










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.20 no.2 Manizales July/Dec. 2016
https://doi.org/10.17151/bccm.2016.20.2.4
DOI: 10.17151/bccm.2016.20.2.4
COLONIZATION BY AQUATIC MACROINVERTEBRATES IN LEAVES OF Miconia sp. AND Eucalyptus sp. IN HIGHT MICROCATCHMENT OF CHINCHINÁ RIVER, COLOMBIA
FR: 10-VII-16. FA: 12-VII-16.
1 Programa de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Caldas, Manizales, Caldas, Colombia. E-mail: julirubio87@hotmail.com
2 Magíster en Ciencias Biológicas; Grupo de Investigación BIONAT, Universidad de Caldas, Manizales, Caldas, Colombia. E-mail: anamariamezasalazar@gmail.com
3 Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales; Grupo de Investigación BIONAT, Universidad de Caldas, Manizales, Caldas, Colombia. E-mail: lucimar.dias@ucaldas.edu.co
Los procesos de colonización en ambientes lóticos son regulados por la interacción entre el tipo de vegetación y la cantidad de hojarasca que se incorpora a los ríos e influye sobre la estructura y dinámica del bentos. Los macroinvertebrados bentónicos son un componente importante en los ecosistemas dulceacuícolas, cumplen un papel fundamental en la descomposición y recirculación de nutrientes, además de contribuir en la red trófica como alimento de otros animales. Con el objetivo de comparar la colonización de macroinvertebrados en hojas de árboles nativos e introducidos a través del tiempo en las quebradas El Diamante, La Oliva y el río Chinchiná, se utilizó la metodología de paquetes de hojarasca. La primera quebrada presenta como vegetación ribereña un bosque nativo, la segunda está influenciada por plantaciones de Eucalyptussp. y la tercera no presenta vegetación ribereña. Para el estudio se utilizaron paquetes de tela plástica, donde fueron depositadas hojas de Miconia sp. (especie nativa), hojas de Eucalyptussp. y una mezcla de las dos especies, las cuales demoraron aproximadamente 60 días para descomponerse. En total, se colectaron 8595 individuos de macroinvertebrados, la menor riqueza se presentó a los 30 días y la mayor entre los 45 y 60 días. No se encontraron diferencias significativas en la riqueza y abundancia de macroinvertebrados con respecto a las dos especies vegetales utilizadas. Al evaluar la diversidad (q=0, q=1) no se encontraron diferencias significativas en el número efectivo de géneros o morfotipos entre sustratos (paquetes con hojas nativas, introducidas y mezcladas). El grupo funcional de los recolectores generalistas fue dominante entre los macroinvertebrados colectados.
Palabras clave: descomposición, grupos funcionales, insectos acuáticos, paquetes de hojarasca, vegetación ribereña.
AbstractColonization processes in lotic environments are regulated by the interaction between the type of forest and the amount of litter that goes to the rivers and influences the structure and dynamics of the benthos. Benthic macroinvertebrates are an important component of freshwater ecosystems, playing a fundamental role in decomposition and nutrient recirculation, in addition to contributing to the food chain as food for other animals. Leaf litter methodology was used with the objective of to compare the colonization by macroinvertebrates on native and introduced tree leaves throughout time, in El Diamante Creek, La Oliva Creek, and the Chinchiná River. The first creek presents native forest riparian vegetation, the second creek is influenced by Eucalyptussp. plantations, and the third creek does not present riverine vegetation. In the study, plastic cloth bags were used, in which leaves from a native species (Miconia sp.), leaves from a Eucalyptussp., and a mix of leaves from both species were deposited; the native and introduced leaves decomposed in approximately 60 days. In total, 8595 macroinvertebrate individuals were collected, the least richness was seen after 30 days and the greatest richness was seen between 45 days and 60 days. With diversity (q=0, q=1) no significant differences in the effective number of genres and/or morphotypes found between substrates (packs with native, introduced and mixed leaves). The dominant group within the macroinvertebrates collected was the trophic guild of the generalist collectors.
Key words: trophic groups, aquatic insects, leaf packs, leaf breakdown, riparian vegetation.
Los ríos y las quebradas se caracterizan por presentar una alta diversidad, un alto grado de heterogeneidad temporal-espacial y una interacción con el ambiente terrestre circundante (GILLER & MALMQVIST, 1998). Este ambiente terrestre ribereño presenta gran importancia en los ecosistemas acuáticos, ya que se ha observado que tanto la cantidad y calidad de materia orgánica suministrada por determinado tipo de vegetación, tiene un efecto sobre la estructura y dinámica del bentos (GILLER & MALMQVIST, 1998), lo que influye en el establecimiento de las comunidades biológicas en estos ambientes (MARQUES et al., 1999).
Una de estas comunidades que está integrada al recurso hídrico durante toda su vida es la de los macroinvertebrados acuáticos (ALBA-TERCEDOR, 1996; ROLDÁN, 2003), la cual es considerada uno de los componentes más importantes de los ecosistemas dulceacuícolas en términos de abundancia y diversidad, pues cumplen un rol ecológico destacado en la descomposición y recirculación de nutrientes, además de contribuir en la red trófica como alimento de otros invertebrados y vertebrados (MERRITT & CUMMINS, 1996). A su vez, estos organismos se alimentan de materia orgánica particulada en forma de fragmentos de plantas, algas, bacterias y detritus producidos dentro del arroyo o suministrados desde fuentes externas como la vegetación ribereña (GUEVARA-CARDONA et al., 2006).
De acuerdo con lo anterior y al tener en cuenta la fuente de alimento y las estrategias usadas para la obtención de éste por parte de los macroinvertebrados, se han identificado seis grupos funcionales (CHARÁ-SERNA et al., 2010; CUMMINS et al., 2005); GILLER & MALMQVIST; 1998, TOMANOVA et al., 2006; RAMÍREZ & GUTIÉRREZ-FONSECA, 2014). Los fragmentadores se alimentan masticando detritos de hojas, madera y plantas acuáticas vivas, los recolectores ingieren materia orgánica particulada fina, que toman del sustrato (recolectores de depósitos) o filtran de la columna de agua (filtradores), los raspadores raspan algas y biomasa que están firmemente adheridas de superficies minerales u orgánicas, los perforadores se alimentan de células y tejido de plantas vivas, y su mecanismo de alimentación es la perforación de macrófitas y la succión de fluidos, finalmente, los depredadores y parásitos se alimentan de otros animales.
Para evaluar la estructura trófica y el proceso de colonización de macroinvertebrados se han utilizado paquetes de hojarasca, esta metodología refleja los cambios que ocurren en la composición de la comunidad a lo largo del tiempo (RIBEIRO & UIEDA, 2005; CARVALHO & UIEDA, 2004). Estos cambios están determinados fundamentalmente por el tipo de vegetación ribereña, donde las especies vegetales nativas e introducidas (o exóticas) tienen una descomposición diferente (BRAATNE et al., 2007) y pueden ser más fáciles o difíciles de fragmentar por los invertebrados (GRAÇA, 2001). Dada la importancia de los macroinvertebrados en los ecosistemas dulceacuícolas y su papel en los procesos de descomposición de la materia orgánica, el objetivo de este estudio es comparar la colonización de macroinvertebrados en hojas de árboles nativos e introducidos a través del tiempo.
METODOLOGÍAÁrea de estudio
Se encuentra localizada en los municipios de Villamaría y Manizales, Caldas, en la subcuenca alta del río Chinchiná. La quebrada El Diamante corresponde a la estación uno (05°06'668" N, 75°23'501" W), se encuentra a una altitud de 2810 m y pertenece a la Reserva Forestal Torre Cuatro, su vegetación ribereña está compuesta por un bosque nativo cuyas especies predominantes, son: Erato vulcanica, Miconia sp., y Sauraruia brachybotrys. La quebrada La Oliva corresponde a la estación dos y es reconocida localmente como el río Chinchiná, pero adquiere este nombre al unirse con la quebrada Cajones; está dentro de la Reserva Forestal Bosques de la CHEC (05°01'286" N, 75°25'293" W), se encuentra ubicada a 2314 m de altitud, la estación está influenciada por una plantación de Eucalyptussp. y su vegetación ribereña está compuesta predominantemente por la especie Chusquea scandens. La estación tres se encuentra ubicada en el río Chinchiná (05°01'301" N, 75°26'185" W) a 2206 m de altitud, en esta estación no hay presencia de vegetación ribereña.
Colonización de macroinvertebradosPara realizar el experimento se realizó un premuestreo que consistió en efectuar registros visuales en un margen de 15 m en la quebrada El Diamante, para determinar los árboles dominantes de la zona. La identificación de las especies fue realizada con la ayuda de expertos en sistemática del Herbario FAUC. Se eligió la especie Miconia sp. de tamaño y tiempo de descomposición similar a la especie introducida (Eucalyptussp.) para ser empleada en los paquetes de hojarasca.
Para analizar la colonización de los macroinvertebrados a través del tiempo, se empleó la metodología para paquetes de hojarasca propuesta por MORETTI et al. (2005). Se utilizaron 90 sacos de tela plástica con ojo de malla de 1 cm y dimensión de 10 x 15 cm. Del suelo aledaño a las quebradas se tomaron un conjunto de hojas de una especie nativa de la región (Miconia sp.), un conjunto de hojas de una especie introducida (Eucalyptussp.) y una mezcla de las especies empleadas, para verificar si un recurso heterogéneo presenta una mayor colonización por los invertebrados. En las orillas de cada estación se ubicaron 30 paquetes de hojas, éstos se distribuyeron en cinco hileras de seis bolsas y se alternaron los tratamientos entre Miconia sp. (N), Eucalyptussp. (E) y mixta (M). Un conjunto de seis bolsas se retiraron de cada estación en tiempos predeterminados (10, 20, 30, 45 y 60 días). Al retirarse, las muestras fueron puestas en bolsas de cierre hermético y se fijaron en alcohol (96%). Los organismos fueron separados de la muestra en el Laboratorio de Zoología de la Universidad de Caldas, por lavado en tamices de malla conforme lo propuesto por NESSIMIAN (1993) y de forma manual mediante la observación de las hojas.
Para la determinación taxonómica de los especímenes hasta género o morfotipo se emplearon las claves de MACHADO & RINCÓN (1989), SPANGLER & FRAGOSO (1992), MERRITT & CUMMINS (1996), ROLDÁN (1996), DOMÍNGUEZ & FERNÁNDEZ (2009), PRAT et al. (2009). El material se depositó en la Colección Entomológica del Programa de Biología de la Universidad de Caldas - CEBUC (Registro Humboldt: No 178). La clasificación de los grupos funcionales de los macroinvertebrados acuáticos colectados fue realizada según lo propuesto por CUMMINS et al. (2005), CHARÁ-SERNA et al. (2010), TOMANOVA et al., 2006 y la revisión de RAMÍREZ & GUTIÉRREZ-FONSECA (2014).
Análisis de datosPara analizar la colonización de macroinvertebrados fue comparada la riqueza y la abundancia por medio de un análisis de varianza (ANOVA de dos vías), en este se utilizaron como factores el tipo de sustrato y el tiempo de colonización. El análisis de varianza fue seguido por una prueba de Tukey, por medio de la cual se realizó una comparación por pares, para determinar entre quienes se dan las diferencias con un nivel de significancia de 0,05. Los análisis estadísticos mencionados fueron realizados mediante el programa StatGraphics versión 5.1.
Para estimar la diversidad de los macroinvertebrados asociados al tipo de hoja se determinó el número efectivo de géneros o morfotipos por medio de la diversidad de orden q (qD; JOST, 2006), la cual se basa en el número efectivo de especies y permite hacer comparaciones bajo el mismo nivel de cobertura de muestreo (CHAO et al. 2014). Para este trabajo se consideraron dos medidas de diversidad, la primera medida es la diversidad de orden cero (0D), cuyo valor equivale simplemente a la riqueza de especies (0D = S), la cual está definida por la incidencia de los morfotipos raros en el muestreo o singletons (f1). La segunda medida es la diversidad de orden uno (1D), en la que se da peso a todas los géneros o morfotipos por su frecuencia, sin favorecer desproporcionalmente ya sea a los más comunes o los géneros raros, el cálculo de la diversidad se realizó usando 100 aleatorizaciones con intervalos de confianza del 95% en el programa RStudio versión 0.99.902 – 2009-2016.
Para relacionar los grupos funcionales con los tratamientos de Miconia sp., Eucalyptussp. y mixta, se realizó una prueba Chi-cuadrado para muestras independientes, por medio de tablas de frecuencia, los contrastes se realizaron bajo una significancia de 0,05 con el programa StatGraphics versión 5.1.
RESULTADOSSe colectaron 8595 individuos pertenecientes a 11 órdenes, 32 familias y 56 géneros o morfotipos, entre los cuales la familia Chironomidae fue la más abundante con un 81% del total de individuos, de los que el 50% pertenecen al género Cricotopus y 49% a Polypedilum, seguida por la familia Simuliidae con 10% de los individuos, en la que el género Gigantodax fue el más representativo con 58%.
Para los diferentes sustratos se encontraron 3183 individuos en Eucalyptussp., 3146 en el paquete mixto y 2266 en Miconia sp., con respecto a la abundancia no se presentaron diferencias significativas entre estaciones y entre sustratos (p-valor > 0,05). La riqueza taxonómica encontrada en Eucalyptus sp. y mixta fue de 39 géneros o morfotipos (35%) en cada uno y de 34 (30%) para Miconia sp. La comparación de la riqueza de macroinvertebrados en los tres sustratos a través del tiempo (Figura 1) indica que no hay diferencias en este (p-valor= 0,331, F= 12,26), así como tampoco existen diferencias entre los sustratos (F= 1,495, p-valor= 0,199). La única diferencia significativa encontrada con respecto a la riqueza de macroinvertebrados fue en el sustrato Miconia sp. para las tres estaciones, donde la estación uno difiere de la dos y de la tres (p-valor < 0,05).
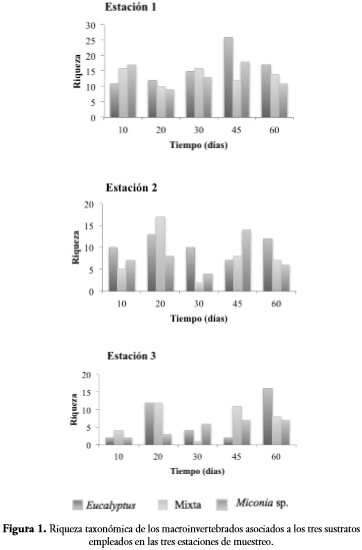
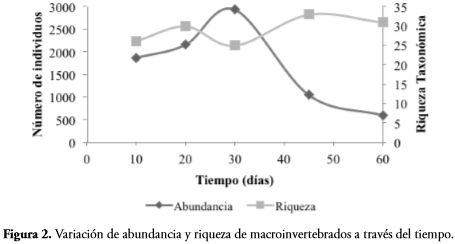
La variación de la comunidad en términos de abundancia durante los dos meses de muestreo presenta una tendencia sigmoidal, se recolectaron 1865 individuos a los diez días, aumentaron hasta los 30 días (2934 individuos) y a partir de ese momento disminuyeron, teniendo en el último muestreo 594 individuos (Figura 2). Los valores de riqueza a través del tiempo fluctuaron entre 25 y 33 géneros o morfotipos, se presentó el menor número a los 30 días y los mayores a los 45 y 60. Tanto la riqueza como la abundancia no tuvieron diferencias significativas en el tiempo (F= 0,929, p-valor= 0,608 y F= 0,169, p-valor= 0,700, respectivamente).
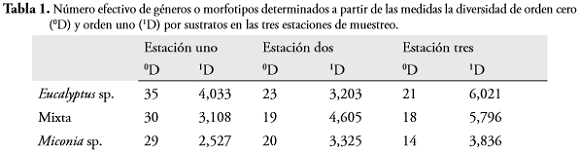
Al evaluar el número efectivo de géneros o morfotipos mediante la diversidad de orden cero (0D), el sustrato Eucalyptussp. fue el que obtuvo mayor riqueza de macroinvertebrados en las tres estaciones, seguido por el tratamiento mixto y por último el sustrato Miconia sp. La diversidad de orden uno (1D) que incluye todos los géneros y su abundancia relativa, mostró este mismo patrón en las estaciones uno y tres, donde Eucalyptussp. y Miconia sp. obtuvieron los mayores y menores valores de géneros o morfotipos efectivos, respectivamente. En la estación dos, el mayor número efectivo de géneros o morfotipos se obtuvo en el tratamiento mixto, seguido por el sustrato Miconia sp. y por último el sustrato Eucalyptussp. (Tabla 1).
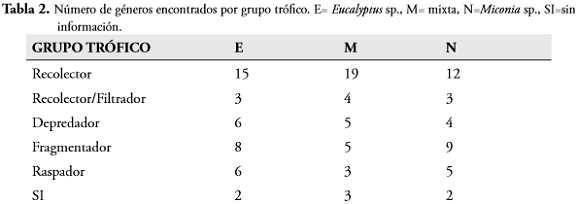
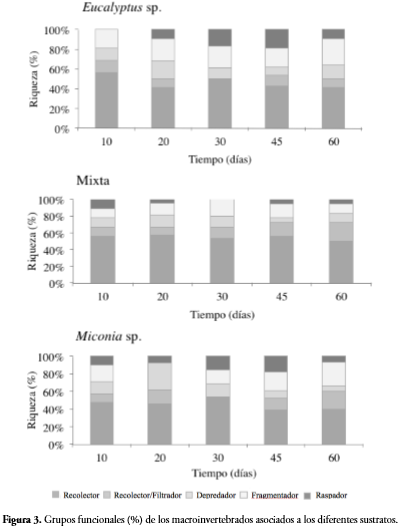
En cuanto a los grupos funcionales, los recolectores generalistas dominaron en los tres sustratos (83,63%), seguido por los fragmentadores (40%), depredadores (27,27%), raspadores (25,45%) y recolectores/filtradores (18,18%). Las diferencias presentadas entre los grupos se dan entre los recolectores con los demás grupos (p < 0,01). Para cada sustrato se evaluaron las diferencias de los grupos funcionales en el tiempo, los valores hallados muestran que no existen diferencias significativas (F= 2,7890, p-valor= 0,0618 para Eucalyptussp., F= 1,058, p-valor= 0,409 en mixta y F= 2,444, p-valor= 0,088 para Miconia sp.). Los resultados de la prueba de Chi-cuadrado y la tabla de contingencia muestran que no existe dependencia entre los grupos funcionales y los sustratos empleados (p-valor=0,925 df= 686) (Tabla 2).
DISCUSIÓN
Los resultados del presente trabajo demuestran que la familia Chironomidae es la más representativa en el experimento, se encuentra desde los primeros días de muestreo y no muestra diferencias significativas en la preferencia de sustrato; este dato es corroborado por diferentes autores que han trabajo con paquetes de hojarasca, los cuales también encontraron una alta dominancia de esta familia en los sustratos empleados en sus investigaciones (MATHURIAU & CHAUVET, 2002; CARVALHO & UIEDA, 2004; NESSIMIAN & HENRIQUES-OLIVEIRA, 2005; CALLISTO et al., 2007; CARVALHO et al., 2008; TORRES & RAMÍREZ, 2014).
En este estudio no se encontraron diferencias significativas con respecto a la riqueza y abundancia de macroinvertebrados en los tres sustratos (nativo, introducido y mixto). Resultados similares fueron encontrados por MATHURIAU & CHAUVET (2002) en Colombia, SCHÜLER (2010) en Brasil y BRAATNE et al. (2007) en Estados Unidos, donde tampoco se encontró relación entre los macroinvertebrados acuáticos y un sustrato en particular. La mayor abundancia de macroinvertebrados se encontró en la mitad del experimento (30 días), momento cuando la hojarasca ya ha sufrido procesos de fragmentación y se encuentra como materia orgánica particulada fina, la cual es alimento para organismos recolectores como Chironomidae, quienes fueron los más abundantes en esta etapa de colonización.
La preferencia de los macroinvertebrados por Miconia sp. en la estación uno, probablemente se da por la dominancia de esta especie de árbol en la zona, lo que hace que siempre esté disponible para los invertebrados y por lo tanto estén altamente adaptados al recurso. Adicionalmente, el bosque de ribera presente en esta estación puede generar un aumento en el aporte de hojarasca que es la principal fuente de energía de los ecosistemas lóticos y además brinda una mayor disponibilidad de recursos para los macroinvertebrados (WINTERBOURN & TOWNSEND, 1991; OSBORNE & KOVACIC, 1993).
Las medidas de 0D y 1D y la riqueza y abundancia de macroinvertebrados en el experimento, evidencian que el proceso de colonización es similar entre estaciones, posiblemente las diferencias estén representadas en los géneros y morfotipos presentes en cada una y que efectúan el proceso de colonización. La estación 1 presenta el mayor número de géneros o morfotipos exclusivos (12 taxa), entre los que se encuentran Helicopsyche, Macronema y Metrichia, seguida por las estaciones 2 y 3 con 7 y 2 géneros o morfotipos exclusivos, respectivamente; esos resultados son similares a lo encontrado por MEZA et al. (2012) al evaluar la comunidad de macroinvertebrados en las mismas estaciones. Además, otros trabajos realizados por LACAN et al. (2010) y EYES-ESCALANTE et al. (2012) tampoco han encontraron diferencias entre sustratos nativos e introducidos, ni entre sitios con plantaciones y sin estas, empleando especies introducidas del género Eucalyptuscomparadas con especies nativas en ríos de California (Estados Unidos) y Santa Marta (Colombia).
La dominancia de los recolectores en todas las estaciones evaluadas y sustratos utilizados, puede ser corroborada por autores como DANGLES et al. (2001), MATHURIAU & CHAUVET (2002), RIVERA & SEGNINI (2004), RIVERA et al. (2009), MORETTI et al. (2005). Probablemente esto se deba a que en los sistemas tropicales la presencia de hojarasca contribuye al aumento de la disponibilidad de sustrato, aportando materia orgánica particulada que se convierte en la fuente de alimento de mayor disponibilidad (CHARÁ-SERNA et al., 2010). La baja representación de los fragmentadores ha sido documentada por otros autores en el trópico (RUEDA-DELGADO et al., 2006; WANTZEN & WAGNER, 2006); sin embargo, la importancia ecológica de este grupo no se refleja en la abundancia de estos sino en la biomasa que aportan (RODRÍGUEZ-BARRIOS et al., 2011; CHARÁ-SERNA et al., 2012).
Los grupos funcionales en el tiempo y en los sustratos empleados no presentaron variaciones, posiblemente se deba a que en todos los días del experimento el recurso alimenticio no fue una limitante y el tipo de hoja no restringió la presencia de algún grupo funcional, lo que coincide con lo encontrado en Brasil por RIBAS (2005). Por lo tanto, en el presente trabajo el tipo de hoja (nativa e introducida) no evidenció un efecto sobre la colonización de macroinvertebrados en las quebradas estudiadas.
RECOMENDACIONESSe recomienda realizar estudios que permitan comprender la relación entre la presencia de los macroinvertebrados en la hojarasca nativa o exótica y su función como alimento o refugio, lo que puede proveer información relevante a cerca de la importancia del recurso alóctono para los macroinvertebrados acuáticos.
AGRADECIMIENTOSA la doctora Beatriz Toro, Jeymmy Walteros, Héctor Chica, doctor Tito Bacca, María del Carmen Zúñiga, Orlando Caicedo, Néstor Fabio Alzate, Marco Tulio Jaramillo, Bedir Martínez, Luis Eduardo Robledo, José Luis Benavides, Camilo Andrés Llano, Patricia Jiménez, Ximena Villada, Mauricio Escobar, Juan Pablo Jaramillo, Mario López y Don Jairo por su colaboración en el proyecto. A la Universidad de Caldas por la infraestructura concedida para la realización del trabajo.
ALBA-TERCEDOR, J., 1996.- Macroinvertebrados acuáticos y calidad de las aguas de los ríos. Libro de resúmenes del IV Simposio el agua en Andalucía (Siaga), Almería, 2: 202-213. [ Links ]
BRAATNE, J.H., SULLIVAN, S.M.P. & CHAMBERLAIN, E. , 2007.- Leaf Decomposition and Stream Macroinvertebrate Colonisation of Japanese Knotweed, an Invasive Plant Species. Int Rev Hydrobiol., 92 (6): 656-665. [ Links ]
CALLISTO, M., GONÇALVES, J.F. & GRAÇA, M.A.S., 2007.- Leaf litter as a possible food source for chironomids (Diptera) in Brazilian and Portuguese headwater streams. Rev. bras. zool., 24 (2): 442-448. [ Links ]
CARVALHO, E.M. & UIEDA, V.S., 2004.- Colonização por macroinvertebrados bentônicos em substrato artificial e natural em um riacho da serra de Itatinga, São Paulo, Brasil. Rev. bras. zool., 21 (2): 287-293. [ Links ]
CARVALHO, E.M., UIEDA, V.S. & MOTTA, R.L., 2008.- Colonization of rocky and leaf pack substrates by benthic macroinvertebrates in a stream in Southeast Brazil. Bioikos, 22 (1): 37-44. [ Links ]
CHARÁ-SERNA, A.M., CHARÁ, J., ZÚÑIGA, M. D.C., PEARSON, R.G., & BOYERO, L., 2012.- Diets of leaf litter-associated invertebrates in three tropical streams. Ann Limnol Int J Lim., 48 (2): 139-144. doi: 10.1051/limn/2012013 [ Links ]
CHARÁ-SERNA, A.M., CHARÁ, J.D., ZÚÑIGA, M.C., PEDRAZA, G.X. & GIRALDO, L.P., 2010.- Clasificación trófica de insectos acuáticos en ocho quebradas protegidas de la ecorregión cafetera colombiana. Univ. Sci., 15 (1): 27-36. [ Links ]
CHAO, A., GOTELLI, N.J., HSIEH, T. C., SANDER, E.L., MA, K.H., COLWELL R.K. & ELLISON, A.M., 2014.- Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol. Monogr., 84 (1): 45-67. [ Links ]
CUMMINS, K.W., MERRIT, R.W. & ANDRADE, P., 2005.- The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in southeast Brazil. Stud Neotrop Fauna Environ., 40 (1): 71 - 90. [ Links ]
DANGLES, O., GUEROLD, F. & USSEGLIO-POLATERA, P., 2001.- Role of transported particulate organic matter in the macroinvertebrate colonization of litter bags in streams. Freshwater Biol., 46: 575-586. [ Links ]
DOMÍNGUEZ, E. & FERNÁNDEZ, H.R., 2009.- Macroinvertebrados bentónicos sudamericanos: sistemática y biología. Fundación Miguel Lillo. San Miguel de Tucumán. [ Links ]
EYES-ESCALANTE, M., RODRÍGUEZ-BARRIOS, J. & GUTIÉRREZ-MORENO L.C., 2012.- Descomposición de la hojarasca y su relación con los macroinvertebrados acuáticos del río Gaira (Santa Marta Colombia). Acta biol. Colomb., 17: 77 - 92. [ Links ]
GILLER, P.S. & MALMQVIST, B., 1998.- The Biology of Streams and Rivers. Oxford University Press. Oxford, UK. [ Links ]
GUEVARA-CARDONA, G., JARA, C., MERCADO, M. & ELLIOTT, S., 2006.- Comparación del macrozoobentos presente en arroyos con diferente tipo de vegetación ribereña en la Reserva Costera Valdiviana, Sur de Chile. ACL - Limnos 1: 98-105. [ Links ]
GRAÇA, M.A.S., CRESSA, C., GESSNER, M.O., FEIO, M. J., CALLIES, K. A., & BARRIOS, C., 2001.- Food quality, feeding preferences, survival and growth of shredders from temperate and tropical streams. Freshwater Biol, 46: 947-957. [ Links ]
JOST, L., 2006.- Entropy and diversity. Oikos, 113: 363-374. [ Links ]
LACAN, I., RESH, V.H. & MCBRIDE, J.R., 2010.- Similar breakdown rates and benthic macroinvertebrate assemblages on native and Eucalyptus globulus leaf litter in Californian streams. Freshwater Biol., 55: 739-752. [ Links ]
MACHADO, T.A. & RINCÓN, J., 1989.- Distribución ecológica e identificación de los coleópteros acuáticos en diferentes pisos altitudinales del departamento de Antioquia. Universidad de Antioquia, Facultad de Ciencias Exactas y Naturales, Departamento de Biología. Medellín. [ Links ]
MARQUES, M.G.S.M., FERREIRA, R.L. & BARBOSA, F.A.R., 1999.- A comunidade de Macroinvertebrados aquáticos e características Limnológicas das Lagoas Carioca e da Barra, Parque Estadual do Rio Doce, MG. Rev. bras. biol., 59 (2): 203-210. [ Links ]
MATHURIAU, C. & CHAUVET, E., 2002.- Breakdown of leaf litter in a neotropical stream. J N AM Benthol Soc., 21 (3): 384-396. [ Links ]
MERRITT, R.W. & CUMMINS, K.W., 1996.- An Introduction to the Aquatic Insects of North America. 3 ed. Kendall/Hunt Publishing. Iowa. [ Links ]
MEZA-S, A.M., RUBIO-M, J., G-DIAS, L., & WALTEROS, J.M. 2012.- Calidad de agua y composición de macroinvertebrados acuáticos en la subcuenca alta del río Chinchiná. Caldasia. 34(2): 443-456. [ Links ]
MORENO, C.E., 2001.- Métodos para medir la biodiversidad. M&T-Manuales y Tesis SEA. Zaragoza. [ Links ]
MORETTI, M.S., GONÇALVES, J.F. & CALLISTO, M., 2005.- Invertebrados associados a detritos foliares em decomposição em um córrego tropical de altitude. In: Moretti M.S. (eds.), Decomposição de detritos foliares e sua colonização por invertebrados aquáticos em dois córregos na Cadeia do Espinhaço (MG). Universidade Federal de Minas Gerais. Belo Horizonte. 32-50 p. [ Links ]
NESSIMIAN, J.L., 1993.- Estrutura, composição e dinâmica da fauna invertebrada bentônica de um brejo entre dunas no litoral do Estado do Rio de Janeiro. Tese de Doutorado. Universidade Estadual Paulista Júlio de Mesquita Filho. Rio Claro. [ Links ]
NESSIMIAN, J.L. & HENRIQUES-OLIVEIRA, A.L., 2005.- Colonização do "litter" de Eleocharis sellowiana Kunth. (Cyperaceae) por larvas de Chironomidae (Diptera) em um brejo no litoral do estado do Rio de Janeiro. Entomol. Vectores, 12 (2): 159-172. [ Links ]
OSBORNE, L & KOVACIC, D., 1993.- Riparian vegetated buffer strips in water quality restoration and stream management. Freshwater Biol., 29 (2): 243-258. [ Links ]
PRAT, N., ACOSTA, R., VILLAMARÍN, C. & RIERADEVALL, M., 2009.- Guía para el reconocimiento de las larvas de Chironomidae (Diptera) de los ríos altoandinos de Ecuador y Perú. Clave para la determinación de los géneros. Universidad de Barcelona. Grupo de Investigación F.E.M. Departamento de Ecología. Barcelona. [ Links ]
RAMÍREZ, A., & GUTIÉRREZ-FONSECA, P.E., 2014.- Functional feeding groups of aquatic insect families in Latin America: a critical analysis and review of existing literature. Rev. Biol. Trop., 62: 155-167. [ Links ]
RIBAS, A.C., 2005.- Sucessão de macroinvertebrados ao longo da decomposição da serapilheira em uma nascente de região tropical. Tese de doutorado. Universidade Federal de Mato Grosso do Sul, Campo Grande. [ Links ]
RIBEIRO, L.O. & UIEDA, V.S., 2005.- Estrutura da comunidade de macroinvertebrados bentônicos de um riacho de serra em Itatinga, São Paulo, Brasil. Rev. bras. zool., 22 (3): 613-618. [ Links ]
RIVERA, R.A. & SEGNINI, S.E., 2004.- Estructura y composición de la comunidad de macroinvertebrados bentónicos en ríos de páramo y zonas boscosas, en los Andes venezolanos. Universidad de Los Andes. OAI Repositorio Institucional Referencial de Tesis y Trabajos (Sistema LIBRUM), SERBIULA. Mérida. [ Links ]
RIVERA, C.A., ZAPATA, A.M., PÉREZ, D., MORALES, Y., OVALLE, H. & ÁLVAREZ, J.P., 2009.- Caracterización limnológica de humedales de la planicie de inundación del río Orinoco (Orinoquía, Colombia). ABC, 15 (1). Disponible en: http://www.revistas.unal.edu.co/ [ Links ]
RODRÍGUEZ-BARRIOS, J., OSPINA-TORRES, R., GUTIÉRREZ, J., & OVALLE, E., 2007.- Densidad y biomasa de macroinvertebrados acuáticos derivantes en una quebrada tropical de montaña. Bogotá, Colombia. Caldasia, 29 (2): 397-412. [ Links ]
ROLDÁN, G.A., 1996.- Guía para el estudio de los macroinvertebrados acuáticos del departamento de Antioquia. Universidad de Antioquia. Fondo FEN Colombia. COLCIENCIAS. Medellín. [ Links ]
ROLDÁN, G.A., 2003.- Bioindicación de la calidad del agua en Colombia. Universidad de Antioquia. Medellín. [ Links ]
RUEDA-DELGADO, G., WANTZEN, K.W., & TOLOSA, M.B., 2006.- Leaf-Litter decomposition in an Amazonian floodplain stream: effects of seasonal hydrological changes. J North Am Benthol Soc., 25: 233-249. [ Links ]
SCHÜLER, C., 2010.- Quebra foliar e colização de macroinvertebrados em riachos do bioma Pampa. Dissertação de Mestrado, Programa de Pós-Graduação em Ecologia, Universidade Federal do Rio Grande do Sul, Brasil. [ Links ]
SPANGLER, P.J. & FRAGOSO, S.S., 1992.- The aquatic beetle subfamily Laraine (Coleoptera: Elmidae) in México, Central America and the West Indies. Smithsonian Contributions to Zoology. Washington, D.C. [ Links ]
TOMANOVA, S., GOITIA, E. & HELESIC, J., 2006.- Trophic levels and fuctional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia, 556: 251-264. [ Links ]
TORRES, P.J., RAMÍREZ, A., 2014.- Land use effects on leaf litter breakdown in low-order streams draining a rapidly developing tropical watershed in Puerto Rico. Rev. Biol. Trop., 62: 129-142. [ Links ]
WANTZEN, K.M., & WAGNER, R., 2006.- Detritus processing by invertebrate shredders: a neotropical-temperate comparison. J North Am Benthol Soc, 25: 216-232. [ Links ]
WINTERBOURN, M. & TOWNSEND, C., 1991.- Streams and rivers: One way flow systems. In: Barnes, R; Mann, K. eds. Fundamentals of aquatic ecology. Oxford, UK, Blackwell Scientific Publications. [ Links ]

