










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.17 no.1 Manizales Jan./June 2013
MARIPOSAS HESPERIOIDEA Y PAPILIONOIDEA (INSECTA: LEPIDOPTERA) EN UN FRAGMENTO DE BOSQUE SECO TROPICAL, ATLÁNTICO, COLOMBIA*
BUTTERFLIES PAPILIONOIDEA AND HESPERIOIDEA (INSECTA: LEPIDOPTERA) IN THE RESERVA ECOLÓGICA LURIZA (REL), ATLANTICO, COLOMBIA
Carlos Boom–Urueta1, Leidys Seña-Ramos2, María Vargas-Zapata2*, & Neis Martínez–Hernández3
* FR: 14-XII-2012. FA: 14-IV-2013.
1 Estudiante de Maestría en Biología. Universidad del Atlántico. Semillero Investigación Insectos (NEOPTERA) del Caribe colombiano. Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Ciudadela universitaria, km 7 vía Puerto Colombia, Barranquilla, Colombia. Cjboomu@gmail.com
2 Semillero Investigación Insectos (NEOPTERA) del Caribe colombiano. Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Ciudadela universitaria, km 7 vía Puerto Colombia, Barranquilla, Colombia. senaramosl@gmail.com,mariavargaszapata@gmail.com
3 Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Ciudadela universitaria, km 7- vía Puerto Colombia. Barranquilla, Colombia. neyjosemartinez@gmail.com
* Autor para correspondencia
Resumen
Las distintas actividades antrópicas en los últimos años ha aumentado la amenaza sobre el Bosque Seco Tropical (Bs-T) en el Caribe colombiano, disminuyendo considerablemente la fauna de insectos como los lepidópteros. Este grupo es usado ampliamente como bioindicador del estado de conservación de bosques, por su sensibilidad a los cambios ambientales y la relación estrecha con los recursos que ofrecen el bosque. Con el fin de aportar a esta discusión, se evaluó la variación de la riqueza y abundancia de las mariposas Papilionoidea y Hesperioidea en un fragmento de Bs-T en la Reserva Ecológica de Luriza (REL), Atlántico, Colombia. Se realizaron nueve eventos de muestreos desde marzo a julio de 2009; recolectándose los individuos con redes entomológicas y trampas tipo van Someren-Rydon cebadas con una mezcla de fruta fermentada, calamar en descomposición, orina humana y cerveza. Se capturaron 614 adultos que corresponden a 48 especies y 39 géneros. La familia Nymphalidae presentó la mayor riqueza (23) y abundancia (500), destacándose la especie Mechanitis lysimnia (Fabricius, 1793) con 307 ejemplares. Los valores más altos de riqueza y abundancia se presentaron durante el segundo muestreo con 21 especies y 106 individuos, coincidiendo con el período de ausencia de lluvias en la zona. Con respecto a la diversidad, los valores más alto de los números de Hill se presentaron en el último muestreo (N1= 9,76) y para N2= 6,642 en el octavo muestreo. El análisis de agrupamiento de Bray–Curtis demostró que los tres primeros muestreos presentaron una similitud mayor del 65%, los cuales coinciden con las ausencias de lluvias en el área; demostrándose que la composición y abundancia de Papilionoidea y Hesperioidea presenta un patrón temporal en la zona. Con las curvas de acumulación de especies se espera la existencia de más especies de lepidópteros en el bosque, por lo que se hace recomendable la realización de más muestreo en el área de estudio.
Palabras clave: Bosque Seco Tropical, lepidópteros, Bioindicador, Reserva Ecológica Luriza.
Abstract
The various human activities in recent years have caused a threat to the tropical dry forest ecosystems (Bs-T) in the Colombian Caribbean, significantly reducing the fauna of insects such as lepidoptera. This group is widely used as bioindicators of forest condition, for their sensitivity to environmental changes and relationship with the resources offered by the forest. In this study the variation of richness and abundance of Papilionoidea butterflies and Hesperioidea in the Reserva Ecologica Luriza (REL) were evaluated. Nine samples events were made from March to July 2009; the butterflies were captured with entomologic nets and van Someren-Rydon type traps with a mixture of fermented fruit, rotting squid, human urine and beer. 614 adults were registered corresponding to 48 species and 39 genera. The Nymphalidae family had the highest richness (23) and abundance (500), represented mainly by Mechanitis lysimnia (Fabricius, 1793) with 307 individuals. The highest values of richness and abundance occurred during the second sampling with 21 species and 106 individuals, coinciding with the period of no rain in the area. With respect to diversity, higher values of the Hill numbers were presented at the last sampling (N1 = 9.76) and N2 = 6.642 in the eighth sampling event. The Bray-Curtis analysis shows that the first three samples showed a greater than 65% similarity, which coincide with the absence of rain in the forest. With species accumulation curves the existence of more species of Lepidoptera in the forest is expected, so that it is advisable to carry out more samples in the studied area.
Key words: Tropical dry forest, Lepidoptera, bioindicator, Reserva Ecológica Luriza.
INTRODUCCIÓN
En el Caribe colombiano, encontramos los bosques secos más extensos que cubren unos 1.355 km2, pero casi la mitad de estos se encuentran intervenidos (DÍAZ, 2006; RUIZ & FANIÑO, 2009). El avance de los procesos antropogénicos en los últimos años ha originado una amenaza sobre este ecosistema en esta región del país, donde solo queda el 3% del bosque natural (MILES et al., 2006; PENNINGTON et al., 2006). Esta situación es más evidente en uno de los departamentos de menor extensión y con menor número de áreas protegidas naturales como es el caso del Departamento del Atlántico. En esta zona, la destrucción del Bs-T avanza de manera acelerada, debido a la explotación de madera, la expansión de sitios de forrajeos para la ganadería y la siembra de cultivos; lo que ha disminuido considerablemente este ecosistema, dando origen a remanentes con cierto grado de aislamiento (MARTÍNEZ et al., 2010). Sin embargo, aún existen algunos remanentes de bosque que se destacan por presentar una riqueza faunística importante; la cual interactúa con otros ecosistemas cercanos por migraciones locales que se dan por la disponibilidad de recursos, de microhábitats y la conectividad entre fragmentos (IAVH, 1998).
Debido a la fuerte intervención del Bs-T en la región Caribe colombiana (DÍAZ, 2006; RUIZ & FANIÑO, 2009) y la constante amenaza a este tipo de ecosistema, se hace necesario la identificación de grupos faunísticos que sean indicadores útiles en el monitoreo ambiental (BROWN, 1991). En este caso, las mariposas diurnas en los últimos años se han convertido en un importante grupo para el estudio y monitoreo de la biodiversidad para su conservación (LLORENTE et al., 1993; ANDRADE, 2002; VARGAS et al., 2011); debido a que se caracterizan por ser diversas y abundantes, presentar ciclos de vida cortos, estrecha afinidad con los principales grupos de plantas y alta sensibilidad a la deforestación (BROWN, 1991; KREMEN, 1992; POLLARD & YATES, 1994; VARGAS et al., 2011). Además, las mariposas responden rápidamente a las alteraciones ambientales, por lo tanto, la presencia de ciertos grupos puede indicar la continuidad de comunidades y su ausencia una fragmentación o alteración de la integridad del paisaje (ANDRADE, 1998, BONEBRAKE et al, 2000, SHAHABUDDIN & PONTE, 2005).
Se considera que en Colombia existen aproximadamente 3.500 especies de mariposas diurnas descritas por LE CROM et al. (2004). Sin embargo, la información sobre lepidopterofauna para algunos ecosistemas como el Bs-T en las zonas bajas de Caribe colombiano está muy incipiente; ya que son pocos los estudios taxonómicos, ecológicos y de distribución de especies. En la actualidad, se destacan los realizados por ERAZO & GONZÁLEZ (2008) en el Santuario de vida de los Besote y el de CAMPO & ANDRADE (2009) en el bosque el Aguil (Aguachica) en el departamento del Cesar y en el Magdalena, el realizado por VARGAS et al. (2011) en la Reserva Campesina Las Delicias.
En el caso de los fragmentos de Bs-T en el Departamento del Atlántico, vale la pena mencionar el listado taxonómico de Papilionoidea y Hesperioidea realizado por MONTERO et al. (2009) en cuatro localidades, en el que reporta 123 especies; mientras que PRINCE et al. (2011) reporta 74 especies en dos fragmentos de Bs-T en la localidad de Corrales de San Luis (Tubara). Pese al bajo número de trabajos realizados y teniendo en cuenta que los fragmentos de Bs-T han recibido poca atención por parte de los científicos y escasa gestión para su conservación (PRANCE, 2006), con el acto administrativo 0003 de marzo de 2011 emitido por la CRA, se declara como única área protegida a Luriza; una de las pocas áreas de bosque seco en este departamento. En esta reserva, los fragmentos de Bs-T ameritan estudios sobre el comportamiento de la fauna asociada de invertebrados y su dinámica ecológica; ya que en la actualidad solo se cuenta con inventarios de la fauna de vertebrados y de especies vegetales.
Con el fin de aportar a esta discusión, se determinó la variación de la riqueza y abundancia de mariposas Papilionoidea y Hesperioidea en la Reserva Ecológica de Luriza (REL), Atlántico. Lo anterior sería una contribución a la estimación de la riqueza biológica en este fragmento de Bs-T que servirá para reflejar su estado de conservación y que permita aportar la información necesaria para la elaboración de planes de manejo de conservación del área.
MATERIALES Y MÉTODOS
Este trabajo se llevó a cabo en la Reserva Ecológica de Luriza (REL), la cual se encuentra en el municipio de Usiacurí, Atlántico, Colombia, localizada a 10°45'27.8''N y 75°01'59.0''O (Figura 1) y tiene una extensión de 837.17ha. La reserva pertenece al distrito Montes de María y Piojó, ubicado dentro de la región biogeográfica del Cinturón Árido Pericaribeño (HERNÁNDEZ et al., 1992). Dentro de la reserva, se escogió un fragmento de bosque seco, con una extensión de 41,3ha y una altitud de 140m. El bosque se caracteriza por presentar condiciones higrotropofíticas y está atravesada por una quebrada que mantiene un curso de agua durante todo el año (MARTÍNEZ et al., 2010). El relieve es ondulado; debido a que se encuentra en las estribaciones de la serranía de Piojó. La precipitación anual oscila entre 600 y 700mm, con temperatura promedio anual de 26 a 28ºC, con una humedad relativa entre 60 y el 90% (MARTÍNEZ et al., 2010).
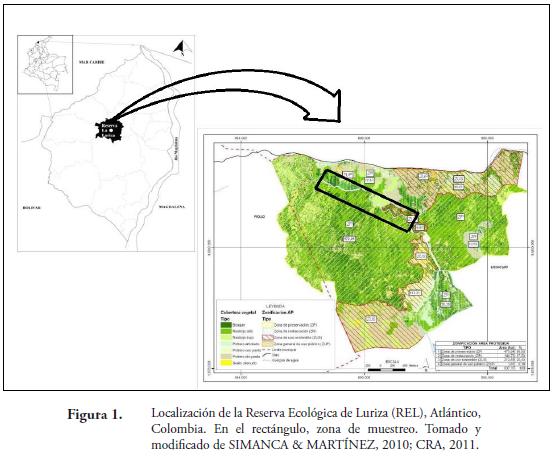
La vegetación dominante en la reserva es típica de Bs- T (IAVH, 1998), con árboles de hasta 17 m de altura. Las familias más comunes son las Apocynaceae, Arecaceae, Bignoniaceae, Euphorbiaceae, Fabaceae, Polygonaceae, entre otras (MARTÍNEZ et al., 2010). Entre los géneros más representativos de la vegetación se encuentran Ceiba, Pseudobombax, Tabebuia, Philodendron, Anthurium, Acrocomia, Aristolochia y las especies Spondias mombin L., Crescentia cujete L., Pseudobombax septenatum (Jacq.) Dugand, Anacardium excelsum L., Parinari pachyphylla Rusby, Justia bracteosa (Mildbr.) Leonard, Malvaviscus aboreus Dill. ex Cav, Myrmecodendron costarriscense Britt. & Rose, Petiveria alliiaceae L., Cordia alba (Jacq.) Roem. & Schult, Mangifera indica L., Guazuma ulmifolia Lam., Pereskia quisqueyana (Ekman) Alain, Bursera simaruba (L.) Sarg., Hura crepitans L., Pachira Aubl. y la presencia de algunas especies de musgos y helechos. El bosque se encuentra altamente amenazado, debido al avance de las áreas ganaderas y agrícolas y sitios donde se lleva a cabo tala y quema de árboles para la producción de carbón; situación que ha originado su reducción en los últimos años.
Diseño y métodos de captura
Se realizaron nueve eventos de muestreos cada 15 días entre los meses de marzo hasta julio de 2009. En el interior del fragmento se demarcó un transecto lineal de 500m de largo por 50m ancho, en el que se capturaron los especímenes a través de redes entomológicas desde las 08:00 hasta las 16:00 horas; con un esfuerzo de 9 horas/persona durante dos días. Adicionalmente, en el transecto se colocaron 15 trampas Van Someren–Rydon (VSR) en zig-zag, distanciadas 50m entre cada una de ellas. Las trampas fueron instaladas a una altura que osciló entre tres y 10m, las cuales fueron cebadas con una mezcla de fruta fermentada (papaya y plátano), calamar en descomposición, orina humana y cerveza.
Las especies capturadas se sacrificaron a través de la técnica de sujeción torácica y guardadas en sobres de papel Milano con códigos de registro y almacenadas en recipientes con naftalina (CAMERO & CALDERÓN, 2007), para su posterior identificación en el laboratorio. Los ejemplares de mariposas de fácil identificación fueron marcados con un marcador indeleble en la celda discal en la ala anterior con una letra diferente para cada muestreo y un número consecutivo (PRIETO et al., 2005) y posteriormente fueron liberados. Esta técnica se realizo con el fin de no tener sesgo de no contar nuevamente los individuos ya capturados y registrados. Los ejemplares recolectados se identificaron a través de las claves taxonómicas de Le CROM et al. (2002, 2004), CHACÓN & MONTERO (2007) y la pagina ilustrada de Butterflies of America (WARREN et al., 2012). Por otro lado, en cada muestreo y punto se midieron in situ la temperatura (°C) y la humedad relativa (%) con un termohigrómetro marca Max-Min Clock; mientras que los datos de precipitación fueron suministrados por el Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM), los cuales fueron ponderados para cada muestreo.
Análisis de datos
La riqueza de especies de Papilionoidea y Hesperioidea se consideró como el número de especies encontradas en cada muestreo y la abundancia como el número de individuos recolectados. Con el fin de estimar la eficiencia de los muestreos y aproximar el número de especies reales de la población en el fragmento de bosque, los resultados se analizaron con estimadores no paramétricos (ICE, Chao 2 y Bootstrap), que usan algoritmos para predecir la diversidad esperada de un lugar cuando se tiene datos de presencia-ausencia y bajo número de muestreos, usando el programa EstimateS 8.2 (COLWELL, 2009). También se estimó la diversidad con la serie de los Números de Hill (N1 y N2) descritos por MORENO (2001) y VILLAREAL et al. (2004); lo cual permite calcular el número efectivo de especies en una muestra y son altamente utilizados, ya que permiten reflejar tanto especies dominantes (N2) como especies comunes en una muestra (N1). De igual manera, se determinó la equitatividad mediante el Índice de Pielou (MORENO, 2001), el cual expresa la equidad como la proporción de la diversidad observada en relación con la máxima diversidad esperada. Estas estimaciones se llevaron a cabo a través del programa PRIMER 6.0 (CLARKE & WARWICK, 2001). Además, se determinó el índice de constancia de especies propuesto por BODENHEIMER (1955) y SILVEIRA et al. (1976), donde las especies fueron clasificadas como i) constantes (C): especies encontradas en más del 50% de ocasiones de muestreo; ii) accesorias (A): especies presentes entre 25% a 50%; iii) accidentales (Ac) especies presentes en menos del 25 % (SACKIS & MORAIS, 2008). También se estableció la similitud entre muestreos teniendo en cuenta los datos de abundancia de las especies por muestreo. A partir de estos resultados, se obtuvo un dendrograma elaborado por un análisis de agrupamiento por pares mediante el método de promedios no ponderados (UPGMA). El agrupamiento se hizo por medio de índice de similitud de Bray-Curtis. Para el análisis y construcción del agrupamiento fue utilizado el programa PRIMER 6.0. Previamente, los datos de abundancia fueron transformados a Ln(X + 1), con el fin de contrarrestar el peso de las especies más dominantes pero sin disminuir su importancia (CLARKE & WARWICK, 2001). Con el propósito de relacionar las variables ambientales (precipitación, temperatura y humedad relativa) con la riqueza y abundancia de lepidópteros Hesperioidea y Papilionoidea, se aplicó el índice de Correlación de Spearman; con la ayuda del programa PAST 1.63 (HAMMER et al., 2001)
RESULTADOS Y DISCUSIÓN
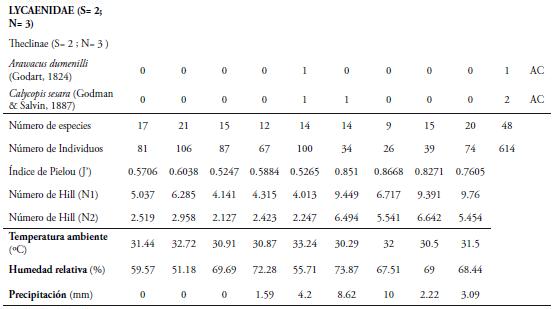
Se registraron 614 individuos de las Superfamilias Papilionoidea y Hesperioidea, distribuidos en 48 especies, 39 géneros, 14 subfamilias y seis familias (Tabla 1), lo que representa el 39% de las reportadas para el departamento del Atlántico por MONTERO et al. (2009) y el 64% de las reportadas por PRINCE et al. (2011). Este porcentaje de recolecta es bajo para el área de estudio, lo cual podría estar indicando que las especies reportadas son características de la época de muestreo o que el fragmento ha sufrido una pérdida de especies; debido a la disminución de la cobertura boscosa del fragmento, la tala de la vegetación circundante, el aumento de las zonas cultivables y de uso ganadero.
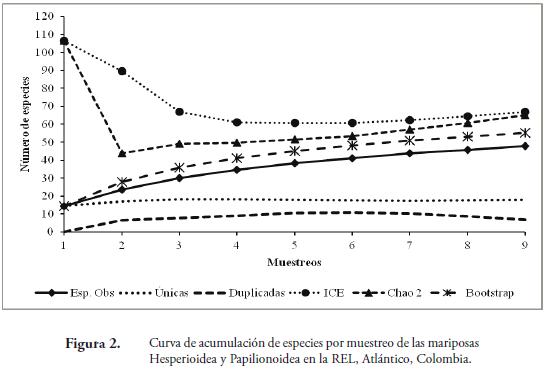
El número total de especies observadas (48), representó 71,71% del valor esperado según el estimador de riqueza ICE (67), el 73,85% para Chao 2 (65) y el 86,88% (55 especies) para el estimador Bootstrap (Figura 2). Teniendo en cuenta la riqueza calculada por los estimadores, se pueden encontrar entre 55 y 67 especies; demostrando que se pueden registrar mas especies para este fragmento (Figura 2). Esta riqueza podría aumentar con la ampliación de más muestreos, con el fin de cubrir la época en que se presenta el aumento de las precipitaciones en el lugar de estudio. Adicional a lo anterior, se debe cubrir una mayor extensión de la reserva, con el fin de tener un mayor número de hábitats representativos como la cima de los cerros, pastizales, ojos de agua; los cuales son lugares propicios para la recolecta de mariposas, ya que estos son frecuentados por mariposas como los licénidos las cuales no son atraídos con trampas con atrayente (PRIETO & DAHNERS, 2006). Lo anterior se ve reflejado en la no estabilización de las curvas para los estimadores utilizados, debido a que de las 48 especies recolectadas, 18 fueron únicas y siete duplicadas (Figura 2); observándose una tendencia de la curva para especies únicas a seguir aumentando. Las estimaciones de diversidad alfa obtenidas en este estudio, son muy parecidas al número de especies reportadas por MONTERO et al. (2009) para fragmentos de Bs-T de La Sierra y Guaibaná, en el departamento del Atlántico, cercanos a la REL, donde encontró 55 y 59 especies respectivamente. Resultados similares fueron obtenidos por VARGAS et al. (2011) en la Reserva Natural Las Delicias, en Santa Marta, Colombia; donde se realizaron cuatro faenas de captura, sin estabilización de las curvas de acumulación, probablemente debido al bajo número de muestreos.
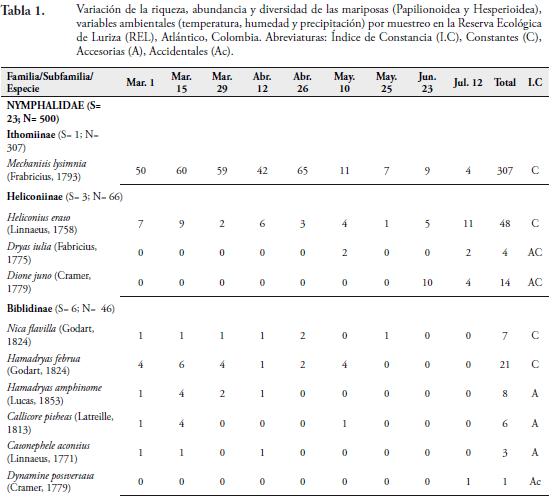
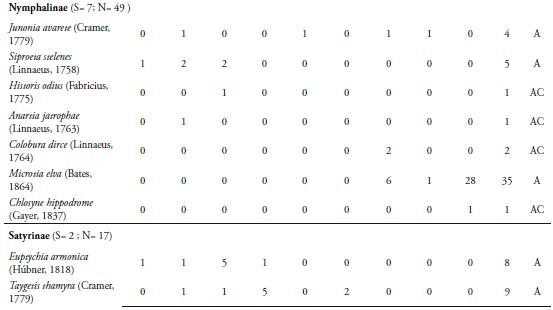
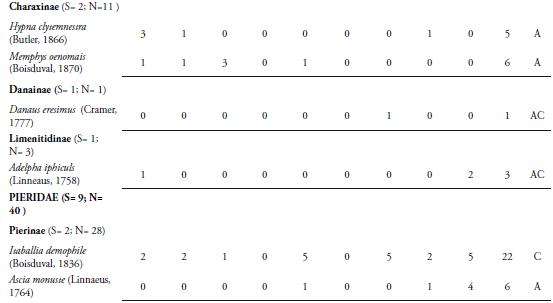
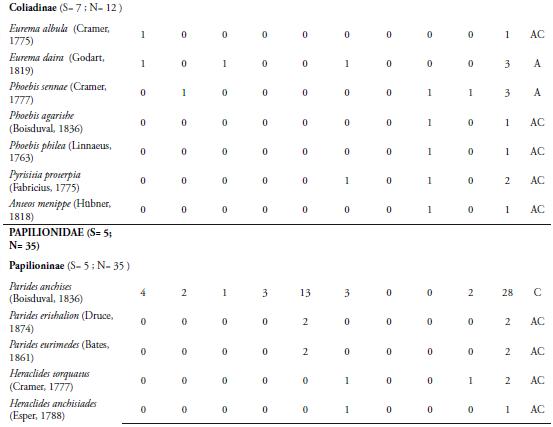
La familia Nymphalidae se caracterizó por ser la más representativa con nueve subfamilias, 23 especies y 500 individuos, representando el 44,9% de las especies recolectadas y el 81,4% del total de individuos capturado respectivamente (Tabla 1). El alto número de especies e individuos aportados por esta familia se explica porque este grupo de mariposas presenta una amplia riqueza de especies, que ocupan una diversidad de hábitats y por presentar amplios rangos de distribución (CHACÓN & MONTERO 2007, (DeVries et al. 1999). Además, la diversidad y abundancia de familias de plantas hospederas como Lauráceas, Sapindáceas, Asteráceas, passifloráceas, urticáceas, acantháceas entre otras en la zona, favorece la riqueza y abundancia de ninfálidos que sirven como sustrato para la ovipostura y alimento para sus larvas (CHACÓN & MONTERO, 2007). La familia Pieridae fue la segunda con mayor número de especies (nueve) y número de individuos (40) (Tabla 1). Esta familia incluye especies multivoltinas, que son de amplia distribución en el territorio y vuelan la mayor parte del año, lo cual hace más frecuente su recolecta, tal como se observó en la localidad de estudio (HERNÁNDEZ et al., 2008). Muchas de estas especies se capturaron en claros de bosque, a poca distancia del borde del fragmento y la matriz antrópica. Entre estas especies encontramos a Ascia monuste (Linnaeus, 1764), Phoebis sennae (Linnaeus, 1758), P. agarithe (Boisduval, 1836), P. philea (Linnaeus, 1763), Pyrisitia proterpia (Fabricius, 1775), Eurema albula (Cramer, 1775) y E. daira (Godart, 1819), Itaballia demophile (Linnaeus, 1763) y Anteos menippe (Hübner, 1818). Adicionalmente, especies como P. sennae, P. philea, E. albula y E. daira se capturaron en zonas abiertas y caminos que son utilizados por el ganado vacuno para llegar a la quebrada donde toman agua. La presencia del cuerpo de agua en el fragmento permite que estas especies entren a esta zona para alimentarse de frutas en descomposición y detritos. Además, es un lugar propicio para la presencia de estas especies libadoras de la arena húmeda y heces de mamíferos, de los cuales obtienen sales que requieren para su metabolismo (HERNÁNDEZ et al., 2008; VARGAS et al., 2011).
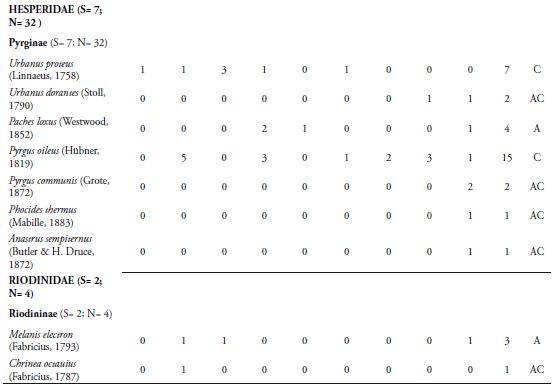
Las familias Lycaenidae y Riodinidae sólo aportaron el 4,16% de la riqueza, con 2 y 7 especies respectivamente. La poca representatividad de la primera familia, estaría relacionado por lo propuesto por JIMÉNEZ & HORTAL (2003), quienes afirman que la baja en la abundancia de las especies de Lycaenidae se debe por lo general a un sesgo en la captura; debido a sus coloraciones opacas y su tamaño pequeño o alta presencia de especies errantes. Además, hay que tener en cuenta que no se encontraron lugares de comportamientos "hilltopping", lo cual podría haber influido en la baja riqueza en este grupo de mariposas (PRIETO & DAHNERS, 2006). Por otro lado, los licénidos presentan densidades poblacionales muy bajas (LAMAS & PÉREZ, 1981), lo cual es una característica distintiva de las especies encontradas en el presente trabajo. Los riodínidos también presentaron densidades poblacionales muy bajas, con ciclos de vida univoltinos y suelen considerarse en su mayoría especies raras (HERNÁNDEZ et al., 2008), lo cual puede estar explicando los resultados arrojados en este estudio.
La especie más abundante fue Mechanitis lysimnia (Frabricius, 1793) (Subfamilia: Ithomiinae) con 307 individuos, con una dominancia del 50%. El resto de especies capturadas se encuentra por debajo del 8% (Tabla 1). La abundancia de M. lysimnia está directamente relacionada a las agregaciones que se forman en ciertos microhábitats que se encontraron alrededor de la quebrada (LAMAS & PÉREZ, 1981; RAMÍREZ et al., 2007). Los individuos de esta especie fueron recolectados cerca de la quebrada que atraviesa la reserva, donde se encuentra una vegetación con mayor cobertura boscosa y plantas hospederas como las del genero Solanum, que sirven como alimento y refugio a las larvas y adultos; lo cual podría estar explicando la mayor abundancia de esta especie en la REL. El mayor número de individuos recolectados de M. lysimnia (60 individuos) se presentó en ausencia de precipitaciones (Marzo 15), con valores de humedad relativa de 51.1% y temperatura de 32.7°C (Tabla 1); observándose muchos grupos de esta especie concentrados en ciertos puntos de la quebrada; lo cual facilitaba su captura, marcaje y liberación. Esta agregación es una adaptación de las especies de la subfamilia Ithomiinae a las condiciones climáticas en los periodos secos, en sitios con abundantes concentraciones de recursos alimenticios, plantas hospederas y en áreas sombreadas (PINHEIRO et al., 2008). Sin embargo, se encontró que a partir del sexto muestreo (Mayo 10) hubo una disminución de su abundancia, esto probablemente estaría relacionado a la disolución de los agregados que forma esta especie con la intensificación de las lluvias (PINHEIRO et al., 2008). VASCONCELLOS (1980) observó una fuerte reducción en las poblaciones de Ithomiinae en el sur de Sao Paulo (Brasil) después de la aparición de las lluvias en la región; lo cual coincide con los resultados obtenidos en este estudio. Por otro lado, FREITAS (1996) y PINHEIRO et al. (2008) en Brasil, encontraron que las asociaciones de Ithomiinae obedecen a factores climáticos, especialmente a una baja humedad relativa, responsable de constituir el ensamblaje. Estos resultados son diferentes a los reportados para otros fragmentos en el departamento por MONTERO et al. (2009), quien encontró que la especie más abundante fue Parides anchises (Boisduval, 1836) con 64 individuos y Prince et al. (2011), quien registra a Microtia elva (Bates, 1864) con 368 individuos. Estas diferencias en la abundancia entre los fragmentos podrían estar influidos por las características del lugar, tales como la composición y estructura vegetal, así como la época de muestreo.
La especie Heliconius erato (Linnaeus, 1758) fue la segunda más abundante (48) (Tabla 1), presentando un mayor número de individuos en el muestreo noveno (Julio 12) con 11 individuos, cuando los valores de precipitación fueron de 3.09mm, temperatura de 31.5°C y humedad de 68.4% (Tabla 1). Esta es una especie de amplia distribución en la región Neotropical y en el área de estudio se observo volando en los bosques y en lugares cercanos a bordes. Estas mariposas fueron más comunes durante los muestreos que presentaron bajos valores de precipitación, lo cual hizo posible la captura principalmente de hembras en actividad de oviposición (BROWN & YEPEZ, 1984)
La tercera especie más abundante (35) fue M. elva (familia Nymphalidae), la cual se recolectó durante los últimos tres muestreos, presentando un mayor número de individuos (28) en el noveno (Julio 12) (Tabla 1). Esta mariposa tiene la capacidad de usar una gran cantidad de recursos florísticos que se encuentran en el sotobosque, en la parte arbustiva y los claros cercanos a bordes y potreros. Esta oferta de recursos se presenta por la llegada de las lluvias en la zona, lo cual produce un cambio en la vegetación e induce al brote y desarrollo foliar de las plantas que utiliza este lepidóptero, tanto para la alimentación como para la reproducción. Además, esta mariposa presenta un vuelo lento, lo que también facilitó su captura en el área de estudio (PRINCE et al., 2011).
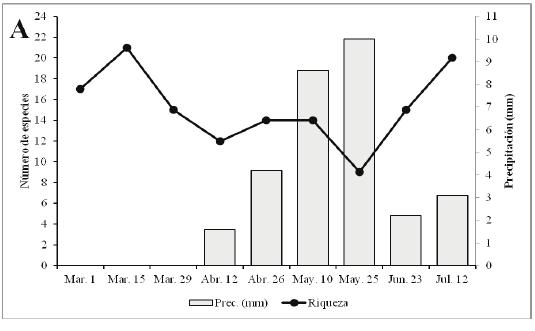
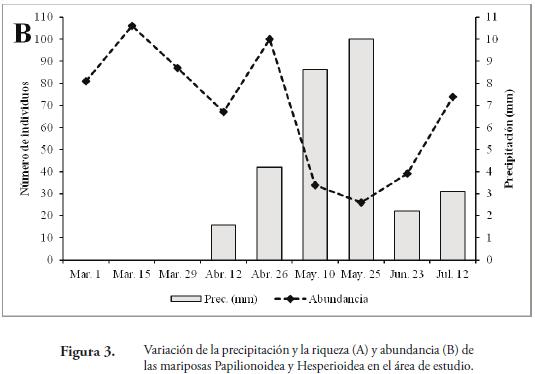
Los valores más altos de riqueza y abundancia se presentaron durante el segundo muestreo (Marzo 15) con 21 especies y 106 individuos, coincidiendo con el período de ausencia de lluvias en la zona y valores de temperatura de 32.7°C y humedad de 51.1% (Tabla 1, Figura 3AB). A partir del tercer muestreo, se observó una reducción en la riqueza y abundancia (Figura 3AB); sin embargo, en el quinto muestreo la abundancia aumentó levemente (100 individuos) originada por una alta proporción de individuos de M. lysimnia y P. anchises (Tabla 1). Durante el octavo (Junio 23) y noveno muestreo (Julio 12), los valores de precipitación disminuyeron, lo cual incidió en un aumento de la riqueza (15 y 20 especies) y la abundancia (39 y 74 individuos) de las mariposas (Tabla 1); cuando las precipitaciones presentaron los valores de 2.22 y 3.09mm respectivamente (Figura 3AB). Lo anterior estaría demostrando que las lluvias juegan un papel importante en la dinámica de la riqueza, abundancia y tasa de recambio de las mariposas Papilionoidea y Hesperioidea en este fragmento. Con la llegada de las lluvias, la actividad de muchas mariposas disminuye, porque imposibilitan el vuelo de los adultos y reducen el área de desplazamiento de las mismas. Además, al reducirse la cantidad de rayos solares que llegan al bosque, muchas se ven obligadas a buscar espacios abiertos fuera del fragmentos para realizar la termorregulación y otras actividades fisiológicas; lo cual puede estar explicando la disminución en las capturas de lepidópteros durante estos muestreos. Adicional a esto, durante la época de lluvias se observaron larvas y hojas con evidencia de haber sido consumidas por estos insectos; lo que sugiere que muchas de las especies no estaban representadas por formas adultas en la zona.
La mayor riqueza y abundancia durante la época seca en la zona contrastan con los resultados obtenidos por VARGAS et al. (2011) y PRINCE et al. (2011), quienes encontraron una mayor riqueza y abundancia de mariposas durante los periodos de lluvias; sugiriendo que en la REL se presenta un comportamiento distinto a otros fragmentos de Bs-T en la región Caribe. Esto puede atribuirse a las condiciones observadas en este fragmento de bosque en la reserva, tales como la presencia de una quebrada con agua permanente; lo cual puede incidir en la fenología de la vegetación circundante a esta (mantener un mayor follaje) y regular las condiciones microclimáticas (Temperatura y humedad moderada y menor intensidad de luz) durante la época seca en los estratos bajos del bosque ribereño y de esta forma puede influir en la dinámica de este grupo de insectos.
El Índice de Constancia demostró que el 52% fueron accidentales (Ac), el 31% se comportan como accesorias (A) y el 17% de las especies que se recolectaron fueron constantes (Tabla 1); destacándose M. lysimnia y H. erato por ser las más frecuentes (100%) durante todo los muestreos. El alto número de especies accidentales refleja que muchas de ellas presentan poblaciones muy bajas en el área, lo que sugiere la necesidad de realizar evaluaciones poblacionales de estas especies en la reserva, con el fin de determinar si se trata de especies raras. Las especies accesorias, únicas o duplicadas, se consideran que son migratorias, las cuales pueden estar desplazándose hacia el interior del bosque, desde zonas abiertas y cultivos de los alrededores de la reserva. Probablemente, los cambios microclimáticos y la búsqueda de alimento sean las causantes de que las especies de lepidópteros se desplacen entre el interior del bosque y la matriz antropogénica que rodea la reserva. Además, muchas especies de mariposas propias de interior de bosque también suelen presentar abundancias muy bajas; debido a la escasez o falta de plantas hospederas o el tamaño del área del fragmento. Además la riqueza y abundancia de insectos como las mariposas de un determinado lugar puede ser modificada por la composición de la matriz del hábitat circundante (Horner-Devine et al. 2003). Esta situación la encontró PINHEIRO et al. (2008) en fragmentos de Bosques cerrado y ribereño en el Campus Universitario Darcy Ribeiro, en Brasil. Sin embargo, esto resultados no coinciden con lo reportado por PRINCE et al. 2011; quien encontró que la mayoría de las mariposas encontradas eran accesorias, seguidas de especies accidentales y de igual forma encontraron que pocas especies son constantes, las cuales fueron las mismas reportadas en la REL. Estas diferencias se pueden presentar debido a que los fragmentos de Bs-T podría modular una dinámica diferente de especies y esto puede estar directamente relacionado a las condiciones propias de cada remanente de bosque, tales como la heterogeneidad vegetal, la altitud, el tamaño de fragmento y la variación de los parámetros ambientales, entre otras características.
Con respecto a la diversidad, los valores más alto de los números de Hill se presentaron en el último muestreo (Julio 12) para N1 (9,76) y para N2 (6,642) en el octavo muestreo (Junio 23) (Tabla 1), evidenciándose que diez especies son muy abundantes y siete son muy abundantes. Además, durante este periodo se observó una mayor actividad de mariposas en busca de recursos y parejas, probablemente debido a la disminución de las lluvias. Estos datos concuerdan con lo encontrado por VARGAS et al. (2011), quienes reportan los valores más altos en los números de Hill (N1 y N2) durante los meses de junio y julio en fragmentos de Bosque. En cuanto al Índice de Pielou, se encontró que los muestreos más equitativos fueron el 6 y 7, con valores cercanos a la unidad (0.85 y 0.86); lo cual coincidió con una mayor precipitación en la zona (Tabla 1), indicando que la mayoría de las especies capturadas aportan de manera más equitativa a la abundancia durante estos muestreos. Adicional a esto, existen lepidópteros que requieren ciertas condiciones que se dan únicamente en esta época del año, como mayor heterogeneidad de hábitat, disponibilidad de recursos, aumento de la actividad de muchas especies y condiciones ambientales estables.
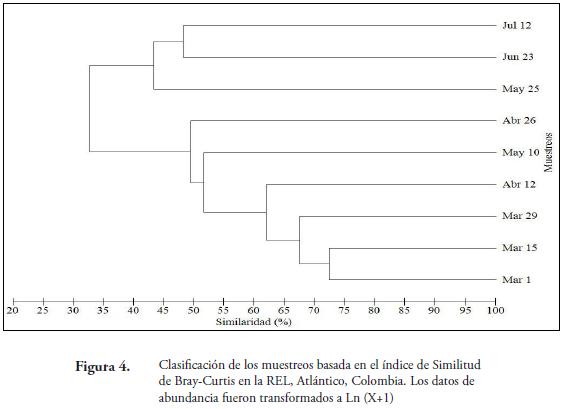
El análisis de agrupamiento de Bray–Curtis, demostró que los tres primeros muestreos presentaron una similitud mayor del 65%, los cuales coinciden con las ausencias de lluvias en la reserva (Figura 4). Esto puede estar relacionado a que en los bosques tropicales se mantiene una mayor similitud en la riqueza y abundancia de especies de mariposas diurnas durante los periodos secos y ésta tiende a variar a medida que inician las precipitaciones (VARGAS et al., 2011).
Los muestreos más disímiles fueron los realizados en mayo, junio y julio, lo cual se puede atribuir a un descenso abrupto de la precipitación pasando de valores altos (10mm en Mayo 10) a valores inferiores (2.2mm en junio y 3.09mm en julio). Lo anterior incide en los cambios en las comunidades de lepidópteros, ya que las variaciones de la precipitación no solo pueden afectar la estructura vegetal, sino que también induce a cambios en las variables ambientales, como la humedad. De igual forma, esta agrupación que se forma por la presencia de las lluvias, guarda relación con la riqueza y abundancia de mariposas, ya que este factor afecta la reproducción, el desarrollo ontogenético y la conducta de los individuos adultos (WOLDA, 1988). Adicionalmente, la presencia o ausencia de lluvias seria una de las condiciones que estaría favoreciendo el aumento o disminución en la abundancia de algunas especies de mariposas Papilionoidea y Hesperioidea en este fragmento de bosque. En este caso, las precipitaciones son un factor ambiental que modela la fenología del bosque seco y por consiguiente la disponibilidad de refugio y alimento necesario para estos insectos (VARGAS et al., 2011). Aunque este factor, también puede afectar indirectamente factores biológicos como mecanismos de alimentación, competencia por alimento, relaciones depredador-presa, los cuales juegan un papel importante en la dinámica temporal de este grupo de insectos en la zona. Además, la presencia de especies vegetales aledañas a la quebrada puede ser considerada como agregadores de biodiversidad de este grupo de lepidópteros; debido a que dan complejidad al hábitat y favorecen el establecimiento de algunas especies en la época seca y otras en la de lluvias, lo cual también estaría explicando las diferentes agrupaciones que se observaron en la comunidad de Hesperioidea y Papilionoidea en la REL (Figura 4)
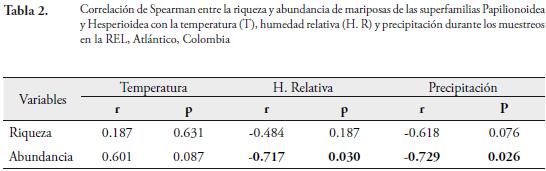
El análisis de correlación de Spearman demostró que variables ambientales como la precipitación y humedad relativa explican la variación temporal de la riqueza y abundancia de las mariposas Hesperioidea y Papilionoidea en el área de estudio durante el tiempo de muestreo. Se determinó una correlación inversa y significativa entre la abundancia con la precipitación (r=-0.72; p=0.026) y la humedad relativa (r=-0,71; p=0,030); mientras que con la temperatura la correlacion no fue significativa (p≥ 0.05). Por otro lado, se determinó una correlación inversa, pero no significativa entre la riqueza y precipitación (r=-0.618; p=0.07); presentándose la misma tendencia con la temperatura y la humedad relativa (Tabla 2). Estos resultados posiblemente podrían demostrar que el aumento de la abundancia de lepidópteros encontrados en la reserva está relacionado con los bajos valores de precipitación en la zona. En esta época, aumenta la proliferación de frutos (Mangifera indica y Spondias mombin) que caen al suelo donde se descomponen. Esta situación favorece un aumento de recursos alimenticios que son aprovechados por algunas especies de mariposas, las cuales pueden encontrarse en diferentes microhábitats en la hojarasca, cuando los valores de humedad son muy bajos y estén beneficiándose del estrés hídrico y las mayores intensidades lumínicas durante la época seca en el fragmento de bosque. Además, la escasez de follaje hace que estos insectos sean más visibles, por lo cual aumenta la probabilidad de ser capturados. Con respecto a la temperatura, es que tenga un efecto mínimo en la comunidad de mariposas, ya que fue un parámetro que vario muy poco en el área entre cada muestreo (Tabla 1); debido a que la presencia de la quebrada contribuye a mantener constante las condiciones de temperatura durante las faenas de captura. Lo anterior indica que esta variable no explica la variación de la fauna de lepidópteros en la REL; mientras que la humedad relativa fue un parámetro clave en la abundancia de estas mariposas, pues influye en el desarrollo del ciclo de vida de este grupo de insectos (PALANCA, 1987).
Finalmente, pese al aumento de las fronteras agrícolas y ganaderas en los alrededores de la reserva; ésta conserva una fauna importante de mariposas Papilionoidea y Hesperioidea de los bosques secos del Caribe colombiano. También se pudo observar que muchas especies de mariposas presentan una mayor abundancia durante la época seca, cuando se presentan recursos alimenticios; lo cual aporta los nutrientes necesarios para llevar a cabo sus funciones fisiológicas.
CONCLUSIONES
La variación temporal de la abundancia de mariposas Papilionoidea y Hesperioidea en la REL está sincronizada con los valores de precipitación, evidenciándose una mayor numero de mariposas en la época seca, lo cual es un patrón de comportamiento distinto a la de otras áreas del departamento del Atlántico. El ensamblaje de mariposas diurnas varió de acuerdo a las condiciones ambientales que se presentan en el parche de Bs-T, además la heterogeneidad de hábitat que se mantuvo en el fragmento durante la época seca por la presencia de la quebrada ofreció condiciones favorables para los lepidópteros. La curva de acumulación de especies muestra la tendencia a seguir apareciendo más especies de mariposas en la REL, por lo que se hace recomendable evaluar la fauna lepidópteros por medio de la realización de más muestreos. Finalmente, con este trabajo se hace necesario un monitoreo continuo de la fauna de mariposas en ausencias y presencias de lluvias y establecer un sistema de protección que involucre a la comunidad de campesinos de la zona.
AGRADECIMIENTOS
A la comunidad de Luriza por facilitar la estancia en este lugar, durante la realización del trabajo de campo. Al señor Manuel Amaranto, su hija Doris y demás familiares por acogernos en su hogar y colaborarnos con la alimentación. A Jaime Padilla, por el transporte al área de estudio. También a la ONG Usiacurí Verde por permitir realizar el trabajo en la reserva. A los miembros del semillero de investigación NEOPTERA del programa de Biología, especialmente a Karina Rodríguez, Alexis Navarro y a Karen Muñoz por su colaboración en trabajo de campo y la Facultad de Ciencias Básicas de la Universidad del Atlántico por facilitar los permisos y las instalaciones del laboratorio de Zoología para el procesamiento e identificación de las muestras.
BIBLIOGRAFÍA
ANDRADE, M.G., 1998.- Utilización de las mariposas como bioindicadoras del tipo de hábitat y su biodiversidad en Colombia. Rev. Acad. Colomb. Cienc., 22 (84): 407-421. [ Links ]
ANDRADE, M.G., 2002.- Biodiversidad de las mariposas (Lepidoptera: Rhopalocera) de Colombia. (en) COSTA, C.; VANIN, S.A. & MELIC, A. (eds.) Proyecto de Red Iberoamericana de Biogeografía y Entomología Sistemática. Colombia [ Links ]
BODENHEIMER, F.S., 1955.- Precis d écologie animale. PAYOT. Paris. [ Links ]
BONEBRAKE, T.C., PONISIO, L.C., BOGGS, C., & EHLRICH, P., 2010. - More than just indicators: A review of tropical Butterfly ecology and conservation. Biological Concervation 143: 1831-1841. [ Links ]
BROWN, K.S., 1991.- Conservation of neotropical paleoenvironments: Insects as indicators (en) COLLINS, N. M. & THOMAS, J. A. (eds.) Conservation of Insects and their Habitats. Press, London [ Links ]
BROWN, K.S. & YEPEZ. F., 1984.- Los Heliconiini (Lepidoptera, Nymphalidae) de Venezuela Boletín de Entomología Venezolana 3(4):29-73. [ Links ]
CAMERO, E. & CALDERÓN, C., 2007.- Comunidad de mariposas diurnas (Lepidóptera: Rhopalocera) en un gradiente altitudinal del Cañón del Río Combeima – Tolima, Colombia. Acta biol. Colom., 21 (2): 95-110. [ Links ]
CAMPO, L & ANDRADE M.G., 2009.- Lepidópteros (Papilionoidea y Hesperioidea) asociados a bosques seco tropical del caribe colombiano, un estudio de caso en el bosque el Aguil, Aguachica. Cesar. Colombia, 615-631 (en) Rangel J.O (ed). Colombia. Diversidad Biotica VIII. Media y baja de la serranía de Perijá Instituto de Ciencias Naturales Universidad nacional de Colombia, Bogotá [ Links ].
CHACÓN, I. & MONTERO, J., 2007.- Mariposas de Costa Rica. Editorial INBio. Costa Rica. [ Links ]
CLARKE, K.R. & WARWICK, R.M., 2001.- Change in Marine Communities: an Approach to Statistical Analysis and Interpretation, 2nd.ed. Primer-E Ltd, Plymouth, UK. [ Links ]
COLWELL, R., 2009.- Estimates: Statistical Estimation of Species Richness and Shared Species from Samples, Version 8.2. [ Links ]
CORPORACIÓN AUTÓNOMA REGIONAL DEL ATLÁNTICO, CRA., 2011. Distrito regional de manejo integrado Luriza, municipio de Usiacurí. 12p. http://www.crautonoma.gov.co/documentos/areasprotegidas/normatividad/Distrito%20manejo%20integrado%20Luriza.pdf [ Links ]
DÍAZ, J.M., 2006.- Bosque Seco Tropical Colombia. Banco de Occidente Credencial. Cali Colombia. [ Links ]
DEVRIES, PJ; WALLA, T.H. & GREENEY, H.F., 1999.- Species diversity in spatial and temporal dimetions of fruit-feeding butterfly from tow Ecuadorian rainforest Forest. Biol. Jour.of the Linn. Soc. 68: 333-353. [ Links ]
ERAZO, M.C. & GONZÁLEZ, L.A., 2008. -Mariposas, pp. 28-116 (en): Rodríguez, J.V.; Rueda, T.D., & Gutierrez H. (ed.), Guía ilustrada de la fauna del Santuario de Vida Silvestre Los Besotes, Valledupar, Cesar, Colombia. Bogotá, Serie de guías tropicales de campo N° 7, Conservación Internacional-Colombia. Editorial Panamericana, Formas e Impresos [ Links ]
FREITAS, V.L., 1996.- Population biology of Heterosais Edessa (Nymphalidae) and its associated Atlantic Forest Ithomiinae community. Lepid. Soc. J., 50: 273–289. [ Links ]
HAMMER, Ø.; HARPER, D.A. & RYAN P.D., 2001.- PAST: Paleontological Statistics Software Package for Education and Data Analysis Palaeontología Electrónica. Vol. 4(1). [ Links ]
HERNÁNDEZ, J., 1992.- Caracterización geográfica de Colombia (en): Halffter G. (ed.). La Diversidad Biológica de Iberoamérica I CYTED-B Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo. Instituto de Ecología, A.C. México. [ Links ]
HERNÁNDEZ, C., LLORENTE, J.; MEJÍA, I. & LUIS, A., 2008.- Las mariposas (Hesperoidea y Papilionoidea) de Malinalco, Estado de México. Rev. Mex. Biod., 79: 117-130. [ Links ]
HORNER-DEVINE, M.C, DAYLI, G.C, EHRLICH, P.R & BOGGS, C.L., 2003.-Countryside Biogeography of Tropical Butterflies. Con. Biol. (17): 168–177 [ Links ]
INSTITUTO ALEXANDER VON HUMBOLDT, IAVH., 1998.- El Bosque seco Tropical (Bs-T) en Colombia. Grupo de Exploraciones y Monitoreo Ambiental GEMA. Villa de Leyva, Colombia. [ Links ]
JIMÉNEZ, A. & HORTAL J., 2003.- Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Rev. Iber. Arac., 8: 151-161. [ Links ]
KREMEN, C., 1992.- Assessing the Indicator Properties of Species Assemblages for Natural Areas Monitoring. Ecol Appl., 2(2):203-217. [ Links ]
LAMAS, G. & PÉREZ, E., 1981.- Danainae e Ithominae (Lepidoptera, Nymphalidae) atraídos por Heliotropium (Boraginacea) en madre de Dios, Peru. Rev. Per. Ent., 24 (1):59-62. [ Links ]
LE CROM, J.F.; CONSTANTINO, L.M. &. SALAZAR, J.A., 2002.- Mariposas de Colombia. T. I: Papilionidae. Carlec Ltda. Colombia. [ Links ]
LE CROM, J.F.; CONSTANTINO, L.M. &. SALAZAR, J.A., 2004.- Mariposas de Colombia. Tomo II: Pieridae. Carlec Ltda. Colombia. [ Links ]
LLORENTE, J.; MARTÍNEZ, A.; VARGAS, I.; SOBERÓN, J., 1993.-Biodiversidad de las Mariposas: Su Conocimiento y Conservación en México. Rev. Soc. Mex. Hist. Nat., Vol. Esp. (XLIV): 313-324. [ Links ]
MARTÍNEZ, N.; GARCÍA, S.; GUTIÉRREZ, M.; SANJUÁN, S. & CONTRERAS, C., 2010.- Composición y estructura de la fauna de escarabajos (Insecta: Coleóptera) atraídos por trampas de luz en la Reserva Ecológica de Luriza, Atlántico, Colombia. SEA., 47: 373-381. [ Links ]
MILES, L.; NEWTON, A.C.; DEFRIES, R.S.; RAVILIOUS, C.; MAY, I.; BLYTH, S.; KAPOS, V. & GORDON, J.E., 2006.- A global overview of the conservation status of tropical dry forests. Journal of Biogeography., 33: 491-505. [ Links ]
MONTERO, F.; MORENO, M. & GUTIÉRREZ, L.C., 2009.- Mariposas (Lepidoptera: Hesperioidea y Papilionoidea) asociadas a fragmentos de Bosque Seco Tropical en el departamento del Atlántico, Colombia. Bol. Cient. Mus. Hist. Nat., 13(2): 157-173. [ Links ]
MORENO, C.E., 2001.- Métodos para medir la biodiversidad. M&T–Manuales y Tesis SEA, vol.1. Zaragoza. [ Links ]
PALANCA, A., 1987.- Aspectos faunísticos y ecológicos de lepidópteros altoaragonenses. Monografías del Instituto Pirenaico de ecología. No. 2. Jaca. [ Links ]
PENNINGTON, R.T.; LEWIS, G.P. & RATTER, J.A., 2006.- Neotropical Savannas and Seasonall y Dry Forests: Plant Diversity, Biogeography, and Conservation. Boca Raton, FL: Taylor and Francis. [ Links ]
PINHEIRO, C.; MEDRI, I. & MOREIRA, A., 2008.- Why do the ithomiines (Lepidoptera, Nymphalidae) aggregate? Notes on a butterfly pocket in central Brazil. Rev. Bras. Entom., 52(4): 610-614. [ Links ]
POLLARD, E. & YATES, T.J., 1994.- Monitoring butterflies for Ecology and Conservation. London: Chapman y Hall. [ Links ]
PRANCE, W., 2006.- Tropical savannas and seasonally dry forests: an introduction. Journal of Biogeography., 33:385-386. [ Links ]
PRIETO, C.; TAKEGAMI C.; RIVERA, M.J., 2005.- Estructura poblacional de Morpho sulkowskyi Kollar, 1850 (Lepidoptera: Nymphalidae) en un sector de la cordillera occidental, departamento del Cauca (Colombia). ENTOMOTRÓPICA., Vol. 20 (1): 15-22. [ Links ]
PRIETO, C. & DAHNERS, H. 2006.- Eumaeini (Lepidoptera: Lycaenidae) del cerro San Antonio: Dinámica de la riqueza y comportamiento de "Hilltopping". Revista Colombiana de Entomología 32:2. 179-190. [ Links ]
PRINCE, E.; VARGAS, M.A.; SALAZAR, J. & MARTÍNEZ, N.J., 2011.- Mariposas Papilionoidea y Hespeoidea (Insecta: Lepidoptera) en dos fragmentos de Bosque Seco Tropical en Corrales de San Luis, Atlántico, Colombia. SEA., 48: 243-252. [ Links ]
RAMÍREZ, L.; CHACÓN, P. & CONSTANTINO, L.M., 2007.- Diversidad de mariposas diurnas (Lepidoptera: Papilionoidea y Hesperioidea) en Santiago de Cali, Valle del Cauca, Colombia. Rev Colomb Entomol., 33:54-63. [ Links ]
RUIZ, J. & FANDIÑO, M.C., 2009.- Estado del bosque seco tropical e importancia relativa de su flora leñosa, islas de la Vieja Providencia y Santa Catalina, Colombia, Caribe suroccidental. Rev. Acad. Colomb. Cienc., 33(126): 5-15. [ Links ]
SACKIS, G.D. &. MORAIS, A.B., 2008.- Butterflies (Lepidoptera: Hesperioidea and Papilionoidea) from Universidade Federal de Santa Maria campus, Santa Maria, Rio Grande do Sul. Biota Neotrop., 8, no 1. [ Links ]
SHAHABUDDIN, G. & PONTE, C.A., 2005.- Frugivorous butter?y species in tropical forest fragments: correlates of vulnerability to extinction. Biodiversity and Conservation 14: 1137–1152. [ Links ]
SILVEIRA, S.; NAKANO, O.; BARBIN, D. & VILLA, N.A., 1976.- Manual de Ecologia de Insetos. São Paulo: Ceres. [ Links ]
SIMANCA, R. & MARTINEZ, N. J., 2010. Nueva técnica para evaluar la estratificación vertical de hormigas (Hymenoptera: Formicidae) en el bosque seco tropical, Colombia. Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), 46: 311-318. [ Links ]
VARGAS, M.A.; MARTÍNEZ, N.J.; GUTIÉRREZ, L. & PRINCE, S., 2011.- Riqueza y abundancia de Hesperoidea y Papilionoidea (Lepidóptera) en la Reserva Natural Las Delicias, Santa Marta, Magdalena, Colombia. Acta Biol. Colomb., 16(1):43-60. [ Links ]
VASCONCELLOS, J. 1980.- Dinâmica de populações de Ithomiinae (Lepidoptera, Nymphalidae) em Sumaré, São Paulo. Ph. D. dissertation, Campinas, Unicamp. [ Links ]
VILLAREAL, H.; ÁLVAREZ, M.; CÓRDOBA, S.; ESCOBAR, F.; FAGUA, G.; GAST, F.; MENDOZA, H.; OSPINA, M. & UMAÑA, A.M., 2004.- Manual de métodos para el desarrollo de inventarios de biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, Colombia. [ Links ]
WARREN, A. D.; DAVIS, K. J.; STANGELAND, E. M.; PELHAM, J. P. & GRISHIN, N. V. ,2012. Illustrated Lists of American Butterflies. [05-V-12]. http://www.butterfliesofamerica.com/ [ Links ]
WOLDA, H. 1988.- Insect seasonality. Annu. Rev. Ecol. Syst., 19: 1-18. [ Links ]
