEDO, et al. 2008). Además, los patrones de variación morfométrica indican diferencias en el crecimiento, pues la forma del cuerpo es producto de la ontogenia, es decir, cambios estructurales en el desarrollo del organismo, lo cual es muy importante para implementar eficientes medidas de manejo pesquero (CADRIN, 2005). Mediante la determinación de las diferentes relaciones entre la estructura de tallas de cada una de las especies, se pueden implementar medidas de manejo para el inicio de una pesquería, tales como: talla media de captura, selectividad en las artes de pesca, tipo de crecimiento (alométrico e isométrico), etc. (BARBOSA-SALDAÑA, et al. 2012). Si bien, la talla es usualmente medida como una longitud, las mediciones de peso son requeridas en pesquerías para calcular el rendimiento pesquero, por lo cual es muy útil determinar las relaciones morfométricas (KING, 2007). Con el propósito de suministrar información biológica de crustáceos de profundidad con importancia comercial para una nueva pesquería en el mar Caribe colombiano; en el presente estudio se describen las relaciones morfométricas de la gamba española (A. foliacea), el camarón rojo real (P. robustus), el camarón rosado manchado (P. serrata) y la langosta de aguas profundas del Caribe (M. binghami).
MATERIALES Y MÉTODOSÁrea de estudio
Las muestras fueron colectadas en el mar Caribe colombiano con pesca de arrastre en profundidades entre 200 y 550 m (estratos de profundidad de 100 m). Se realizaron muestreos en agosto y diciembre de 2009; marzo y mayo de 2010 (Fig. 1), en el barco camaronero comercial "Tee Claude" usando una red de arrastre con tamaño de malla al final del copo de 44,5 mm de nudo a nudo, sobre una grilla de 87 estaciones, con un mínimo de dos lances por cada estrato de profundidad. No fue posible colectar muestras entre Cartagena y el río Magdalena debido a la irregularidad del fondo. La ubicación de los arrastres fue identificada usando un ecosonda comercial FURUNO FCV 1150 con un transductor de frecuencia de 28 kHz. La duración del arrastre fue de 30 minutos y la distancia arrastrada por la red y la Relaciones morfométricas de crustáceos de profundidad 239 velocidad del barco (2,5 nudos en promedio) fue estimada con un GPS Garmin MAP 76CSx. La abertura de la red fue 11,58 m.

ESTRUCTURA DE TALLAS
Se registró el peso total, sexo y número de individuos por especie en cada estación. Todos los especímenes fueron pesados (peso total, PT) con precisión de 0,1 g, la longitud total (LT) fue medida desde la punta del rostro hasta la punta del telson y, la longitud del cefalotórax (LC) desde el borde posterior de la cavidad orbital a la mitad del margen final del cefalotórax, las cuales fueron medidas con precisión de 0,01 mm usando calibradores digitales. Se evaluaron las diferencias en la distribución de frecuencias de las tallas entre hembras y machos usando la prueba no paramétrica Kruskal-Wallis (GOTELLI & ELLISON, 2004). La relación de la longitud total (LT) vs longitud cefalotórax (LC) se determinó con una regresión lineal, usando un modelo linealizado (logaritmo) alométrico con el fin de suavizar la varianza (ZAR, 2009):
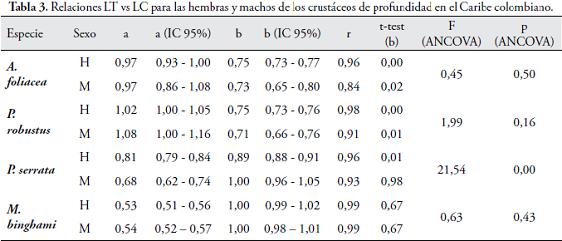
Donde LT es la variable dependiente, LC es la variable independiente, b es la pendiente, el cual muestra una variación de LT o tasa de crecimiento en función de LC, llamado también el coeficiente de alometría (FERREIRA, et al. 2008) (b > 3 alometría positiva, el individuo es más pesado que la longitud que tiene; b < 3 alometría negativa, el individuo es menos pesado que la longitud que tiene; b=3 isometría, el individuo crece a la misma tasa en todas las dimensiones) y "a" es el intercepto que corresponde al punto medio de LT cuando LC se hace cero, lo que permite estimar las longitudes promedio de la longitud total en una longitud de cefalotórax dada (KING, 2007), ∈ es el error. En el modelo PT = a*LTb se utilizó la relación linealizada (logaritmo) Log PT = Log a + b* Log LT + e. Se utilizó el coeficiente de correlación (r) como índice de la fuerza de asociación lineal. La prueba de t-student fue usada para comprobar la hipótesis de isometría (b = 1 o b = 3 para las relaciones LT vs LC y PT vs LT, respectivamente). Para evaluar las diferencias entre los sexos en las relaciones lineales se efectuó un análisis de covarianza (ANCOVA) a una vía, una vez los supuestos de homocedasticidad de las pendientes (paralelismo) fueron encontrados con los datos transformados en logaritmo (ZAR, 2009).
RESULTADOSSe encontraron diferencias estadísticamente significativas en las tallas entre sexos para las especies de camarones de aguas profundas (Fig. 2; Tabla 2), revelando dimorfismo sexual (p = 0,00).
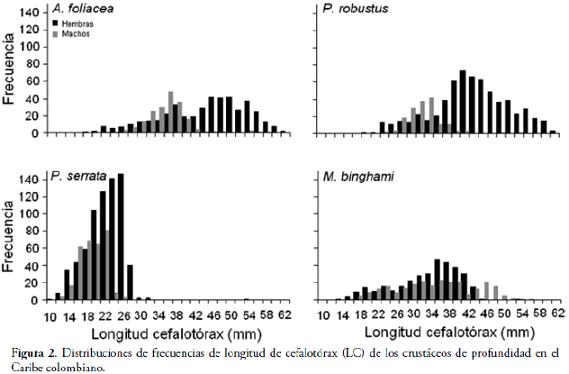

Los machos fueron más pequeños que las hembras en todas las especies de camarones de profundidad. La estructura de tallas para las hembras de A. foliacea varía entre 18,12 y 63,09 mm de LC (talla media 44,22 ± 9,41 mm) y para los machos entre 26,57 y 43,45 mm en LC (talla media 35,26 ± 3,08 mm). La distribución de frecuencia de LC para las hembras de P. robustus varió entre 21,20 y 62,84 mm LC (talla media 43,68 ± 8,37 mm LC) y en machos entre 24,30 y 43,09 mm (talla media 32,43 ± 3,41 mm LC). La estructura de tallas para P. serrata en las hembras estuvo entre 11,53 y 32,70, con una media de 22,10 ±3,83 mm LC, mientras que las tallas de los machos osciló entre 12,21 y 26,16, con una media de 18,94 ±2,57 mm LC. En la langosta del Caribe (M. binghami) la talla de las hembras varió entre 15,5 y 55,3 mm LC (talla media 33,6 ±7,2 mm) y para los machos entre 17,0 y 56,3 mm LC (talla media 34,8 ±8,9 mm). Sin embargo, entre los crustáceos de profundidad estudiados, sólo en la langosta no se encontraron diferencias estadísticamente significativas entre las tallas (M. binghami) (Fig. 2; Tabla 2), revelando homogeneidad en las tallas para ambos sexos (p = 0,09). Se observaron coeficientes de correlación altos en las relaciones lineales entre LT vs LC (Fig. 3; Tabla 3).

Los camarones de profundidad mostraron significativo crecimiento alométrico negativo (b < 1) (p < 0,05), excepto para los machos de P. serrata y, las hembras como machos de la langosta de profundidad M. binghami (p > 0,05) (Fig. 3; Tabla 3). No se encontraron diferencias significativas (ANCOVA: p > 0,05) entre las pendientes de las hembras y machos en las especies A. foliacea, P. robustus y M. binghami, pero si se encontraron diferencias significativas (ANCOVA: p < 0,05) en las pendientes de las hembras y machos de P. serrata (Tabla 3).
Las relaciones lineales entre PT vs LT mostraron coeficientes de correlación altos (Fig. 4; Tabla 4).
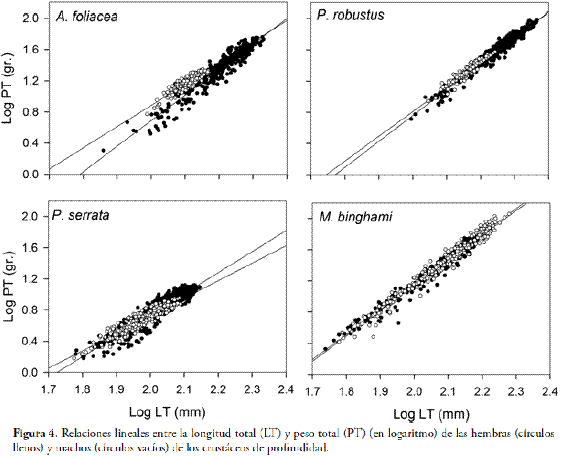
El crecimiento relativo mostró alometría significante positiva (b > 3) (p < 0,05) en las hembras de A. foliacea y P. robustus como en las hembras y machos de M. binghami, pero también alometría significativamente negativa (b < 3) (p < 0,05) en ambos sexos de P. serrata (Fig. 4; Tabla 4). Por el contrario, los machos de A. foliacea y P. robustus mostraron isometría (b=3) (p > 0,05). Se encontraron diferencias significativas (ANCOVA: p < 0,05) entre las pendientes de las hembras y machos en A. foliacea y P. serrata, pero no se encontraron diferencias significativas (ANCOVA: p > 0,05) entre las pendientes por sexo en P. robustus y M. binghami.

DISCUSIÓN
Las pesquerías más importantes de crustáceos son los decápodos (JENNINGS, et al. 2006), de los cuales los camarones (Penaeoidea) y las langostas (Nephropoidea) son los más abundantes de los ecosistemas de aguas profundas tropicales (KING, 2007). La gamba española (A. foliacea) se encuentra en las aguas profundas del Mar Mediterráneo y es un camarón comercialmente importante (D'ONGHIA, et al. 1998; FIGUEIREDO, et al. 2001; PAPACONSTANTINOU & KAPIRIS, 2003; POLITOU, et al. 2004; MOUFFOK, et al. 2008). GRACIA, et al. (2010) recientemente exploraron las aguas profundas de la Península de Yucatán en México y mostraron que A. foliácea y P. robustus representan un recurso pesquero potencial. Actualmente, A. foliacea constituye una pesquería valiosa de camarón de profundidad en la costa sur y sureste de Brasil (PEZZUTO, et al. 2006; DALLAGNOLO, et al. 2009). El camarón rojo real (P. robustus) es capturado en abundantes proporciones frente a la costa del noreste de Florida, cerca de las Islas Dry Tortugas, en el delta del río Mississippi y, también frente de la costa de Venezuela (TAVARES, 2002). El camarón rosado manchado (P. serrata) es de importancia comercial en el Atlántico este y oeste (HOLTHUIS, 1980). M. binghami tiene potencial para una explotación económica en las aguas de Venezuela (GÓMEZ, et al. 2000, 2005). En el Pacífico colombiano la pesquería de crustáceos de aguas profundas ha jugado un papel fundamental para el sustento de las comunidades de pescadores (DIAZ, 2011). Si bien, la pesca ha presentado una disminución en las capturas, se han implementado medidas de manejo como vedas en algunas épocas del año, para el uso sostenible del recurso (RUEDA, et al. 2010). En el Caribe colombiano, el número de hembras fue mayor que los machos en las tres especies de camarón de profundidad (A. foliacea, P. robustus y P. serrata), demostrando dimorfismo sexual. Tal característica también se encontró para A. foliacea en el Mar Mediterráneo indicando segregación entre sexos (D'ONGHIA, et al. 1998; PAPACONSTANTINOU & KAPIRIS, 2003). Sin embargo, la langosta de profundidad tiende a presentar homogeneidad en las tallas entre hembras y machos (MAYNOU, et al. 1998; GÓMEZ, et al. 2000; FONSECA, et al. 2007). La talla media de A. foliacea (hembras 44,2 mm; machos 35,8 mm LC) es similar a la misma especie encontrada en una región de pesca no explotada en el Mar Iónico de Grecia en profundidades entre 250 y 800 m (PAPACONSTANTINOU & KAPIRIS, 2003) y, mayores que la talla de máximo potencial reproductivo en hembras (37,11 mm CL) (D'ONGHIA, et al. 1998). Las tallas de M. binghami en este estudio son similares a Nephrops norvegicus, la cual es capturada en aguas de Portugal, variando entre 18 a 59 mm de LC (FONSECA, et al. 2007) y en el Mar Catalán (España, noroeste del Mediterráneo) variando entre 12 a 58 mm de LC (MAYNOU, et al. 1998), pero de menor tamaño del reportado para N. andamicus en Sur África variando entre 47 a 55 mm de LC (BERRY, 1969). Las hembras de A. foliacea y P. robustus en el Caribe colombiano incrementan el PT alométricamente 3,27 y 3,32 veces cada incremento en LT, respectivamente, aunque los machos de estas dos especies tienen un crecimiento isométrico del PT con relación a la LT. Dicho patrón de las hembras con mayor proporción en peso respecto a los machos para la misma longitud también se encontró para A. foliacea en el Mar Iónico en Grecia (Mediterráneo) (KAPIRIS, 2005). Sin embargo, en el estrecho de Sicilia (Mar Mediterráneo) la relación peso-talla mostró alometría negativa en las hembras y machos (RAGONESE, et al. 1997), lo cual coincide con el crecimiento alométrico negativo en las hembras y machos de P. serrata en el Caribe colombiano. Las hembras y machos de M. binghami muestran un crecimiento alométrico positivo, es decir, M. binghami incrementa más en PT respecto a la LT. Asimismo, se encontró para esta especie en Venezuela un crecimiento alométrico positivo y sin diferencias significativas entre sexos (GÓMEZ, et al. 2000).
De acuerdo a estos resultados, se necesita más investigación biológica para entender los parámetros del ciclo de vida de los crustáceos de aguas profundas en el Mar Caribe colombiano, tales como crecimiento, reproducción, reclutamiento, mortalidad, áreas y épocas de desove, áreas de crianza, además de la biodiversidad asociada antes de iniciar una nueva pesquería comercial. Dicha información ayudará a desarrollar estrategias apropiadas para manejar y sustentar las pesquerías (BIANCHI, 2008). El gran valor comercial de estas especies de crustáceos de aguas profundas, no sólo a nivel local sino también en mercados internacionales (BELCARI, et al. 2003; PAPACONSTGANTINO & KAPIRIS, 2003), hacen de estos crustáceos de aguas profundas un nuevo recurso económico potencial en el Mar Caribe colombiano
AGRADECIMIENTOSEste trabajo es una contribución del grupo de investigación Ciencia y Tecnología Pesquera Tropical (CITEPT) de la Universidad del Magdalena (Colombia). Agradecemos a la tripulación del barco "Tee Claude" y al Capitán José Guillem. El trabajo fue patrocinado por COLCIENCIAS (código 117-452-21288), la Universidad del Magdalena, el Instituto Colombiano de Desarrollo Rural (INCODER) a través de la Subgerencia de Pesca y Acuicultura, la Autoridad Nacional de Acuicultura y Pesca (AUNAP) convenio No. 790 y el Leibniz-Zentrum für Marine Tropenökologie (ZMT), Alemania.
BARBOSA-SALDAÑA, M.; DÍAZ-JAIMES, P. & URIBE-ALCODER. 2012.- Variacion morfológica del camarón café (Farfantepenaeus californiensis) en el Pacífico mexicano. Revista Mexicana de Biodiversidad, 83:45-50. [ Links ] [ Links ]
BERRY, P. F. 1969.- The biology of Nephrops andamicus Wood-Mason (Decapoda, Reptantia). Investigational Report No. 22. Oceanographic Research Institute, Durban. [ Links ] [ Links ]
CADRIN, S.X. 2005.- Morphometric landmarks: 153-172 (en) CADRIN, S.X. FRIEDLAND, K.D. & WALDMAN, J.R. (eds.) Stock identification methods: Applications inf fishery science. Academic Press. Londres. [ Links ] [ Links ]
DIAZ, J.; VIEIRA, C.; MELO, G. 2011.- Diagnostico de las principales pesquerías del Pacífico colombiano. Fundación Mar Viva: 25-30. [ Links ] [ Links ]
FERREIRA, S.; SOUSA, R.; DELGADO, J.; CARVALHO, D. & CHADA, T. 2008. - Weight-length relationships for demersal fish species caught off the Madeira archipelago (eastern-central atlantic). J. Appl. Ichthyol. 24: 93-95. [ Links ] [ Links ]
GÓMEZ, G.; GUZMÁN, R. & MARCANO, L. 2000.- Contribución al conocimiento de la Cigala del Caribe, Metanephrops binghami, en aguas de Venezuela: aspectos biométricos y fecundidad. Zoo. Trop. 18 (1): 107-118. [ Links ] [ Links ]
GOTELLI, N.J. & ELLISON, A.M. 2004.- A Primer of Ecological statistics. Sinauer Associates. [ Links ] [ Links ]
HOLTHUIS, L.B.; 1980.- Shrimps and prawns of the world. An annotated catalogue of species of interest to fisheries. (FAO Species Catalogue, 1). FAO Fisheries Synopsis. 125 (1): 1-271. [ Links ]
JENNINGS, S.; KAISER, M.J. & REYNOLDS, J.D. 2006.- Marine fisheries ecology. Blackwell Publishing. [ Links ] [ Links ]
KING, M. 2007.- Fisheries Biology, Assessment and Management. Wiley-Blackwell. [ Links ] [ Links ]
MAYNOU, F.X.; SARDÁ, F. & CONAN, G.Y. 1998.- Assessment of the spatial structure and biomass of Nephrops norvegicus (L) populations in the northwestern Mediterranean by geostatistics. ICES J. Mar. Sci., 55:102-120. [ Links ] [ Links ]
PAPACONSTANTINOU, C. & KAPIRIS, K. 2003.- The biology of the giant red shrimp (Aristaeomorpha foliacea) at an unexploited fishing ground in the Greek Ionian Sea. Fish. Res., 62: 37-51. [ Links ] [ Links ]
PARAMO, J. & SAINT-PAUL, U. 2010.- Morphological differentiation of southern pink shrimp Farfantepenaeus notialis in Colombian Caribbean Sea. Aquat. Liv. Res. 23(1): 95-101. [ Links ] [ Links ]
PARAMO, J. & SAINT-PAUL, U. 2012a.- Deep-sea shrimps Aristaeomorpha foliacea and Pleoticus robustus (Crustacea: Penaeoidea) in the Colombian Caribbean Sea as a new potential fishing resource. J. Mar. Biol. Ass. U.K. 92(4): 811-818. [ Links ] [ Links ]
PARAMO, J. & SAINT-PAUL, U. 2012c.- Spatial structure of deep sea lobster (Metanephrops binghami) in the Colombian Caribbean Sea. Helg. Mar. Res. 66: 25-31. [ Links ] [ Links ]
POLITOU, CH-Y.; KAPIRIS, K.; MAIORANO, P.; CAPEZZUTO, F. & DOKOS, J. 2004.- Deep-sea Mediterranean biology: the case of Aristaeomorpha foliacea (Risso, 1827) (Crustacea: Decapoda: Aristidae). Sci. Mar. 68(3): 129-139. [ Links ] [ Links ]
RUEDA, M.; RICO, F.; ANGULO, W.; GIRON, A.; RODRÍGUEZ, A.; GARCIA, L.; ARENAS, L. 2010.- Evaluación biológico-pesquera del estado de las poblaciones de camarón de aguas profundas, mediante la aplicación de métodos directos (prospección pesquera) e indirectos (estadísticas de captura y esfuerzo) en el Pacífico colombiano. Informe final de Proyecto código 031 2007T6650-909-07. MADR. INVEMAR. Santa Marta, Colombia. [ Links ] [ Links ]
ZAR, J.H. 2009.- Biostatistical Analysis. Prentice Hall. [ Links ]
