










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.18 no.2 Manizales July/Dec. 2014
ASPECTOS ECOLÓGICOS DE LAS AVES MIGRATORIAS NEÁRTICAS EN EL CAMPUS DE LA UNIVERSIDAD DEL VALLE*
ECOLOGICAL ASPECTS OF NEARTIC MIGRANT BIRDS IN THE UNIVERSIDAD DEL VALLE CAMPUS
Juan David Ardila-Téllez1, Lorena Cruz-Bernate2
* FR: 4-VII-14. FA: 10-I-2015.
1 Departamento de Biología, Universidad del Valle, Calle 13 No. 100-00, A. A. 25360, Cali, Colombia. E-mail: juan.david.ardila@hotmail.com.
2 Departamento de Biología, Universidad del Valle,Cali, Colombia. E-mail: lorena.cruz@correounivalle.edu.co.
CÓMO CITAR: ARDILA-TÉLLEZ, J.D. & CRUZ-BERNATE, L., 2014.- Aspectos ecológicos de las aves migratorias neárticas en el campus de la Universidad del Valle. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 18 (2): 93-108.
Resumen
Debido a la destrucción y fragmentación de hábitats prístinos, expansión urbana y ausencia de información base, se hace prioritario investigaciones sobre ecología de la comunidad de aves. Las zonas arbóreas urbanas constituyen hábitat de apoyo permanente para el mantenimiento y conservación de la avifauna local y continental. Los estudios de aves migratorias neárticas en Colombia se enfocan en la determinación de la riqueza, pero carecen de detalles ecológicos sobre su dinámica migratoria. El objetivo fue determinar aspectos ecológicos de la migración de aves neárticas en un hábitat urbano, como son: fechas de llegada y partida; abundancia absoluta; proporción de sexos; variación de los depósitos de grasa e interacciones con residentes. Entre septiembre de 2011 y mayo de 2012 se hicieron recorridos de observación y capturas con redes de niebla en el campus de la Universidad del Valle a 990 msnm. El primer y último registro se efectuó el 20 de septiembre de 2011 yel 5 de mayo de 2012. Se registraron 13 especies de aves migratorias, de las cuales Setophaga petechia y Piranga rubra presentaron el mayor índice de importancia relativa (IIR=2,386 y IIR=1,136, respectivamente). Hay diferencias por sexo y especie en los patrones de llegada y partida a los territorios de invernada. No se encontró correlación entre peso y tiempo de permanencia de las aves y se describe la variación de las reservas energéticas a través de la temporada. Se demuestra el uso de áreas arbóreas dentro de las zonas urbanas como sitios importantes de estadía temporal para las aves migratorias.
Palabras clave: depósitos de grasa, interacciones interespecíficas, ornitología urbana,patrones de llegada, sexo, territorios de invernada, tiempos de permanencia.
Abstract
Due to the destruction and fragmentation of pristine habitats, urban sprawl and lack of basic information, it is necessary to research on community ecology of birds. Urban arboreal areas represent permanent arboreal habitats that support,and maintain conservation of local and continental avifauna. The studies on Nearctic migratory birds in Colombia are focused on the determination of richness but there is a lack on ecological details about their migration dynamics. The objective of this study was to determine ecological aspects of Neartic birds in an urban habitat such as arrival and departure dates, abundance, sex ratio, variation in fat deposits and interactions with residents. Between September 2011 and May 2012, bird watching and capture tours were carried out using mist nets on the Universidad del Valle campus at 990 meters above sea level. The first and last entries were done on September 20, 2011 and May 5, 2012, respectively. A total of 13 migratory bird species were recorded from which Setophaga petechia and Piranga rubra displayed the highest relative importance index (IIR = 2.386 and1.136, respectively). Differences in sex and species patterns were observed on the arrival and departure toward wintering areas. No correlation was found between weight and length of stay of birds, and the variation of energy reserves through the season is described. The study demonstrates importance of the arboreal sites, within urban areas, as temporary harbor for migratory birds.
Key words: arrival fat deposits, interspecific interactions, urban ornithology, arrival patterns, residence time, sex, wintering territories, time of stay
INTRODUCCIÓN
Colombia cuenta con una favorable ubicación geográfica que la convierte en un paso obligatorio para gran variedad de aves migratorias neárticas (OCAMPO-PEÑUELA, 2010), ellas lo usan como lugar de residencia de inverno o como zona de abastecimiento energético para continuar su recorrido por Suramérica (MORENO, 2009). Cerca del 10% de la avifauna colombiana corresponde a especies migratorias del hemisferio norte, estas especies se distribuyen en 28 familias, enlas que las reinitas (Parulidae), los playeros (Scolopacidae), las gaviotas (Laridae) y los atrapamoscas (Tyrannidae) poseen el mayor número de especies con el 54% del total (MORENO, 2009). De acuerdo con HILTY & BROWN (2001), es posible encontrar 146 especies de aves que se reproducen en la zona templada del norte y que llegan a Colombia como migratorias de larga distancia; de estas, ocho tienen poblaciones residentes que se reproducen en Colombia y seis son especies cuyo estatus como migratoria del norte no está comprobado. Respecto a la preferencia en el uso de diferentes hábitats, OREJUELA et al. (1980) realizaron una comparación de la comunidad de aves migratorias en tres tipos de bosque en el Valle del Cauca, encontrando que en el bosque húmedo tropical la utilización del hábitat por parte de las aves migratorias fue nula, ya que no se observó ningún individuo a diferencia de los otros tipos de bosques donde las aves fueron observadas, el promedio de aves migratorias fue de 17,15%, probablemente el efecto competitivo de especies especialistas locales es bastante alto impidiendo que las especies migratorias usen dichoshábitats, restringiéndolas a hábitat intervenidos.
Desde hace algunas décadas se ha detectado una disminución en las poblaciones de aves migratorias neárticas (RAPPOLE, 1994; FUNDACIÓN PROAVES, 2009), considerándose como causas: la pérdida del hábitat de reproducción; fragmentación; degradación de hábitat; y pérdidas de sitios de paso intermedio y de invernada en los trópicos (FINCH, 1991; RAPPOLE, 1995). Adicionalmente, la urbanización que es uno de los crecientes fenómenos que rigen el uso del suelo a nivel mundial, aumenta la fragmentación del hábitat y provoca diferentes grados de amenaza tanto a especies residentes como migratorias (ORTEGA-ÁLVAREZ et al., 2011).
Conocemos que la conservación de los bosques continuos contribuyen al sostenimiento de las poblaciones de avifauna silvestre, pero las áreas de vegetación urbana se han constituido también en zonas de gran relevancia para este fin (ANGARITA, 2002; REYES-GUTIÉRREZ et al., 2002; RIVERA-GUTIÉRREZ, 2002; MUÑOZ et al., 2007; DAGMA, 2010). Actualmente, la ornitología urbana en Latinoamérica presenta un marcado aumento (ORTEGA-ÁLVAREZ et al., 2011), enfocándose en parques o zonas verdes con algún grado de arborización que han sido establecidos con fines recreativos o como objeto de conservación (RIVERA-GUTIÉRREZ, 2002). Estas áreas urbanas albergan no solo una alta riqueza de especies residentes y migratorias, sino también abundancia de ellas, cuyos individuos encuentran los hábitats y recursos necesarios para la supervivencia y reproducción (MACGREGOR-FORS & ORTEGA-ÁLVAREZ, 2013).
A pesar de que el estudio de la migración en el Neotrópico se ha venido incrementando desde hace un tiempo (OREJUELAet al., 1980; STILES, 1994), todavía existen vacíos de información referente a la abundancia, distribución invernal, ubicación de sitios de parada, estrategias y rutas migratorias fuera de Norte América, esto es importante en vista de los rápidos cambios en el hábitat que ocurren en elNeotrópico (RAPPOLE, 1995;GÓMEZet al.,2010). La gran mayoría de trabajos de ecología urbana se han enfocado en realizar los listados de las aves presentes en determinado lugar, no obstante, faltan temas como la fenología de la migración.
El objetivo de este estudio fue determinar los tiempos de arribo y permanencia de las especies migratorias, su abundancia, proporción sexual, estado de las reservas energéticas e interacciones entre estas y las residentes en el campus de la Universidad del Valle.
MÉTODOS
Área de estudio
Este estudio se llevó a cabo entre septiembre de 2011 y mayo de 2012, en el campus de la Universidad el Valle (desde ahora "el campus"), departamento del Valle del Cauca (3° 22' N, 76° 32' W, 990 msnm), con precipitación promedio anual que varía entre 900 y 1800 mm y una temperatura promedio de 24°C. Según la clasificación de Holdridgese ubica en la zona de vida de bosque seco tropical (bs-T) (ESPINAL, 1968). El campus cuenta conun área, aproximadamente, de 100 ha, de las cuales 8,5 ha se encuentran ocupadas por edificaciones, 44 ha por árboles, 45,5 ha por prados y pastizales y 1 ha por dos lagos.Entre las especies de árboles más comunes se encuentran los chiminangos (Pithece llobium dulce), samanes (Samanea saman), mangos (Mangifera indica), tulipanes africanos (Spathodea campanulata), cauchos (Ficus elastica) y guayacanes amarillo y rosado (Tabebuia chrysantha y T. rosea, respectivamente) (MUÑOZ et al., 2007).
Métodos de campo
Se hicieron tres recorridos semanales de observación de ruta fija a una velocidad constante y con longitud de 4,8 km aprox. La duración de cada uno fue de cuatro horas y se efectuaron entre 06:00 y 10:00 horas, 10:00 y 14:00 horas y 14:00 y 18:00 horas. De cada especie observada se determinó el número de individuos,sexo (en aquellas dimórficas), altura sobre el suelo, comportamiento (alimentándose, descansando, acicalándose, entre otros) e interacciones agresivas (ataque directo, vuelo de suplante, persecución al vuelo, y demás) tanto intercomo intraespecíficas.
Complementario a los recorridos y en días distintos se instalaron nueve redes de niebla, de 9 m largo, que cubrieron 3 m de altura durante 12 horas por semana. Las redes fueron revisadas a intervalos de 30 minutos por seguridad de las aves y estuvieron abiertas en dos jornadasentre las 06:00 y las 12:00 horas, así como entre las 12:00 y las 18:00 horas.
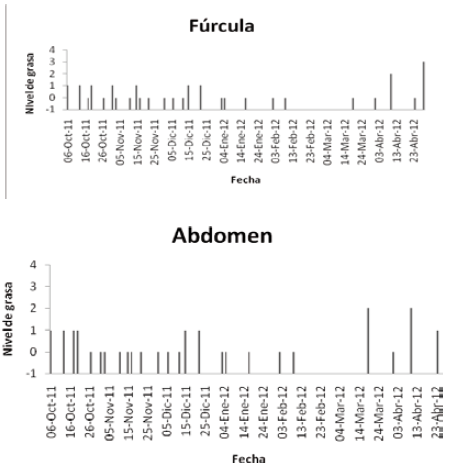
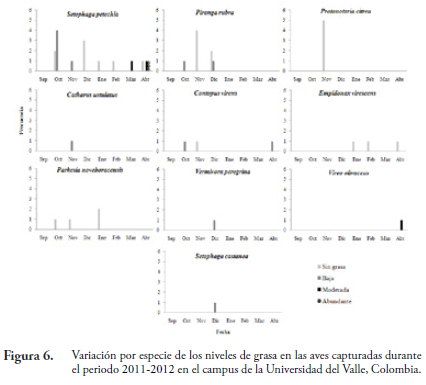
Cada individuo capturado se identificó hasta especie, utilizando la guía de aves de Norte América (ROBBINS et al., 2001) y la guía de las aves de Colombia (HILTY & BROWN, 2001). Se examinó para determinar la presencia de grasa subcutánea en fúrcula, flancos y abdomen con las siguientes categorías: sin grasa (nivel 0); grasa baja (1); grasa moderadas (2); y grasa abundante (3) (NORTH AMERICAN BANDING COUNCIL, 2001) (Fig. 1). Se marcó con anillostarsales de colores (Darvic®) en combinaciones únicas y se midió morfométricamente (ala, culmen, rectrices, tarso y peso).
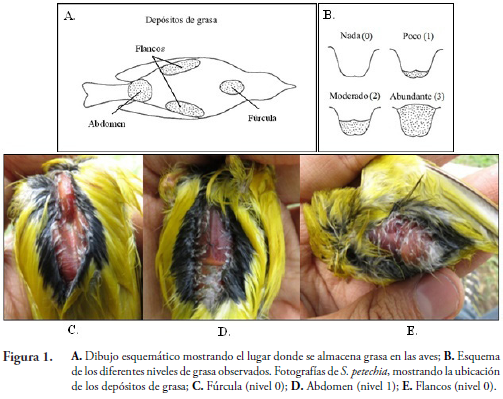
Se estableció la fecha de arribo y partida de las diferentes especies migratorias por medio de la observación diaria dentro del campus. La abundancia absoluta se calculó como número de aves migratorias registradas por jornada de observación a través de la temporada y se calculó el índice de importancia relativa (IIR), según JOSENSet al. (2012) como: IIR= (Ni/Nt)* (Mi/Mt)* 100, donde Ni corresponde al número de individuos de la especie "i", Ntel total de especies, Mi el número de jornadas de observación en donde la especie "i" fue observada y Mt el número total de jornadas de observación.
Se analizó la proporción de sexos por especie, en aquellas dimórficas, al inicio y final de la temporada de migración. Se determinó si existía relación entre el peso de las aves y el tiempo de permanencia en los territorios de invernada por medio del índice de correlación de Spearman (ZAR, 2010), con la especie que presentó el mayor número de datos (S. petechia). Con base en la similitud relativa de observaciones registradas sobre hábitos alimentarios e interacciones entre individuos se comparó la altura de forrajeo de las dos especies de aves migratorias, con más registros como son S. petechia (insectívora) y P. rubra (frugívora), con la de dos especies residentes del campus: Coereba flaveola (nectarívora/insectívora)y Thraupis episcopus (frugívora).
RESULTADOS
Permanencia, abundancia y sexo
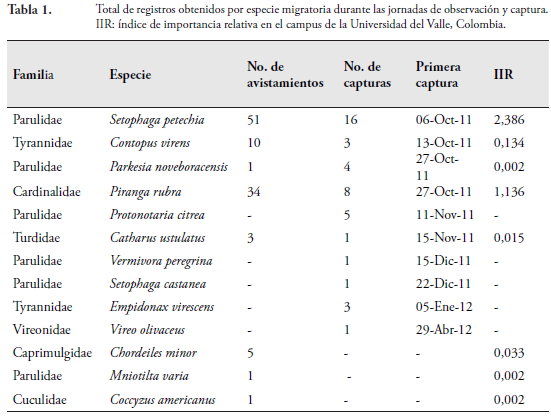
Se registró un total de 13 especies migratorias neárticas en el campus de la Universidad del Valle con un total de 148 registros, 105 de los cuales corresponden a avistamientos y 43 a individuos capturados con redes de niebla. Las especies identificadas pertenecieron a seis familias: Parulidae, Tyrannidae, Cardinalidae, Turdidae, Vireonidae y Caprimulgidae, donde aquella con mayor número de registros fue Parulidae (Tabla 1).
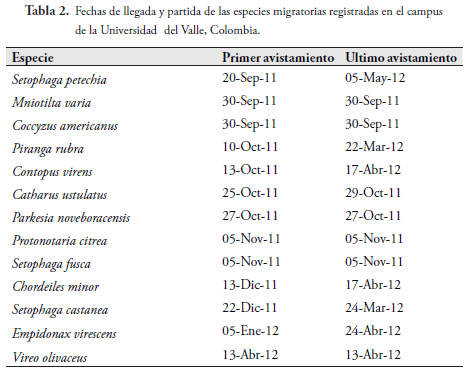
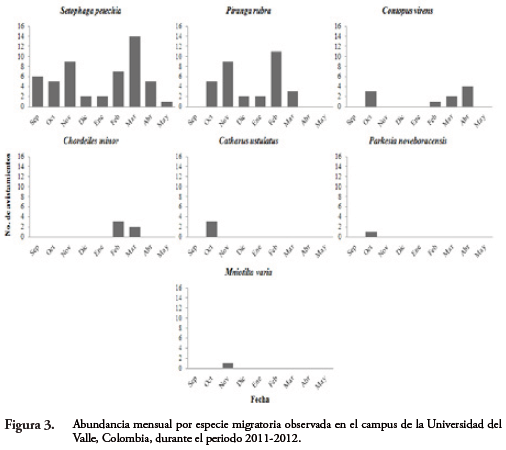
La Tabla 2 muestra las fechascorrespondientes a la primera y última observación de cada una de las especiesmigratorias.Lastres especies con mayor tiempo de permanencia en el campus fueron: S. petechia,P. rubra y C. virens, en contraste C. americanus, M. varia, C. ustulatus, P. noveboracensis, P. citrea y S. fusca se observaron una sola vez durante todo el periodo.
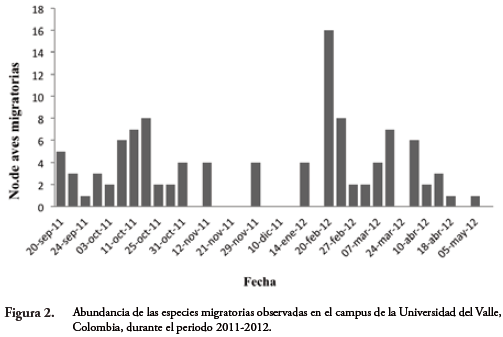
La especie más abundante registrada en los recorridos durante todo el periodo demuestreo fue S. petechia (n=51), seguida de P. rubra (n=34), la variación en las abundancias totales de especies migratorias por jornada de observación a través de la temporada de migración se muestra en la Figura 2. También se presenta la variación en la abundancia por especie durante la temporada (Fig. 3).
Se logró recapturar dos individuos de diferente especie: (1) S. petechia, capturada el día 18 de octubre de 2011 y recapturada el día seis de diciembre de 2011; y (2) P.citrea, capturada el día 17 de noviembre de 2011 y recapturada el día 22 del mismo mes y año. Para la información morfométrica de los individuos capturados ver CRUZ-BERNATE et al. (2014).
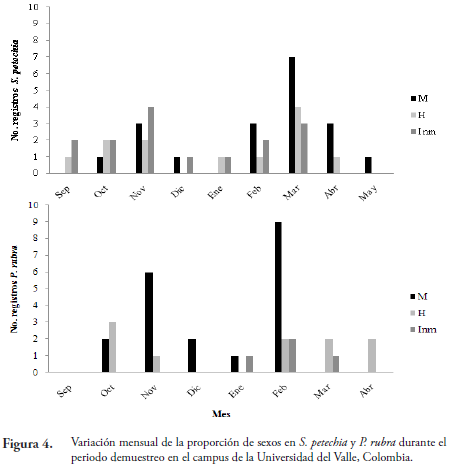
Las especies con dimorfismo sexual presentes en el campus fueron: S. petechia, P. rubra y M. varia, pero aquellas con información suficiente para analizar las frecuencias relativas por sexo fueron las dos primeras, donde se observaron diferencias en los patrones de llegada para cada sexo (Fig. 4).
Depósitos de grasa
No se encontró correlación entre el peso y el tiempo de permanencia en el sitio de invernada en S. petechia (ro0.5 (2) 43=0,469; P= 0.091).
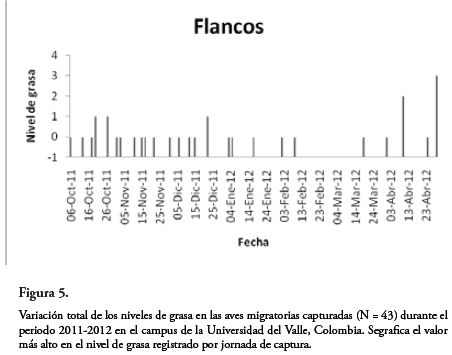
Los niveles de grasa en la fúrcula, abdomen y flancos permanecieron relativamente estables durante gran parte de la temporada y tan solo en los meses próximos a la partida de las aves (marzo y abril) se observó un marcado aumento (Figura 5). Se presenta también la variación en los niveles de grasa por especie a través de la temporada de invernada (Fig. 6).
Interacciones inter e intraespecíficas
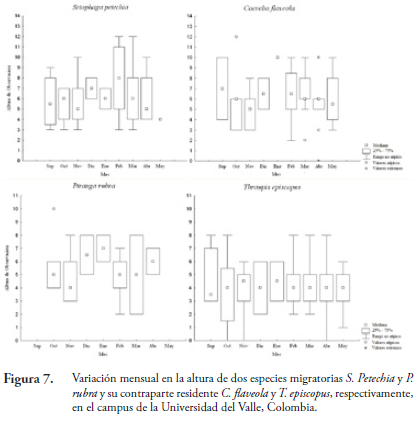
Durante las 42 jornadas de observación se registró dos interacciones agresivas de especies residentes hacia migratorias. S. petechia fue expulsada de un Chiminango (P. dulce) por T. episcopus con vuelo de suplante a 3 metros de altura y P. rubra fue perseguida en vuelo durante seis segundos por T. episcopus muy cerca de un Ficus (F. elastica). Al analizar la variación mensual en las alturas de forrajeo de dos especies migratorias frente a sus contrapartes residentes, se observa una ligera exclusión en la altura de forrajeo de las aves residentes hacia las migratorias, pero sus alturas se solapan en la mayoría de los meses (Figura 7 y 8).
DISCUSIÓN
Permanencia, abundancia y sexo
La riqueza de especies migratorias en el campus universitario es similar a la registrada por MUÑOZ et al. (2007) y REYES-GUTIÉRREZ et al. (2002), con 13 y 21 especies, respectivamente, donde se tienen en comúnocho de ellas. El mayor número presentado en el segundo estudio obedece al registro acumulado de especies durante cinco años, por lo que esta escala temporal demuestra la dinámica que puede tener la ecología de la migración año tras año. A diferencia de lo registrado por MUÑOZ et al. (2007), este estudio evidencia la presencia de cinco especies migratorias adicionales que utilizan el campus universitario como hábitat dentro de su ruta de migración. Estas especies son C. ustulatus, V. peregrina, S. castanea, P. noveboracensis y Empidonax virescens.
La presencia de P. citrea y S. castanea en el campus, evidencia el uso y la importancia de áreas con vegetación densa en zonas urbanas dentro de las ruta de migración. Estas dos especies han sido consideradas como prioritarias para la conservación dado que sus poblaciones han bajado constantemente desde los últimos 40 años en Colombia (BAYLY et al., 2011) y se conoce que llegan hasta Ecuador y Perú en su viaje de migración (RAPPOLE, 1995; DEL HOYO et al., 2010; OCAMPO-PEÑUELA, 2010; NARANJO et al., 2012).
S. petechia la especie migratoria más abundante durante el estudio, según DEL HOYO et al. (2010) es la más ampliamente distribuida de los parúlidosy en invierno habitan hábitats ligeramente boscosos o enmalezados (incluyendo los parques y jardines), zonas pantanosas y ribereñas, usualmente, debajo de los 500 m, pero ocasionalmente hasta 2600 m en Colombia. Su abundancia en este estudio, por tanto, obedece a que el campus ofrece condiciones de hábitat requeridos por la especie.
Los picos en el número de migrantes en el campus fueron similares a los observados por BAYLYet al. (2011) en El Darién, donde el máximo se dio a mediados de octubre ya finales de abril, así como en la Reserva El Dorado en la Sierra Nevada de Santa Marta (GÓMEZ et al., 2011), donde se registró el pico de migrantes a comienzos de abril.
Especies como C. americanus, M. varia, C. ustulatus, P. noveboracensis, P. citreay S. fusca, se observaron muy pocas veces al inicio de la temporada de migración, posiblemente, debido a que el campus se encuentre en un extremo de su franja de migración (e.g. región pacífica) (RAPPOLE, 1995; DEL HOYO et al., 2010; OCAMPO-PEÑUELA, 2010;NARANJO et al., 2012); al final del periodo de invernada, ninguna se observó de nuevo, probablemente por el uso de otra ruta para retornar a sus territorios reproductivos (NARANJO et al., 2012).
Las variaciones en las densidades por sexo en P. rubra durante la segunda mitad del periodo de invernada, posiblemente, se deban al uso de otra ruta en su migración diferencial al momento de regresar a sus sitios de reproducción (RAPPOLE, 1995). Según OTAHAL (1995) existen diferentes presiones selectivas que actúan sobre machos y hembras en gran cantidad de especies migratorias (migración diferencial).Los tiempos de migración y llegada a los territorios de reproducción pueden variar y conferir ventajas según el sexo. Mayor disponibilidad de recursos y posibilidad de escoger mejores territorios para los machos y condiciones óptimas de anidamiento para hembras (OTAHAL, 1995).
STOUFFER & DWYER (2003) determinaron la existencia de migración diferencial en Catharus guttatus, encontrando que las hembras parten primero que los machos en la migración de otoño debido a que no pueden competir con ellos por recursos en los sitios de invernada. También FRANCIS & COOKE (1986) encontraron que en algunas especies de parúlidoslos inmaduros de un año migran primero que los machos y las hembras. Lo anterior, se observó en S. petechia donde las hembras e inmaduros son los primeros en llegar a los sitios de invernada durante la migración de otoño (septiembre) y también los primeros en partir, los machos llegaron y partieron de regreso un mes después que el resto de los individuos.
Para Setophaga fusca en un estudio de la fenología de migración durante la primavera en cercanías de la Sierra Nevada de Santa Marta se encontró que los machos migraron primero a sus territorios de invernada que las hembras (GÓMEZ et al., 2011), contrario a lo observado en S. petechia. A pesar de que ambas especies pertenecen al mismo género no presentan patrones similares en su migración.
Depósitos de grasa
Desde el inicio de la migración de otoño, los niveles de grasa reportados para las aves capturadas no sobrepasaron el nivel 1 en las diferentes ubicaciones posibles y en su gran mayoríalas aves no presentaron depósitos energéticos. Probablemente, porque en los meses de septiembre a noviembre estas aves están cerca de su destino final, por lo que ya han utilizado la energía acumulada, por tanto, sus niveles de grasa son bajos o nulos (DEL HOYO et al., 2010; EBIRD,2013). Se espera que durante el periodo de invernada el abastecimiento energético conseguido en el hábitat supla las necesidades mínimas para mantener al individuo en buenas condiciones sin necesidad de acumular tejido adiposo (RAPPOLE, 1995). Como se observó en un individuo marcado de la especie S. petechia, que fue recapturado 50 días después en la misma zona, sus lugares de almacenamiento lipídico mostraron similares condicionesen los dos momentos de captura.
No se encontró correlación entre el peso y el tiempo transcurrido en los territorios de invierno, probablemente, por la naturaleza misma de la hiperfagia.Este es un fenómeno característico de especies migratorias quese produce en respuesta a diferentes estímulos tanto exógenos (e.g. horas de luz por día) como endógenos, justo antes de la migración (RAPPOLE, 1995; OCAMPO-PEÑUELA, 2010). La hormona prolactina cumple un papel fundamental en inducir la hiperfagia, la lipogenia (acumulación de grasa) y la inquietud migratoria (BERTHOLD, 1996). Sin embargo, como consecuencia de esto, el aumento de peso no es gradual mes a mes, sino que se centra en los meses previos al regreso hacia sus territorios de reproducción, entre marzo y abril (NARANJO et al., 2012).
Al final de la temporada de migración, durante los meses de marzo y abril, las aves presentaron en su mayoría reservas de grasa moderadas y de acuerdo con BAYLYet al. (2011) pueden continuar su migración hacia los territorios de reproducción.
Interacción aves migratorias vs. residentes
El nicho de los individuos migrantes está condicionado por el nicho de su contraparte ecológica residente, las aves migratorias presentan un alto grado de plasticidad ecológica en comparación con especies residentes y están restringidas a hábitats marginales (RAPPOLE, 1995). Se considera que el rol de las especies migratorias en los territorios de invernadaes de especies "fugitivas", que son capaces de sobrevivir por su relativamente rápida y temporal ocupación de nuevos hábitats adecuados tan pronto estos estén disponibles (RAPPOLE, 1995).
La variación mensual en las alturas de T. episcopus y de P. rubra muestran un solapamiento considerable en los meses de octubre, noviembre, febrero y marzo, siendo la variación de P. rubra mayor que la observada en T. episcopus. En cuanto a la mediana en la altura mensual, T. episcopus varió muy poco a través de los meses, posiblemente, efecto de su condición de ave residente, por el contrario, los valores de esta misma medida para P. rubra varían considerablemente a través de los meses y aunque hubo cierto solapamiento, la mediana de ambas nunca coincidieron.
Contrario a la visión tradicional de que las especies migratorias son simplemente visitantes estacionales de los trópicos, se plantea que las comunidades invernales en los trópicos están bien integradasy que cuando las especies migratorias regresan a sus territorios de nidada, las especies residentes no expanden sus nichos (GOCHFELD, 1985).Los resultados obtenidos podrían deberse a que P. rubra no genera interferencia que modifique la altura de forrajeo de T. episcopus a través de la temporada invernal, ya que, según GOCHFELD (1985), no son excluyentes. La mayor variación mensual observada para P. rubra, posiblemente, es un efecto del número de individuos registrados.
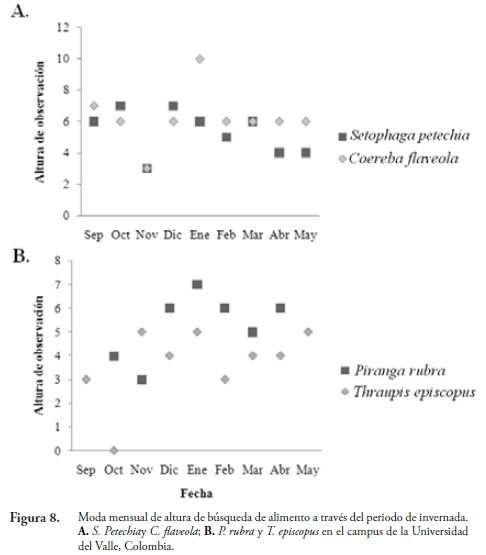
Para S. petechia y C. flaveola hay un solapamiento mayor en todos los meses de muestreo excepto en el mes de enero y mayo debido a los pocos registros de S. petechia con dos y un avistamiento, respectivamente. Probablemente, porque los hábitos de alimentación de C. flaveola, primariamente nectarívoro y en menor medida insectívoro (LIKOFF, 1986), no interfieren con los hábitos alimenticios de S. petechia, lo que permite que ambas especies compartan alturas similares y no se excluyan mutuamente.
Al comparar la moda mensual en los registros de altura de las especies anteriores, se encontró que solo en los dos picos de lluvia anual (noviembre y marzo) S. petechiay C. flaveola presentan el mismo valor de la moda (6 m). Probablemente, la mayor disponibilidad de artrópodos por el aumento en las lluvias permite que ambas especies usen el mismo estrato. En contraste, la moda mensual para P. rubra y T. episcopus nunca coincidió en el periodo de invernada.
A pesar de no haber observado una interferencia marcada en las alturas de forrajeo de las aves residentes por parte de las migratorias, RAPPOLE (1995) indica que este tipo de interacción migratoria-residente existe, pero estas interacciones resultan siendo bastante complejas.
Aún existen vacíos en el conocimiento ecológico de la comunidad de aves migratorias neárticas que utilizan gran parte de territorio colombiano como parada de abastecimiento o como destino final de su viaje, por lo que continuar realizando estudios enfocados específicamente en este tipo de aves debería ser una prioridad debido a su dependencia de conservación con estas áreas. Es necesario realizar estudios específicos para cada especie de ave migratoria, con la intención de generar estrategias específicas de conservación que permitan seguir manteniendo las poblaciones.
AGRADECIMIENTOS
Agradecemos a la Vicerrectoría de Investigaciones y al Departamento de Biología de la Universidad del Valle, en su convocatoria interna 01 de 2011, por la financiación del proyecto. A Carlos A. Ardila, Adriana Téllez y Vanessa Ardila por su continuo apoyo durante este proceso. A Adriana del Pilar Caicedo, Paola Montoya y a Óscar Hernández por su ayuda en campo. A Mónica Rodríguez y a Pamela Carvajal, quienes aportaron comentarios valiosos al manuscrito. A los jurados por sus valiosos aportes al manuscrito.
BIBLIOGRAFÍA
ALCALDÍA DE SANTIAGO DE CALI, DEPARTAMENTO ADMINISTRATIVO DE GESTIÓN DEL MEDIO AMBIENTE -DAGMA-,2010.-Las aves de mi ciudad: una guía de las aves de Cali. DAGMA, Cali, Colombia. [ Links ]
ANGARITA,I.,2002.-Composición y estructura de la avifauna de la ciudad de Cali: Tesis de grado, Universidad del Valle, Cali. [ Links ]
BAYLY, N.J., GÓMEZ, C. & CÁRDENAS,L.C., 2011.- Darién, Monitoreo de la migración durante el 2011 en la Reserva Natural de la Sociedad Civil Tacarcuna. Proyecto ando el Caribe. SELVA: Investigación para la conservación en el Neotrópico, Bogotá. Informe Técnico No.CEC06. [ Links ]
BERTHOLD, P., 1996.-Control of bird migration.Springer. [ Links ]
CRUZ-BERNATE, L., ARDILA-TÉLLEZ, J.D. & CAICEDO-ARGÜELLES, A.P.,2013.-Monitoreo y morfometría de la avifauna del campus de la Universidad del Valle 2011-2012. Biota Colombiana, 15 (1): 126-132.Recuperado de http://ipt.sibcolombia.net/valle/resource.do?r=avifauna_migratoria. [ Links ]
DEL HOYO, J., ELLIOT, A. &CHRISTIE, D.A., 2010.- Handbook of the birds of the world. Vol. 15. Weavers to New World Warblers. Lynx Editions, Barcelona. [ Links ]
ESPINAL, S., 1968.-Visión ecológica del departamento del Valle del Cauca. Universidad del Valle, Corporación Autónoma Regional del Valle (CVC) y Ministerio de Agricultura. Cali, Colombia. [ Links ]
FRANCIS, C.M. &COOKE, F.,1986.-Differential timing of spring migration in wood warblers (Parulinae). Auk, 103:548-556. [ Links ]
FINCH, D.M., 1991.- Population ecology, habitat requirements, and conservation of neotropical migratory birds. USDA Forest Service, Rocky Mountain Forest and Range Experiment Station. General Technical Report 205. [ Links ]
FUNDACIÓN PROAVES, 2009.- Plan para la conservación de las aves migratorias en Colombia. Conservación Colombiana, 11: 1-154. [ Links ]
GOCHFELD, M., 1985.-Numerical Relationships between Migrant and Resident Bird Species in Jamaican Woodlands. Ornithological Monographs, Neotropical Ornithology, 36: 654-662. [ Links ]
GÓMEZ, C., BAYLY, N.J., GONZÁLEZ, A.M., ABRIL, E., ARANGO, C., GIRALDO, J.I., SÁNCHEZ-CLAVIJO, L.M., BOTERO, J.E., CÁRDENAS, L., ESPINOSA, R., HOBSON, K., JAHN, A.E., JOHNSTON, R., LEVEY, D., MONROY, A., ANDNARANJO, L.G., 2010.- Avances en la investigación sobre aves migratorias neárticas-neotropicales en Colombia y retos para el futuro: trabajos del III Congreso de Ornitología Colombiana. Revista Ornitología Colombiana, 11: 3-13. [ Links ]
GÓMEZ, C. & BAYLY, N.J., 2011.- La migración de aves en la Reserva El Dorado, Sierra Nevada de Santa Marta, Colombia. Proyecto Cruzando el Caribe: Identificación de sitios de parada críticos para aves migratorias Neotropicales en el norte de Colombia. SELVA: Investigación para la Conservación en el Neotrópico, Bogotá. Informe técnico No. CEC05. [ Links ]
HEDENSTRÖM, A., 2010.-Extreme Endurance Migration: What Is the Limit to Non-Stop Flight? PLoSBiol., 8(5): e1000362. [ Links ]
HILTY, S.L. & BROWN, W.L., 2001.- Guía de las aves de Colombia. SAO, Universidad del Valle y American Bird Conservancy -ABC-. Colombia. [ Links ]
JOSENS, M.L., FAVERO, M. & ESCALANTE, A.H., 2012.-Diversity, seasonality and structure of birds assemblages associated with three wetlands in the southeastern Pampas, Argentina. Ardeola, 59 (1): 93-109. [ Links ]
LIKOFF, L.E., 1986.-The Encyclopedia of birds, Volume 1. Infobase Publishing, New York. [ Links ]
MACGREGOR-FORS, I. & ORTEGA-ÁLVAREZ, R., 2013.- Ecología urbana: experiencias en América Latina. Instituto de Ecología, A. C. [ Links ]
MORENO, M.I., 2009.- Aves migratorias en Colombia. Revista Conservación Colombiana, 11: 9-26. [ Links ]
MUÑOZ, M.C., FIERRO-CALDERÓN, K. & RIVERA-GUTIERREZ, H.F., 2007.- Las aves del Campus de la Universidad del Valle, una isla verde urbana en Cali, Colombia. Ornitología Colombiana, 5(5): 5-20. [ Links ]
NARANJO, L.G., AMAYA, J.D., EUSSE-GONZÁLEZ, D. & CIFUENTES-SARMIENTO, Y., 2012.-Guía de las Especies Migratorias de la Biodiversidad en Colombia. Aves. Vol. 1. Ministerio de Ambiente y Desarrollo Sostenible/WWF Colombia. Bogotá, Colombia. [ Links ]
OTAHAL, C.D., 1995.- Sexual Differences in Wilson's Warbler Migration. Journal of Field Ornithology, 66 (1): 60-69. [ Links ]
OCAMPO-PEÑUELA, N., 2010.- El fenómeno de la migración en aves: una mirada desde la Orinoquia. Orinoquia, 14 (2): 188-200. [ Links ]
OREJUELA, J.E.,RAITT, R.J. & ÁLVAREZ-LÓPEZ, H.,1980.-Differential use by North American migrants of three types of Colombian forest:253-264 (en) KEAST, A. & MORTON, E.S. (ed.). Migrant Birds in the Neotropics: Ecology, Behavior, Distribution, and Conservation. Smithsonian Institution Press, Washington. [ Links ]
ORTEGA-ÁLVAREZ, R., MACGREGOR-FORS, I., 2011.-Dusting off the file: A review of knowledge on urban ornithology in Latin America. Landscape and Urban Planning, 101: 1-10. [ Links ]
RAPPOLE, J.H. &MCDONALD, M.V., 1994.- Cause and effect in populations declines of migratory birds. Auk, 111:652-660. [ Links ]
RAPPOLE,J.H., 1995.-The ecology migrant birds. A Neotropical perspective Smithsonian Institution Press, Washington. [ Links ]
REYES-GUTIERREZ,M., SEDANO, R. & DURÁN, S., 2002.- Lista anotada de la avifauna de la Universidad del Valle, Cali, Colombia. Boletín SAO, 13: 12-25. [ Links ]
RIVERA-GUTIERREZ, H.F., 2002.- Aspectos ecológicos de una comunidad de aves en una mancha de bosque del áreas uburbana de la ciudad de Cali: Tesis de grado, Universidad del Valle, Cali. [ Links ]
ROBBINS, C.S., BRUUN, B. & ZIM, H.S., 2001.-Birds of North America: A Guide to Field Identification. Golden press, New York. [ Links ]
STOUFFER, P.C. &DWYER, G.M., 2003.-Sex-Biased Winter Distribution and Timing of Migration of Hermit Thrushes (Catharus guttatus) in Eastern North America. Auk, 120 (3): 836-847. [ Links ]
THE NORTH AMERICAN BANDING COUNCIL, 2001.- Guía de estudio del anillador de Norteamérica. California. [ Links ]
ZAR, J.H., 2010.-Biostatistical analysis, 5d. Prentice Hall, New Yersey. [ Links ]
