










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
versión impresa ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.20 no.1 Manizales ene./jun. 2016
https://doi.org/10.17151/bccm.2016.20.1.10
DOI: 10.17151/bccm.2016.20.1.10
FIJACIONES DE LOS MÚSCULOS CAUDO-MEDIALES DEL ANTEBRAZO DE PRIMATES: UNA REVISIÓN Y ADAPTACIÓN A LA TERMINOLOGÍA ANATÓMICA VETERINARIA*
PRIMATES' CAUDOMEDIAL FOREARM MUSCLE FIXATION: A REVIEW AND ADAPTATION TO VETERINARY ANATOMICAL TERMINOLOGY
* FR. 20-III-2015 FA. 7-IV-2016
1 Departamento de Sanidad Animal, Facultad de Medicina Veterinaria y Zootecnia, Universidad del Tolima, Ibagué, Colombia.
2 Grupo de investigación en Medicina y Cirugía de Pequeños Animales, Universidad del Tolima, Ibagué, Colombia.
3 Docente Cátedra Facultad de Ciencias Pecuarias, Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. Bogotá, Colombia.
CÓMO CITAR:
VÉLEZ-GARCIA, J.F. & CATAÑEDA, F.E., 2016.- Fijaciones de l,os músculos caudio-mediales del antebrazo en primates: una revisión y adaptación a la terminología anatómica veterinaria. Bol. Cient. Mus. Hist. Nat. U. de Caldas, 20 (1): 126-140. DOI : 10.17151/bccm.2016.20.1.10
Resumen
Los músculos caudo-mediales del antebrazo de primates presentan sus fijaciones en los huesos del brazo, antebrazo y mano, para realizar funciones apropiadas para su patrón de locomoción, al igual que, pueden presentar semejanzas y variantes intraespecíficas e interespecíficas, las cuales nos permiten encontrar que en algún momento compartieron un ancestro en común; y por otro lado, el cirujano veterinario las podrá tener en cuenta al realizar un abordaje quirúrgico que involucre la cara disto-medial del brazo y la cara caudo-medial del antebrazo; por lo tanto, el presente documento revisa los orígenes e inserciones de los músculos caudo-mediales del antebrazo de primates, presentando un análisis comparativo entre distintas especies de primates. Palabras clave: Antebraquial, inserción, miología, origen.
Abstract
The caudal-medial forearm muscles of primates present their attachments in the arm, forearm and hand bones to carry out suitable functions for their locomotion pattern, as well as they may present intraspecific and interspecific similarities and variationswhich allow finding that at some point they shared a common ancestor.On the other hand, the veterinary surgeon may take them into account when making a surgical approach that involves the distal-medial face of the arm and the caudal-medial face of the forearm.Therefore, this paper reviews the origins and insertions of forearm caudal-medial muscles, presenting a comparative analysis between different species of primates.
Key words: Antebrachial, insertion, myology, origin.
INTRODUCCIÓN
En primates se han descrito diversas formas de locomoción, de las cuales, la mayoría las realizan mediante el apoyo de los cuatro miembros locomotores, a excepción del desplazamiento braceador como lo tienen los gibones (Hylobates sp. y Symphalangus sp.); y la bípeda en el caso del humano (ANKEL-SIMONS, 2007). Para su desplazamiento, los músculos permiten la movilidad de las palancas formadas por los huesos y articulaciones (KARDONG, 2011), y estos movimientos permiten al animal desenvolverse en su ambiente, bien sea para adquirir sus alimentos, defenderse y huir de potenciales predadores, por lo tanto, su sistema músculo-esquelético se ha adaptado para realizar diferentes tipos de movimientos esenciales para cada especie de primate (AVERSI-FERREIRA et al., 2005a, 2006). El sistema musculoesquelético se ha desarrollado a partir del mesodermo, del cual, se forman los huesos, articulaciones y músculos de los miembros y paredes corporales (LANGMAN, 2007; SINOWATZ, 2010); entre estos, se encuentran los músculos del antebrazo, los cuales, presentan diversas fijaciones en el húmero, radio, ulna y en los huesos de la mano, para permitir diversos movimientos en las articulaciones del antebrazo, carpo y dedos; para ello se dividen en dos grupos, que se clasifican por su posición anatómica como cráneo-laterales y caudo-mediales, donde los cráneo-laterales participan en la extensión de carpo y dedos, flexión de codo y supinación; y los caudo-mediales en la flexión de carpo y dedos, y pronación (DYCE et al., 2012; EVANS & DE LAHUNTA, 2013; STANDRING, 2008).
El conocimiento de las características morfológicas de los músculos de los primates explicarán las diversas adaptaciones entre especies (AVERSI-F et al., 2005b); y además tienen un significado clínico múltiple (CRIBILLERO et al., 2009), ya que permitirán al médico veterinario implementar mejores tratamientos quirúrgicos y ortopédicos en primates no humanos que lo requieran, por haber sufrido un trauma producto de agresiones grupales, accidentes durante la captura o por la presentación de tumores localizados; por lo tanto, el presente estudio tiene como objetivo comparar el origen e inserción de los músculos caudo-mediales del antebrazo de primates.
MATERIALES Y MÉTODOS
Se utilizaron las descripciones anatómicas de origen e inserción de los músculos caudo-mediales del antebrazo de primates reportados en la literatura, y se adaptaron a los términos que indican situación y dirección en el miembro torácico de la Nómina Anatómica Veterinaria (Nomina Anatomica Veterinaria) (ICVGAN, 2012), describiendo las diferencias entre primates, incluyendo al humano, que aunque es una especie bípeda, para respetar términos comparativos, también fue adaptado como si fuera cuadrúpedo, por lo tanto, sus músculos anteriores del antebrazo fueron incluidos como caudo-mediales, y fueron divididos en un grupo superficial y uno profundo, resumidos en la tabla 1 y 2 respectivamente.
RESULTADOS
Músculos superficiales de la parte caudo-medial del antebrazo
El músculo pronador terete (Pronator teres) se origina en el epicóndilo medial del húmero y se inserta en el margen medial del tercio medio del radio en el mico de Goeldi -Callimico goeldii-(HILL, 1959), macaco cangrejero -Macaca fascicularis- (KIMURA & TAKAI, 1970), mono maicero -Sapajus apella-, mono silbador -Sapajus libidinosus- (AVERSI-F et al., 2006, 2011), capuchino de frente blanca -Cebus albifrons- (CRIBILLERO et al., 2009), gorila (Gorilla gorilla), en gibones como el gibón de manos blancas -Hylobates lar-, el gibón crestado -H. pileatus-, el gibón plateado -H. moloch-, y el siamang –Symphalangus syndactylus-(HEPBURN, 1892; MICHILSENS et al., 2009), y en papiones como Papio anubis (CHAMPNEYS, 1871) y Papio hamadryas (ACKERMANN, 2003). En el gálago del Senegal -Galago senegalensis- se origina además en el septo entre el bíceps braquial y tríceps braquial (STEVENS et al., 1977). En el mono Rhesus -Macaca mulatta-, el orangután (Pongo pygmaeus), el chimpancé (Pan troglodytes) y el humano (Homo sapiens) se presenta otra cabeza diferente a la humeral, que se origina en el proceso coronoides de la ulna (ACKERMANN, 2003; CHAMPNEYS, 1871; DRAKE et al., 2005; HEPBURN, 1892), aunque esta puede estar ausente en H. sapiens (STANDRING, 2008), P. pygmaeus, G. gorilla, y P. troglodytes (TESTUT & LATARJET, 1984). En cuanto su inserción, se presenta en la parte media del margen medial del radio en el H. sapiens (STANDRING, 2008), y más distal en P. troglodytes y P. pygmaeus (ACKERMANN, 2003; CHAMPNEYS, 1871; HEPBURN, 1892), aunque PRIMROSE (1900) en este último lo encuentra insertado en el tercio proximal del radio, similar a lo encontrado en
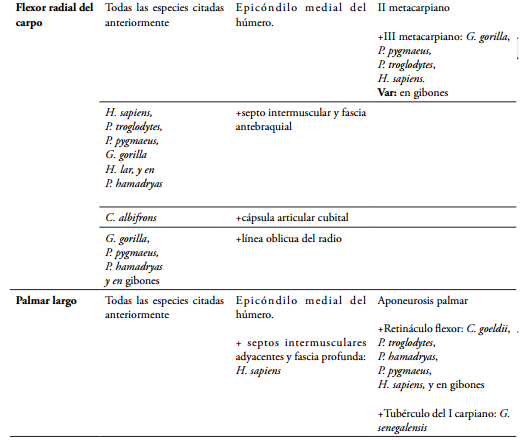
El músculo flexor radial del carpo (Flexor carpi radialis) se origina en el epicóndilo medial del húmero, y se inserta en la base del segundo metacarpiano en C. goeldii (HILL, 1959), M. fascicularis, M. mulatta (ACKERMANN, 2003; KIMURA & TAKAI, 1970), G. senegalensis (STEVENS et al., 1977), en gibones (MICHILSENS et al., 2009), S. apella, S. libidinosus (AVERSI-F et al., 2006, 2011), en el género Lemur y Propithecus (DIOGO et al., 2012). En otras especies se origina además en el septo intermuscular con el pronador terete y fascia antebraquial, como ocurre en H. sapiens (STANDRING, 2008; TESTUT & LATARJET, 1984), P. troglodytes, P. hamadryas (CHAMPNEYS, 1871), G. gorilla, P. pygmaeus, y se ha encontrado como una variante en H. lar (MICHILSENS et al., 2009), la cual ya se había descrito en un gibón sin especificar (HEPBURN,1892). En C. albifrons se puede encontrar originado además desde la cápsula articular cubital (CRIBILLERO et al., 2009). En otros se puede presentar un origen de más, en la línea oblicua del radio inmediatamente al lado de la inserción del pronador terete, como sucede en G. gorilla, P. pygmaeus (HEPBURN, 1892; PRIMROSE, 1900), P. hamadryas (CHAMPNEYS, 1871) y en gibones (HEPBURN, 1892; DIOGO et al., 2012), aunque MICHILSENS et al. (2009), lo reportan originado exclusivamente del húmero, siendo el origen radial una variante ocasional en H. sapiens (PRIMROSE, 1900). El músculo presenta doble inserción en la parte palmar de la base del segundo y tercer metacarpiano en Haplorrinos (DIOGO et al., 2012) como G. gorilla, P. pygmaeus, y P. troglodytes (CHAMPNEYS, 1871; HEPBURN, 1892), al igual que lo reportan en H. sapiens (DRAKE et al., 2005; STANDRING, 2008), mientras TESTUT & LATARJET (1984) lo reportan como una variedad y lo encuentran como patrón común insertado únicamente en el II metacarpiano, al igual que lo reporta MICHILSENS et al. (2009) en gibones, y PRIMROSE (1900) en P. pygmaeus, aunque HEPBURN (1892) encuentra en un gibón la doble inserción en el II y III metacarpiano. Este músculo puede llegar a estar ausente en H. sapiens (STANDRING, 2008).
El músculo palmar largo (Palmaris longus) se origina en el epicóndilo medial del húmero, y su tendón se inserta en la aponeurosis palmar en S. apella, S. libidinosus (AVERSI-F et al., 2006, 2011), C. albifrons (CRIBILLERO et al., 2009), C. olivaceus, A. paniscus, A. seniculus (YOULATUS, 2000), M. fascicularis, M. mulatta (ACKERMANN, 2003; KIMURA & TAKAI, 1970), y en gibones (HEPBURN, 1892; MICHILSENS et al., 2009). En H. sapiens se origina además en los septos intermusculares adyacentes y fascia profunda (STANDRING, 2008). Se inserta además en el retináculo flexor en C. goeldii (HILL, 1959), P. troglodytes, P. hamadryas, P. pygmaeus, H. sapiens, y en gibones (ACKERMANN, 2003; CHAMPNEYS, 1871; HEPBURN, 1892; PRIMROSE, 1900; STANDRING, 2008). En G. senegalensis se alcanza a insertar en la base del primer metacarpiano a través del tubérculo del primer carpiano (trapecio) (STEVENS et al., 1977). En H. sapiens puede estar ausente en uno o ambos antebrazos (DRAKE et al., 2005; STANDRING, 2008), al igual que, en G. gorilla y en P. troglodytes (DIOGO et al., 2012; TESTUT & LATARJET, 1984); o llegar en H. sapiens a presentar dos palmares largos (PAI et al., 2008).
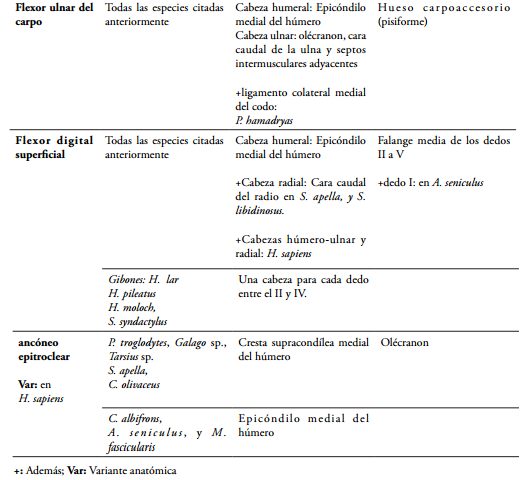
El músculo flexor ulnar del carpo (Flexor carpi ulnaris) presenta dos cabezas, una humeral que se origina en el epicóndilo medial del húmero; y una ulnar, que se origina en el olécranon, en los dos tercios proximales del margen caudal de la ulna y septos intermusculares adyacentes; y se insertan en el hueso carpoaccesorio (pisiforme) en S. apella, S. libidinosus (AVERSI-F et al., 2006, 2011), C. albifrons (CRIBILLERO et al., 2009), M. fascicularis (KIMURA & TAKAI, 1970), G. senegalensis (STEVENS et al. 1977), en gibones (MICHILSENS et al., 2009), P. pygmaeus, P. troglodytes, G. gorilla y H. sapiens (DRAKE et al., 2005; HEPBURN, 1892; PRIMROSE, 1900; STANDRING, 2008). En el P. hamadryas la cabeza humeral también se origina desde el ligamento colateral medial del codo (CHAMPNEYS, 1871); y en el género Ateles, el origen ulnar se presenta a lo largo de casi todo el hueso (ACKERMANN, 2003), siendo similar a C. goeldii (HILL, 1959) y A. seniculus (YOULATUS, 2000); aunque YOULATUS (2000) lo representa gráficamente en A. paniscus, originado únicamente en los dos tercios proximales de la ulna.
El músculo flexor digital superficial (Flexor digitorum superficialis) en S. libidinosus y S. apella presenta dos cabezas de origen, una humeral en el epicóndilo medial del húmero, y una radial en la cara caudal del radio; y se inserta en la superficie palmar de la falange media del II al V dedo (AVERSI-F et al., 2006, 2011). Únicamente se presenta la cabeza humeral en C. albifrons (CRIBILLERO et al., 2009), M. fascicularis, M. mulatta (ACKERMANN, 2003; KIMURA & TAKAI, 1970), G. senegalensis (STEVENS et al. 1977) y en P. hamadryas (CHAMPNEYS, 1871). Los géneros Ateles y Alouatta poseen una cabeza superficial y una cabeza profunda que se originan en la superficie cráneo-medial y craneal del epicóndilo medial del húmero, respectivamente (YOULATUS, 2000). En C. goeldii es un músculo que se divide en dos porciones, entre las cuales, una se origina en el epicóndilo medial del húmero, y la otra se origina en la parte caudal del cóndilo humeral, profundo al origen del palmar largo (HILL, 1959). En H. sapiens presenta dos cabezas, una húmero-ulnar que se origina en el epicóndilo medial del húmero, y la zona adyacente del margen medial del proceso coronoides de la ulna; y una cabeza radial que se origina en la línea oblicua caudal del radio (DRAKE et al., 2005; STANDRING, 2008). Según CHAMPNEYS (1871), este músculo en el chimpancé, únicamente se diferencia del humano, en que sus tendones son diferenciados más proximalmente y son más largos. En P. pygmaeus se presentan unos orígenes similares, aunque se describe la cabeza ulnar por separado, originándose desde el proceso coronoides y el olécranon (ACKERMANN, 2003; PRIMROSE, 1900). En gibones se presentan cuatro orígenes distribuidos para los cuatro dedos, donde el que va para el II dedo se origina en la parte proximal de la ulna, el del III dedo en el margen medial del radio, el del IV dedo en el epicóndilo medial del húmero, y el del V dedo en la mitad proximal de la ulna (MICHILSENS et al., 2009), diferenciándose de lo hallado en un gibón (HEPBURN, 1892), en el cual se encuentra que el II y III dedo reciben tendones desde el origen radial. En S. syndactylus se puede encontrar como variante, que las cabezas para los dedos IV y V pueden estar fusionadas (MICHILSENS et al., 2009), siendo similar a lo encontrado en el humano, donde ésta se denomina cabeza húmero-ulnar (FICAT, 1998). En los primates no hominoides diseccionados por DIOGO et al. (2012), este músculo se origina exclusivamente desde el brazo, bien sea en el epicóndilo medial del húmero, tendón común, o cápsula articular del codo; y en los hominoides encuentra orígenes además en radio y ulna, corroborando lo descrito por TESTUT & LATARJET (1984), quienes describen que el origen radial es de regla en los antropoides, sin embargo en estos puede faltar en algunas ocasiones, como sucede a veces en H. sapiens (STANDRING, 2008). En la mayoría de los primates reportados, este músculo se inserta en la falange media de los dedos II a V, a excepción de A. seniculus, en el cual se encuentran tendones que aunque se insertan en la falange media, se dirigen para el I a IV dedo o hasta el V dedo (YOULATUS, 2000).
El músculo ancóneo epitroclear (Anconeus epitrochlearis) es un músculo que se ha encontrado en P. troglodytes, Galago sp., Tarsius sp. (HOWELL & STRAUS, 1932), S. apella, C. olivaceus (YOULATUS, 2000), en los cuales, se origina en la cresta supracondílea medial del húmero, y se inserta en el olécranon, diferenciándose de C. albifrons (CRIBILLERO et al., 2009), A. seniculus (YOULATUS, 2000), y M. fascicularis (KIMURA & TAKAI, 1970), donde se origina únicamente desde el epicóndilo medial del húmero, al igual que, cuando se presenta ocasionalmente en H. sapiens (BLAND-SUTTON, 1897; DIOGO & ABDALA, 2010). También se ha encontrado en primates de los géneros Lemur, Propithecus, Tarsius, Pithecia, Callithrix, Aotus, Saimiri, Cercopithecus, Colobus y Papio (DIOGO & WOOD, 2012).
Músculos profundos de la parte caudo-medial del antebrazo
El músculo flexor digital profundo (Flexor digitorum profundus) presenta diversos orígenes entre primates, como sucede en C. albifrons, en el cual, presenta tres cabezas, donde la cabeza humeral se origina en el epicóndilo medial del húmero; una radial en la superficie caudo-medial del radio y la membrana interósea; y una ulnar que se origina en la cara caudo-lateral de la ulna (CRIBILLERO et al., 2009), siendo similar a A. paniscus, aunque en este último la ulnar se origina además en la membrana interósea (YOULATUS, 2000). Únicamente se presentan las cabezas radial y ulnar en S. apella, C. olivaceus, A. seniculus (YOULATUS, 2000), M. mulatta, M. fascicularis, P. hamadryas, P. troglodytes, G. gorilla, y P. pygmaeus (ACKERMANN, 2003; CHAMPNEYS, 1871; HEPBURN, 1892; KIMURA & TAKAI, 1970; PRIMROSE, 1900). En el H. sapiens únicamente se presenta la cabeza ulnar, y se origina en los tres cuartos proximales de la superficie caudal y medial de la ulna, y región adyacente de la membrana interósea, y se puede encontrar como variante, fascículos originados en la cara caudal del radio a diferentes niveles, pero estos refuerzan al II dedo (STANDRING, 2008; TESTUT & LATARJET, 1984). AVERSI-F et al. (2005b, 2011) en S. apella y S. libidinosus, al igual que, CHAMPNEYS (1871) en P. troglodytes, describen este músculo, únicamente con la presencia de la cabeza ulnar. En el patrón común de gibones, se encuentra originado en los dos tercios medios de la superficie del radio, en la parte proximal de la ulna, y superficie caudal y margen medial de la ulna, y como variantes se puede encontrar originado en S. syndactylus, además desde el tercio medio del radio y membrana interósea, y en H. lar originado además en el epicóndilo medial del húmero (MICHILSENS et al., 2009), además HEPBURN (1892) en un gibón encuentra un segmento húmero-radial y uno ulnar, donde el primero se origina en el epicóndilo medial del húmero y en el radio, el cual, envía tendones para el I, II y III dedo; y el segundo envía tendones para el IV y V dedo. En S. syndactylus (MICHILSENS et al., 2009) y en H. gabriellae (DIOGO et al., 2012), se puede encontrar como variante que el vientre muscular para el I dedo va por separado, pero su tendón puede presentar conexiones con el tendón del II dedo. En S. syndactylus también se puede encontrar un vientre y un tendón individuales para el tercer dedo (MICHILSENS et al., 2009). En G. senegalensis se describe originado mediante cinco porciones; la porción humeral se fusiona con el pronador terete y el flexor radial del carpo, y se origina en la cresta supracondílea medial del húmero; la porción húmero-radial se origina en el epicóndilo medial del húmero y superficie caudal del radio; la porción radial se origina en los tres cuartos proximales caudo-mediales del radio y membrana interósea; la porción ulnar profunda se origina en la mitad distal de la ulna y membrana interósea; y la porción ulnar superficial se origina desde la superficie proximal de la ulna hasta casi el extremo distal (STEVENS et al., 1977). En C. goeldii se describen dos cabezas ulnares, una superficial que se origina desde el ligamento anular del radio, y desde el tercio proximal del cuerpo de la ulna; y una profunda que se origina en la cara caudal de la ulna en la parte distal de su cuerpo; también presenta dos cabezas humerales que se originan en el epicóndilo medial del húmero, y una radial que se origina en la cara caudal del radio y membrana interósea (HILL, 1959).
Los tendones del músculo flexor digital profundo se insertan en la parte palmar de la base de la falange distal en todos los primates, pero presentan diferentes aportes de los vientres musculares para los dedos. Por un lado, se fusionan antes de llegar al canal del carpo y se separan cuando pasan a través de él, para ir a cada dedo, como sucede en M. fascicularis (ACKERMANN, 2003; KIMURA & TAKAI, 1970), y G. senegalensis (STEVENS et al., 1977). En P. pygmaeus, P. troglodytes, y G.. gorilla la cabeza radial envía tendones para los dedos I a III, y la ulnar para el IV y V dedo (ACKERMANN, 2003; HEPBURN, 1892), aunque PRIMROSE (1900) en P. pygmaeus no encuentra tendón para el I dedo, similar a lo reportado por DIOGO et al. (2012), quienes afirman que este tendón en grandes simios es muy delgado, vestigial o puede estar ausente. En S. apella, S. libidinosus (AVERSI-F et al., 2005b, 2011) y P. troglodytes (CHAMPNEYS, 1871) envían tendones para el III, IV y V dedo. En A. paniscus la cabeza radial envía tendones para el II, III y IV dedo, y la cabeza ulnar para el IV y V dedo (YOULATUS, 2000), diferenciándose del Ateles diseccionado por ACKERMANN (2003), quien reporta que la cabeza radial envía tendón para el II dedo, y la ulnar envía del II al V dedo. MICHILSENS et al. (2009) en S. syndactylus encuentran una variación importante entre los vientres musculares y la distribución de tendones para los dedos, desde vientres musculares hasta tendones fusionados. En C. goeldii las cabezas humerales y la radial envían tendones para el I, II y III dedo; y las cabezas ulnares para el III, IV y V dedo (HILL, 1959). En A. paniscus la cabeza radial envía tendones para los dedos I a IV, y la cabeza ulnar para los dedos III a V (YOULATUS, 2000).
Dentro de los músculos flexores profundos de los dedos, en H. sapiens se encuentra de manera independiente el músculo flexor largo del I dedo (Flexor pollicis longus), el cual se origina en la cara caudal del radio y en la mitad adyacente de la membrana interósea (DRAKE et al., 2005; STANDRING, 2008). En gibones se puede encontrar este músculo como una variante (DIOGO et al., 2012; DIOGO & WOOD, 2012; HEPBURN, 1892; MICHILSENS et al., 2009), al igual que un origen desde el epicóndilo medial del húmero (MICHILSENS et al., 2009), como también puede ocurrir en H. sapiens; e inclusive en esta última especie se puede presentar un fascículo desde la ulna (STANDRING, 2008; TESTUT & LATARJET, 1984). En otras especies de primates, por homología de origen, describen un flexor largo del I dedo, pero este se diferencia del H. sapiens porque además de su origen en el radio también se origina en el epicóndilo medial del húmero, y además envía tendones para el I y II dedo como sucede en S. apella, S. libidinosus (AVERSI-F et al., 2005b; 2011) y P. troglodytes (CHAMPNEYS, 1871), por lo tanto, este debe ser comprendido como la cabeza radial del músculo flexor digital profundo, ya que se origina en la superficie caudal del radio y en el epicóndilo medial del húmero, como lo descrito por YOULATUS (2000) en S. apella y C. olivaceus, aunque en estos encuentra que la cabeza radial, además, también envía tendón para el III dedo, por lo tanto, no hay una masa muscular independiente para el I dedo, como sucede en el patrón común del H. sapiens (STANDRING, 2008; TESTUT & LATARJET, 1984).
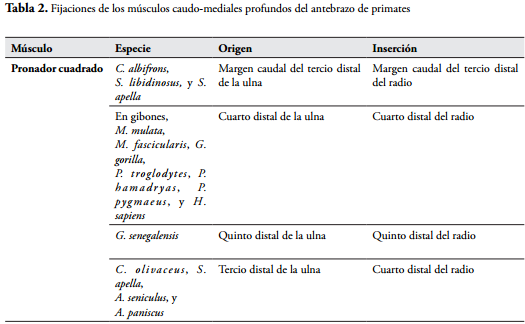
El pronador cuadrado (Pronator quadratus) se origina en el margen caudal del tercio distal de la ulna y se inserta en el margen caudal del tercio distal del radio en C. albifrons (CRIBILLERO et al., 2009), S. libidinosus y S. apella (AVERSI-F et al., 2005b, 2011). En otras especies se origina en el cuarto distal del margen caudal de la ulna, y se inserta en el cuarto distal de la cara y margen caudal del radio, como sucede en gibones, M. mulatta, M. fascicularis, G. gorilla, P. troglodytes, P. hamadryas, P. pygmaeus, y H. sapiens (ACKERMANN, 2003; CHAMPNEYS, 1871; DRAKE et al., 2005; HEPBURN, 1892; KIMURA & TAKAI, 1970; MICHILSENS et al., 2009; PRIMROSE, 1900; STANDRING, 2008; TESTUT & LATARJET, 1984). En G. senegalensis se origina a nivel del quinto distal de la cresta medial de la ulna y se inserta en la parte caudal del quinto distal del radio (STEVENS et al., 1977). En C. olivaceus, S. apella, A. seniculus, y A. paniscus se origina en el tercio distal de la cara caudal de la ulna, y se inserta en el cuarto distal de la cara caudal del radio (YOULATUS, 2000).


DISCUSIÓN
Las descripciones anatómicas de las fijaciones de los músculos son realizadas en diferentes primates con términos de dirección y situación que se utilizan en H. sapiens (ACKERMANN, 2003; CHAMPNEYS, 1871; HEPBURN, 1892; HILL, 1959; KIMURA & TAKAI, 1970; MICHILSENS et al., 2009; PRIMROSE, 1900; YOULATUS, 2000), siendo esta última, una especie cuyos términos han sido adaptados a su postura bípeda y con sus manos en supinación, por lo tanto, los músculos que permiten la flexión de carpo, dedos y pronación se ubican en la parte anterior del antebrazo (STANDRING, 2008; TESTUT & LATARJET, 1984); pero estos mismos términos no deberían ser usados en primates no humanos, ya que estos presentan tipos de locomoción que requieren el apoyo de sus manos en pronación o en semi-supinación para su desplazamiento terrestre, así como entre los troncos y ramas de los árboles (ANKEL-S, 2007); por lo tanto, sus músculos flexores se ubican caudales y mediales en el antebrazo, como se describe en mamíferos domésticos (DYCE et al., 2012; EVANS & DE LAHUNTA, 2013; ICVGAN, 2012). Por otro lado, CRIBILLERO et al. (2009) y STEVENS et al. (1977), en C. albifrons y en G. senegalensis respectivamente, realizan las descripciones respetando su posición cuadrúpeda, pero utilizan los términos anterior y posterior, los cuales, en la Nómina Anatómica Veterinaria, están restringidos para la descripción de estructuras en el bulbo ocular y en el oído interno (ICVGAN, 2012), por lo tanto, las descripciones anatómicas en primates no humanos deberían adaptarse a los términos sugeridos en la Nómina Anatómica Veterinaria, como se hizo en la presente revisión monográfica con las fijaciones de los músculos caudo-mediales del antebrazo reportadas por diferentes autores.
Una vez adaptados los términos, se puede observar que en la parte caudo-medial del antebrazo de las diferentes especies de primates, se encuentran músculos homólogos que van a permitir movimientos de pronación, flexión de carpo y de las articulaciones de los dedos de la mano. En los diferentes tipos de locomoción que hay en primates, la mano entra a ser un componente activo para el desplazamiento, y entre los movimientos, la pronación se puede llevar a cabo gracias a la presencia de dos músculos, entre ellos, el pronador terete, el cual se encuentra en el grupo superficial, y puede presentar entre una y dos cabezas de origen, así como diferencias en cuanto la distribución de su inserción, la cual, permite aplicar que especies con inserciones más proximales tengan movimientos más rápidos pero con menor fuerza (KARDONG, 2011), como sucede en Ateles, siendo un género de locomoción cuadrúpeda y braquiadora (DEFLER, 2010); y las inserciones más distales dan movimientos más lentos pero con mayor fuerza (KARDONG, 2011), como sucede en P. troglodytes y P. pygmaeus, siendo especies que se desplazan de manera cuadrúpeda (ANKEL-S, 2010). Otro músculo pronador se ubica en el grupo profundo, y es el pronador cuadrado que presenta diferencias interespecíficas en cuanto su distribución entre la ulna y el radio, pudiéndose encontrar fijado en estos, desde un tercio distal como sucede en la mayoría de primates descritos o hasta un quinto distal como ocurre en G. senegalensis (STEVENS et al., 1977).
Entre los músculos flexores del carpo se pueden encontrar el flexor radial del carpo, el flexor ulnar del carpo y el palmar largo, músculos que no presentan casi variaciones entre especies de primates, aunque en Ateles y C. goeldii el origen del flexor ulnar del carpo se extiende casi a lo largo de la ulna (HILL, 1959; ACKERMANN, 2003), lo que le da una mayor extensión a su origen, por lo tanto, una mayor fuerza de flexión del carpo, necesaria para la propulsión de la mano durante la locomoción cuadrúpeda, suspensoria, e inclusive saltos entre las ramas de los árboles (DEFLER, 2010). El palmar largo es un músculo que se encuentra en el patrón común de primates, aunque se ha reportado ausente en el humano y en primates filogenéticamente más cercanos a él (PERELMAN et al., 2011), como G. gorilla y P. troglodytes (DIOGO et al., 2012; TESTUT & LATARJET, 1984). El flexor radial del carpo se puede presentar originado en el epicóndilo medial del húmero, fascia antebraquial, septos intermusculares adyacentes, e inclusive desde el radio; y su inserción se puede presentar en el II o III metacarpiano.
Los músculos que permiten la flexión de las articulaciones de los dedos son los que más varían entre especies de primates, por un lado, el flexor digital superficial puede presentar entre una y cuatro cabezas de origen, como sucede en H. sapiens (STANDRING, 2008) y en gibones (MICHILSENS et al., 2009) respectivamente, pero finalmente en todas las especies terminan enviando tendones para el II, III, IV y V dedos, y en ocasiones para el I dedo como lo describe YOULATUS (2000) en un A. seniculus. Por otro lado, el flexor digital profundo también presenta diversidad de número de cabezas entre especies de primates, desde una hasta cinco cabezas, distribuidas desde el epicóndilo medial del húmero, radio, ulna y membrana interósea, que envían tendones para el I, II, III, IV y V dedo; y se ha encontrado un músculo independiente para el I dedo en H. sapiens (STANDRING, 2008; TESTUT & LATARJET, 1984) y en gibones, como es el caso del músculo flexor largo del I dedo (pulgar) (DIOGO et al., 2012; DIOGO & WOOD, 2012; HEPBURN, 1892; MICHILSENS et al., 2009).
En la parte caudo-medial del antebrazo de algunos primates también se ha descrito el músculo ancóneo epitroclear, que por su origen e inserción es un músculo accesorio para la extensión del codo, por lo tanto, se pueden encontrar primates que lo presenten y en estos, también involucrar la extensión del codo como una de las funciones dentro del grupo muscular caudo-medial del antebrazo, aunque esta función le corresponde mejor a los músculos caudales del brazo, y por lo tanto, HOWELL & STRAUS (1878) lo describen como parte de la musculatura braquial.
CONCLUSIONES
Los estudios anatómicos de los músculos caudo-mediales de la región antebraquial de primates permiten encontrar semejanzas y variantes interespecíficas e intraespecíficas, las cuales nos dejan establecer que en algún momento provinieron de un ancestro en común, y debido a la gran diversidad de hábitats, han sufrido adaptaciones morfológicas que les permiten movilizarse en ellos. Pero no únicamente se deben haber adaptado los músculos, sino también los huesos y las articulaciones, por lo tanto, sugerimos que la presencia de una o varias cabezas de los diferentes músculos puede ser mejor entendida si involucramos la acomodación adaptativa que han tenido los huesos en sus relieves óseos, para que el primate únicamente necesite una, o por otro lado, haya primates que presentan más de una cabeza en un músculo. Estas variantes interespecíficas e intraespecíficas también deben ser tenidas en cuenta por el cirujano veterinario al momento de necesitar incidir por la región caudo-medial del antebrazo para realizar un mejor procedimiento quirúrgico, bien sea para alcanzar la parte disto-medial del húmero a nivel braquial, y las partes caudo-mediales de radio y ulna a nivel antebraquial, lo que evitará daños ortopédicos por desconocimiento, y así permitirá una mejor y más rápida recuperación y rehabilitación para su regreso al grupo.
REFERENCIAS
ACKERMANN, R.R., 2003.- A Comparative primate anatomy dissection manual. University of Capetown. Disponible en: http://web.uct.ac.za/depts/age/people/dissect.pdf
Consultado en: 17/01/2012. [ Links ]
ANKEL-SIMONS, F., 2007.- Primate Anatomy: An introduction. 3.ed. USA: Elsevier [ Links ]
AVERSI-FERREIRA, T.A.; DA SILVEIRA, P; PIRES, R.M.; AVERSI-F, R.A.G.M.F. & SILVA, Z., 2005a.- Anatomia dos músculos flexores superficiais do antebraço do macaco Cebus. I. J. Morphol., 23 (1): 48. [ Links ]
AVERSI-FERREIRA, T.A.; AVERSI-F, R.A.G.M.F.; SILVA, Z; GOUVEA-E-SILVA, L.F. & PEMHA-SILVA, N., 2005b.- Estudo anatômico de dos músculos profundos do antebraço de Cebus apella (Linnaeus, 1766). Acta Sci. Biol. Sci., 27 (3): 297-301 [ Links ]
AVERSI-FERREIRA, T.A.; VIEIRA, L.G.; PIRES, R.M.; SILVA, Z. & PENHA-S, N., 2006.- Estudo anatómico dos músculos flexores superficiais do antebraço no macaco Cebus apella. Biosci. J., 22 (1): 139-144. [ Links ]
AVERSI-FERREIRA, T.A.; MAIOR, R.S.; CARNEIRO-E-SILVA, F.O.; AVERSI-F, R.A.G.M.F.; TAVARES, M.; NISHIJO, H. & TOMAZ, C., 2011.- Comparative anatomical analyses of the forearm muscles of Cebus libidinosus (Rylands et al. 2000): manipulatory behavior and tool use. Plos One, 6 (7). [ Links ]
BLAND-SUTTON, J., 1897.- Ligaments, their nature and morphology. H.K.Lewis, London. [ Links ]
CHAMPNEYS, F., 1871.- On the muscles and nerves of a chimpanzee (Troglodytes niger) and a Cynocephalus anubis. Part I. J. Anat. Physiol., 6 (Pt1):176-211. [ Links ]
CRIBILLERO, N.; SATO, A. & NAVARRETE, M., 2009.- Anatomía macroscópica de la musculatura del miembro anterior del mono machín blanco (Cebus albifrons).Rev. Investig. Vet. Perú, 20 (2):143-153. [ Links ]
DEFLER, T.R., 2010.- Historia natural de los primates colombianos. 2.ed. Universidad Nacional de Colombia. Facultad de Ciencias, Departamento de Biología, Bogotá [ Links ].
DIOGO, R. & ABDALA, V., 2010.- Muscles of vertebrates: Comparative anatomy, evolution, homologies and development. Science Publishers, USA. [ Links ]
DIOGO, R.; RICHMOND, B.G. & WOOD, B., 2012.- Evolution and homologies of primate and modern human hand and forearm muscles, with notes on thumb movements and tool use. J. Hum. Evol., 63 (1): 64-78. [ Links ]
DIOGO, R. & WOOD, B., 2012.- Comparative anatomy and phylogeny of primate muscles and human evolution. CRC Press. [ Links ]
[ Links ]
DYCE, K.M.; SACK, W.O. & WENSING, C.J.G. 2012.- Anatomía veterinaria. 4.ed. El Manual Moderno, México [ Links ]
FEDERATIVE INTERNATIONAL COMMITTEE ON ANATOMICAL TERMINOLOGY (FICAT), 1998.- Terminologia Anatomica. Thieme Medical Publishers, New York. [ Links ]
EVANS, H.E. & DE LAHUNTA, A., 2013.- Miller's anatomy of the dog. 4.ed. Elsevier Saunders, Missouri. [ Links ]
HEPBURN, D., 1892.- The comparative anatomy of the muscles and nerves of the superior and inferior extremities of the anthropoid apes. J. Anat. Physiol. 26 (1): 149-186. [ Links ]
HILL, W.C., 1959.- The Anatomy of Callimico goeldii (Thomas). T. Am. Philos. Soc., 49 (5): 1-116. [ Links ]
HOWELL, A.B. & STRAUS, W.L., 1878.- The brachial flexor muscles in primates. Proc. U. S. Nat. Mus. Disponible en: http://archive.org/details/cbarchive_122812_brachialflexormusclesinprimate1878 accesado Consultado en: 27 de enero de 2013. [ Links ]
INTERNATIONAL COMMITTEE ON VETERINARY GROSS ANATOMICAL NOMENCLATURE (ICVGAN), 2012.- Nomina Anatomica Veterinaria. 5.ed (revised version). ICVGAN, Hannover. [ Links ]
KARDONG, K., 2011. Vertebrates: Comparative anatomy, function, evolution.6.ed. Mc Graw Hill, New York. [ Links ]
KIMURA, K. & TAKAI, S., 1970.- On the musculature of the forelimb of the crab-eating monkey. Primates, 11 (2): 145-170. [ Links ]
LANGMAN, S., 2007.- Embriología médica. 10.ed. Médica Panamericana, Buenos Aires. [ Links ]
MICHILSENS, F.; VEREECKE, E.E.; D'AOÛT, K. & AERTS. P. 2009.- Functional anatomy of the gibbon forelimb: adaptations to a brachiating lifestyle. J. Anat., 215 (3): 335-354. [ Links ]
PAI, M.M.; PRABHU, L.V.; NAYAK, S.R.; MADHYASTHA, S.; RAJANIGANDHA, V.; KRISHNAMURTHY, A. & KUMAR, A., 2008.- The palmaris longus muscle: its anatomic variations and functional morphology. Rom. J. Morphol. Embryol., 49 (2): 215–217. [ Links ]
PERELMAN, P.; JOHNSON, W.E.; ROOS, C.; SEUÁNEZ, H.; HOVARTH, J.; MOREIRA, M.; KESSING, B.; PONTIUS, J.; ROELKE, M.; RUMPLER, Y, SCHNEIDER, M.; SILVA, A.; O'BRIEN, J.;& PECON-SLATERY, J. 2011.- A molecular phylogeny of living primates. PLoS Genet., 7 (3): e1001342. doi:10.1371/journal.pgen.1001342 [ Links ]
PRIMROSE, A., 1900. The anatomy of the orang outang. University Library, Toronto. Disponible en: http//:www.archive.org/details/cihm_14732
Accesado Consultado en: 15/10/2011. [ Links ]
SINOWATZ, F., 2010.- Musculo-skeletal system: 286-316 (en) HYTTEL, P; SINOWATZ, F. & VEJLSTED, M. (ed.) Essentials of domestic animal embryology. Saunders Elsevier, Oxford. [ Links ]
STANDRING, S., 2008.- Gray's Anatomy: The anatomical basis of clinical practice. 40.ed. Churchill Livingstone: Elsevier. [ Links ]
STEVENS, J.L.; MEYER, D.M. & EDGERTON, V.R., 1977.- Gross anatomy of the forelimb and shoulder girdle of the Galago senegalensis. Primates, 18 (2): 435-452. [ Links ]
TESTUT, L. & LATARJET, A., 1984.- Tratado de anatomía humana. 9.ed. Salvat editores, Barcelona. [ Links ]
YOULATUS, D., 2000.- Functional anatomy of forelimb muscles in Guianan Atelines (Platyrrhini: Primates). Ann. sci. nat., 21 (4): 137-151. [ Links ]

