










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.20 no.2 Manizales July/Dec. 2016
https://doi.org/10.17151/bccm.2016.20.2.19
DOI: 10.17151/bccm.2016.20.2.19
GENERA OF REDUVIIDAE (HEMIPTERA: HETEROPTERA) IN THREE FRAGMENTS OF TROPICAL DRY FOREST (TDF) IN THE DEPARTMENT OF ATLANTICO, COLOMBIA
1 Semillero de Investigación de artrópodos del Caribe colombiano NEOPTERA, Grupo Biodiversidad del Caribe colombiano. Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia. mcristy16@gmail.com ORCID: 0000-0002-1753-346, ORCID: 0000-0002-2455-9154
2 Grupo de Investigación Ecología y Sistemática Neotropical "ECOSIN". Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla, Colombia. neyjosemartinez@gmail.com
Se analizó la variación espacio-temporal de la diversidad de Reduviidae (Hemiptera) en tres fragmentos de BST (Reserva Campesina la Montaña= RCM, La Esmeralda= L.E y Puerto Colombia= P.Col) en el departamento del Atlántico entre noviembre (2012) a junio (2013). En cada fragmento se diseñó un transecto de 600 m de largo, donde se marcaron 10 puntos distanciados 50 m y se realizó la captura de los Reduviidae con red de perturbación de follaje, cernido de hojarasca y recolecta manual. En este mismo transecto, se colocaron cuatro trampas de luz (dos blanca y dos violeta) distanciadas 150 m. Se capturaron 265 individuos distribuidos en 19 géneros y nueve subfamilias, de las cuales la más diversa (7 géneros) fue Harpactorinae, lo que representa el 37% de la riqueza total. El género más abundante fue Zelurus Hahn con 143 individuos, el cual presentó una correlación significativa y directa con las precipitaciones y mostró estacionalidad con la llegada de las lluvias en el área de estudio. Por otro lado, la mayor riqueza de Reduviidae se presentó en el fragmento de L.E y RCM con 13 géneros cada uno, mientras que la mayor abundancia (155) se presentó en mayo. La composición de Reduviidae presentó variación espacio-temporal en los fragmentos de BST del departamento del Atlántico, demostrándose que los géneros de esta familia dependen tanto de la disponibilidad de microhábitats, recursos alimenticios y los cambios fenológicos en la vegetación en este tipo de ecosistema.
Palabras clave: Harpactorinae, trampa de luz, variación espacio-temporal, Zelurus.
AbstractThe variation of diversity of Reduviidae (Hemiptera) in three fragments of Tropical Dry Forest (Reserva Campesina La Montaña= RCM, La Esmeralda= L.E and Puerto Colombia= P.Col) from November 2012 to June 2013 in the department of Atlántico was analyzed. A lineal transect of 600 m was installed in each fragment. Ten points were marked and separated every 50 m. The Reduviidae were collected those points using hitting (agitation of bushes), sifting of leaf litter and manual gathering. Four light traps were also installed (two white and two purple) every 150 meters. Two hundred and sixty-five specimens were captured. They represented 19 genera and 9 subfamilies. The most diverse subfamily was the Harpactorinae with 37% of the total 143. The Zelurus Hahn was the most common genera with 143 specimens which was directly related to the rainy season in the area. On the other hand, the L.E and the RCM fragments had the highest population of Reduviidae with 13 genera in each one. The highest population was in May with 155 specimens. The composition of Reduviidae varied during the time and through the space in the DTF fragments in the department of Atlantico which shows that this family depend on micro-habitat availability, food sources and changes in the vegetation in the these ecosystems.
Key words: Harpactorinae, seasonality, light trap, spatial and temporal variation, Zelurus.
El bosque seco tropical (BST) es un ecosistema de suma importancia, ya que constituye una amplia cobertura, que abarca el 42% de los ecosistemas tropicales en todo el mundo, lo que representa globalmente el segundo tipo de bosque tropical (MILES et al., 2006). Actualmente, los BST aparecen de forma discontinua, en áreas de diferentes tamaños y con diferentes niveles de aislamiento. En Colombia, el 98,5% del BST ha desaparecido en su mayoría y los pocos remanentes que aún se conservan se encuentran dispersos en pequeños parches desconectados entre sí. Se pueden localizar algunos en la zona costera y serranías bajas de la región Caribe colombiana, así como en los valles interandinos y algunos pocos en las islas de San Andrés y Providencia (DÍAZ, 2006).
La presión que se viene presentando sobre los fragmentos de BST los está llevando cada vez más a la reducción, debido a que sus tierras están siendo utilizadas como zonas para la ganadería, el cultivo y la extracción de material vegetal que provoca la alteración del hábitat; afectando a las comunidades de animales que en él se encuentran (DIRZO et al., 2011; GARCÍA et al., 2014). Entre estos se encuentran los insectos, los cuales constituyen una parte importante de la diversidad biológica y algunos de ellos son considerados indicadores de heterogeneidad de los hábitats, de la diversidad de ecosistemas y del estrés ambiental (CANO & SCHUSTER, 2007). Dentro de este grupo de artrópodos, se encuentran los chinches de la familia Reduviidae, los cuales pueden verse afectados por la estrecha relación que tienen con sus presas y las variables ambientales que fluctúan en el bosque seco, de acuerdo con los niveles de precipitación que se presenten en este tipo de ecosistema.
Los estudios sobre la diversidad de la familia Reduviidae en el BST es importante por ser una de las más grandes y morfológicamente más diversas dentro de los heterópteros, constituyendo unos de los grupos más variables; puesto que poseen hábitos depredadores y hematófagos (SCHUH & SLATER, 1995; FORERO, 2004). Los Reduviidae se encuentran distribuidos en una gran cantidad de hábitats y presentan variedad en formas y tamaños. Adicionalmente presentan un gran valor ecológico, económico y biomédico, por su estrecha asociación a ciertas presas y microhábitats; así como algunos son considerados agentes de control biológico (FORERO, 2008) para uso en agricultura y de gran importancia en salud pública, con especies vectores causantes de la enfermedad de Chagas (LENT & WYGODZINSKY, 1979; SCHUH & SLATER, 1995; FORERO, 2004).
El estudio de los Reduviidae en Colombia ha sido muy fragmentado y disperso y las pocas investigaciones realizadas para ciertas subfamilias están enfocadas en la descripción de nuevas especies (FORERO, 2006; CHAVERRA-RODRÍGUEZ et al., 2010) y la sistemática de géneros específicos (CASTRO-HUERTA, 2010). Por otro lado, solo se ha observado un amplio interés sobre la subfamilia Triatominae, debido a su gran importancia en salud pública y control de especies vectores; sobre todo la distribución e importancia ecoepidemiológica (MOLINA et al., 2000; GUHL et al., 2007, 2009; PARRA-HENAO et al., 2009; CANTILLO-BARRAZA et al., 2010).
Debido a la poca información sobre esta familia de hemípteros en fragmentos BST en las zonas bajas del Caribe colombiano; se analizó la variación espacial y temporal de la diversidad de géneros de Reduviidae (Hemiptera: Heteroptera) en tres fragmentos de BST en el departamento del Atlántico (Colombia).
MATERIALES Y MÉTODOSÁrea de estudio
En el departamento del Atlántico se escogieron tres fragmentos de BST de acuerdo con su extensión, cobertura, facilidad de acceso y la cercanía con asentamientos humanos (Figura 1). La vegetación dominante en los tres fragmentos es el BST descrito por DIRZO et al. (2011), caracterizado por una pérdida del follaje durante la época seca y la permanencia de las hojas durante la época lluviosa.
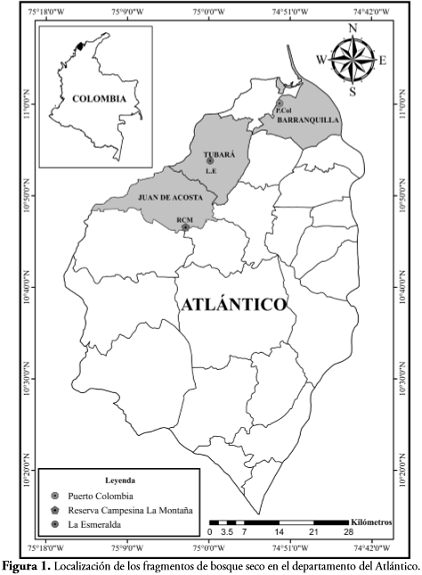
El primer fragmento se encuentra en la Reserva Campesina La Montaña (RCM) (Figura 2), en el municipio de Juan de Acosta, localizada a 10°46'2.6"N 75°0.2'34"O, a una altitud entre 177 y 250 m. El área cuenta con una extensión de 365 ha, de las cuales 47 están ocupadas por un fragmento de bosque seco, declarado en categoría de zona de reserva campesina (Decreto 1777 de 1996). La RCM presenta un tipo de suelo arcilloso-arenoso (ArA), relieve ondulado y hace parte de los ecosistemas de serranías, con pendientes entre 12% y 25%, con grado de erosión moderado (PANZA et al., 2001) y presenta una temperatura anual promedio de 28°C. De acuerdo con los últimos datos obtenidos por MARTÍNEZ et al. (2012), el régimen pluviométrico promedio en la zona es de 179 mm/año; con lluvias bimodales (dos picos a lo largo del año) entre los meses de abril a junio y de septiembre a noviembre. Los valores de humedad relativa presentan una mayor inestabilidad atmosférica en los períodos abril-mayo y agosto-noviembre, con un rango promedio entre 79% y 82% anual (Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia, IDEAM). El fragmento de bosque presenta una cobertura vegetal continua, observándose tres estratos vegetales conformados por el; predominando el bejuco bignoniáceo (Adenocalymna inundatum), poaceas y ocasionalmente se presenta una especie de helecho (Blechnum sp). En el estrato arbustivo se presenta la dominancia de Capparis baducca, C. odoratissima y C. indica. Para el estrato arbóreo se observa la dominancia de especies como jobo (Spondias mombim), indio encuero (Bursera simaruba) y majagua (Pseudobombax septenatum).
El segundo fragmento se encuentra en el corregimiento de Corrales de San Luis, municipio de Tubará. El parche de bosque se denomina La Esmeralda (L.E) (Figura 2), y se encuentra localizado a 10°54'18"N 75°00'10"O a una altitud de 125 msnm en el complejo Serranías de Tubará, que hace parte de las estribaciones septentrionales de la Cordillera Occidental. Presenta una extensión de 30 ha; dentro de las cuales algunas son utilizadas para potreros y cultivos como yuca (Manihot esculenta), millo (Sorghum bicolor), maíz (Zea mays), fríjol (Phaseolus vulgaris), guandul (Cajanus cajan). Además, se encuentra rodeado de árboles frutales como el mango (Mangifera indica) y mamón (Melicoccus bijugatus) y de especies como matarratón (Gliricidia sepium), sabanero (ecoma stans), trébol (Platymiscium pinnatum), utilizadas principalmente para la producción de carbón vegetal y extracción de madera; lo cual ha influido en la pérdida de especies vegetales nativas y en la modificación de la estructura de la vegetación.
Este fragmento presenta terrenos de pendientes ligeramente moderadas, pequeñas ondulaciones, con alturas mayores de 50 m y menores de 200 m (IGAC, 1994). Presenta una precipitación promedio de 179 mm/mes, con una temperatura que oscila entre los 25-28°C y humedad relativa con valores entre 75%-88% (IGAC, 1994, HIJMANS et al., 2004). Aproximadamente el 20% del área total está cubierta por vegetación en estado de sucesión más avanzado, donde las familias vegetales más representativas son Capparaceae, Bignonaceae, Sapindaceae, Fabaceae y Moraceae; destacándose especies como Hura crepitans, Enterolobium cyclocarpum, Brosimum alicastrum, Capparis frondosa, Melicoccus bijugatus, Coursetia ferruginea.
El último fragmento (P.Col) se encuentra ubicado en el km 6, vía a Puerto Colombia (Figura 2), en la finca Vista al Mar en el municipio de Puerto Colombia, localizado a 11º00' 6.14"N 74º52'11.3"O. Presenta una extensión de 29 ha, con terrenos planos y pocos ondulados y un clima cálido con temperatura media de 27,8°C. La zona presenta un paisaje modificado antrópicamente, con la presencia de linderos dentro del bosque y cultivos temporales de patilla (Citrullus lanatus), maíz (Zea mays), guandul (Cajanus cajan) y melón (Cucumis melo) con presencia de árboles de tamarindo (Tamarindus sp ) y ciruelos (Spondias purpurea). Adicionalmente, se observaron arbustos juveniles de uvita (Cordia dentata), abundantes lianas y formaciones de matorral, destacándose especies de los géneros Capparis, Iresine, Croton, Nee, Hura, Pseudobombax, Prosopis, Jaquinia, Peresquia y Gliricidia. En el área se presentan quemas esporádicas, que coinciden con la siembra de los cultivos y la extracción de madera para la realización de carbón vegetal.

Diseño y método de captura
En cada fragmento se realizaron ocho muestreos entre noviembre de 2012 y junio de 2013, de los cuales cuatro fueron realizados durante la época seca y los restantes durante las lluvias, con una duración de dos días para cada uno durante luna nueva. Lo anterior, debido a que algunos insectos en el BST en el Caribe colombiano tienen su periodo de actividad sincronizada con la presencia o ausencia de lluvias (BARRAZA et al., 2010).
Para los muestreos diurnos, en cada parche de bosque se estableció un transecto de 600 m de largo por 20 m de ancho, donde se marcaron diez puntos distanciados 50 m y en cada uno se realizó la captura de los especímenes por medio de una red para perturbación de follaje (JPF) y captura manual (CM). La JPF consistió en una red con un diámetro de 50 cm, un cono de 30 cm y un mango por donde se sostiene de 50 cm de largo. Para esta técnica se utilizó un pedazo de madera, con el cual se realizaron 20 golpes repetitivos a los arbustos y vegetación baja que se encontraba en cada punto, con el fin de que los Reduviidae que cayeran en la jama fueran capturados con ayuda de pinzas. Este procedimiento se realizó tres veces para un total de 60 golpes/puntos y 600 por transecto. Adicional a esto, en cada punto se invirtieron 20 minutos por recolector (día) en la revisión manual de piedras, hojarasca, tronco en el suelo, corteza, ramas, hojas, flores, nidos y madrigueras; para una inversión total de 200 minutos por transecto/muestreo.
Adicionalmente, en cada punto se midieron 10 L de hojarasca que fueron cernidos en un tamizador Winkler, devolviendo la hojarasca sobrante al bosque. El material recolectado fue revisado en el laboratorio en bandejas blancas con la ayuda de pinceles y pinzas. Por otro lado, se marcaron cuatro puntos distanciados 150 m aproximadamente y en cada uno se colocó una trampa de luz (T.L) descrita por MARTÍNEZ et al. (2009), para un total de cuatro trampas (dos de luz blanca y dos violeta) por transecto. Las T.L fueron colocadas de manera alternada y encendidas a partir de las 6:00 p.m. Adicional a esto, en cada punto se realizó una revisión manual de 15 min alrededor de las trampas, con el fin de complementar el muestreo. Por otro lado, tanto en los muestreos diurnos como nocturnos, se tomaron parámetros de temperatura (°C) y humedad relativa (%), con la ayuda de un termohigrómetro marca Jumbo-Display. También se midió la precipitación diaria en la zona con la ayuda de un prototipo de pluviómetro (marca Hellman), instalado cerca de cada uno de los fragmentos.
Todos los individuos se guardaron en recipientes debidamente rotulados con alcohol al 70% para su posterior identificación en el laboratorio. Fueron llevados hasta género con las clave de FORERO (2004). En el caso de triatóminos, se utilizaron las de LENT & WYGODZINSKY (1979) y SOTO-VIVAS (2009). Para la corroboración de los especímenes se contó con la colaboración del especialista nacional Dimitri Forero. El material de cada uno de los géneros reposa en la colección entomológica del museo entomológico de la Universidad Javeriana (MUJ) y en la colección de la Universidad del Atlántico (CEUA- UARC).
Análisis de los datosLa diversidad de Reduviidae se determinó como el número de géneros encontrados por cada técnica de captura, fragmento de bosque y muestreo. Se realizó una matriz de presencia-ausencia y a partir de estos resultados fue elaborada una curva de acumulación de géneros para toda el área de estudio y para cada uno de los fragmentos, utilizando los estimadores no paramétricos ICE y Chao 2, los cuales fueron usados por proveer un menor sesgo y una mayor exactitud, aun en el análisis de muestras pequeñas (COLWELL & CODDINGTON, 1994); así como el comportamiento de los géneros Uniques y Duplicates, los cuales fueron calculados con el programa ESTIMATE 9.1 (COLWELL, 2009).
Por otro lado, con el fin de observar un patrón espacio-temporal en la composición de géneros de Reduviidae se determinó la diversidad beta (ß) en términos de similitud a través de una matriz de presencia-ausencia. Una vez construida la matriz de similitud, se realizó un análisis de agrupamiento mediante el método de promedios ponderados (UPGMA) por muestreo y fragmento por medio del índice de similitud de Jaccard. Para el análisis y construcción del agrupamiento se utilizó el programa PRIMER 6.0 (CLARKE & WARWICK, 2001).
Con el propósito de relacionar las variables ambientales (temperatura, humedad ambiente y precipitación) con la riqueza de Reduviidae, se realizó una correlación de Spearman. Asimismo, se realizó la correlación entre la abundancia de géneros más frecuentes y las variables ambientales en el área de estudio, con el fin de determinar cuál fue la variable que incidió en la variación de la abundancia de estos insectos. Los análisis fueron realizados usando el programa PAST 1.63 (HAMMER et al., 2001).
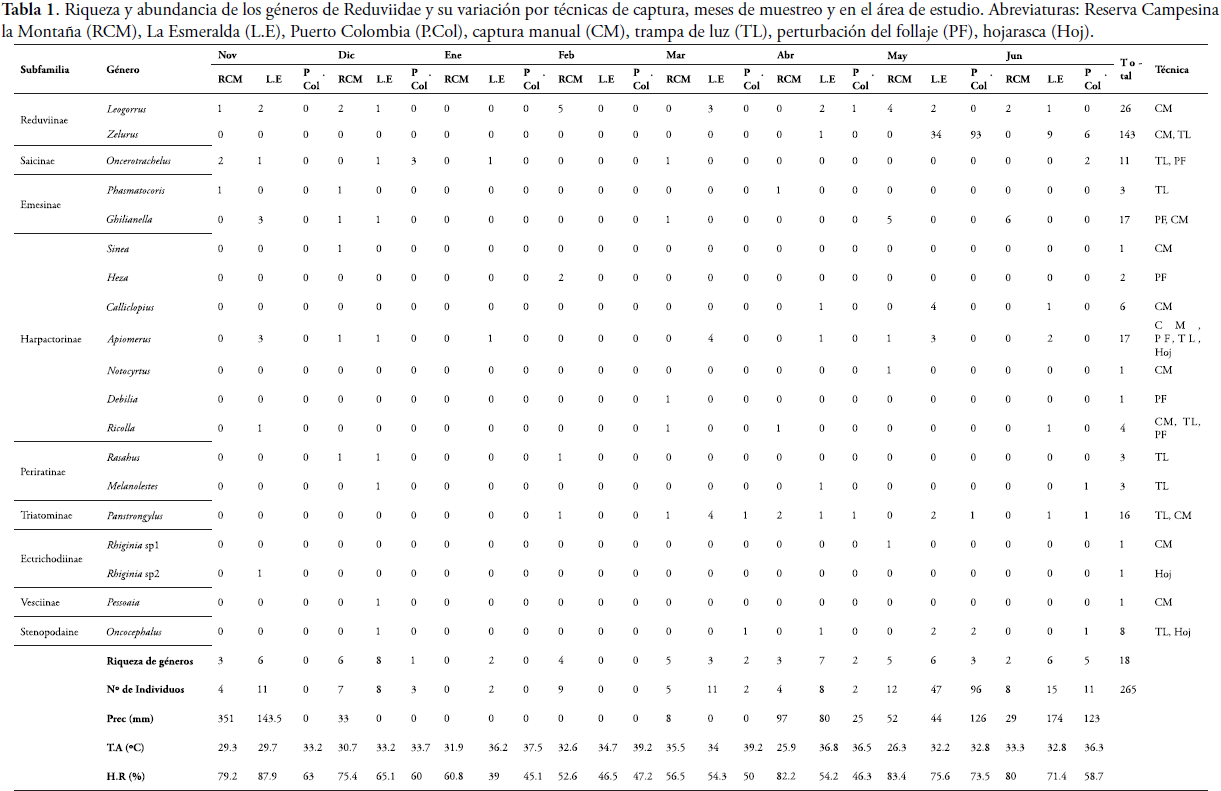
RESULTADOSEn los tres fragmentos se recolectaron un total de 265 individuos distribuidos en 18 géneros y nueve subfamilias (Tabla 1). La subfamilia más diversa fue Harpactorinae con siete géneros que representan el 37% de la riqueza; mientras que la más abundante fue Reduviinae con el 64% de la abundancia total. Teniendo en cuenta los fragmentos, en L.E se registraron 9 subfamilias y 13 géneros y en la RCM fueron capturadas 6 subfamilias con 13 géneros; mientras que en P. Col se reportaron 6 géneros pertenecientes a 5 subfamilias. El mayor número de géneros se registró durante diciembre y abril (8 y 7, respectivamente) en el fragmento L.E, mientras que mayo fue el más abundante con 34 y 96 individuos en L.E y P. Col respectivamente (Tabla 1).
El género más abundante fue Zelurus Hahn con 143 individuos, presentando su mayor valor durante mayo y junio, apareciendo únicamente en los fragmentos de L.E y P. Col. Luego le sigue Leogorrus Stål con 26 individuos presentes en los tres fragmentos y teniendo una aparición muy variable durante los meses de muestreos. Ghilianella Spinola y Apiomerus Hahn (con 17 individuos cada uno) fueron encontrados solo en la RCM y L.E. Por último, se destaca la presencia de 16 individuos del género Panstrongylus Berg, capturados en los tres fragmentos. Además, se enfatiza en la presencia de seis géneros (aportando >30 %) que se podrían calificar de 'raros'M, ya que solo fue capturado un ejemplar de cada uno, a pesar de que se llevaron a cabo muestreos intensivos en toda el área de estudio (Tabla 1).
En el caso de la variación por muestreo de los géneros más abundantes (>10%), el mayor número de individuos se registró en mayo, destacándose Zelurus con 93 individuos en P. Col y 34 en L.E; cuando los niveles de precipitación oscilaron alrededor de los 126 y 44 mm (Tabla 1), la temperatura ambiente fue de 32,8°C y 32,2°C y la humedad relativa osciló entre 73,5% y 75,6%. En el caso de Apiomerus, el mayor valor (4) se observó en L.E en marzo, cuando la precipitación fue de cero; mientras que Ghilianella presentó su mayor valor (6) en junio en la RCM, cuando la precipitación fue de 29 mm. Leogorrus, fue más abundante (5) en febrero y Panstrongylus en marzo (4) para L.E. En enero y febrero se obtuvo el menor número de individuos, lo cual puede estar relacionado con un periodo seco (precipitaciones medidas en el área = 0mm) (Tabla 1).
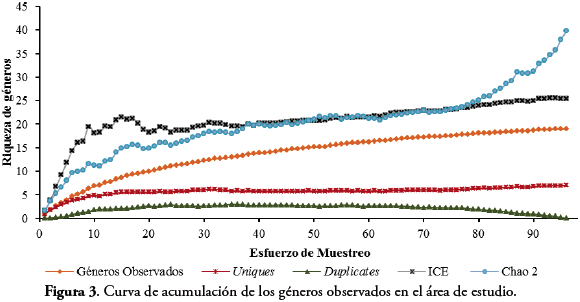
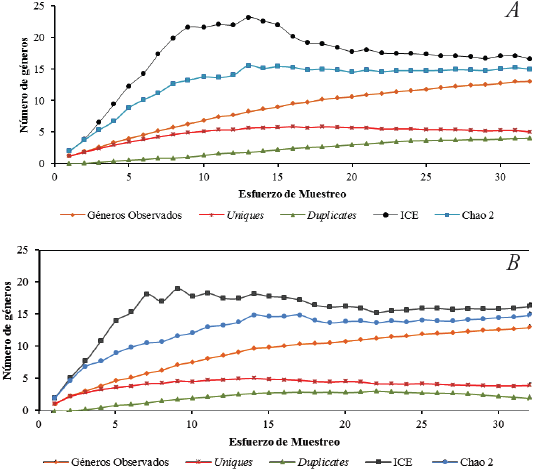
La curva de acumulación indica que los géneros de Reduviidae observados (18) se encuentran entre 76% (ICE= 25) y el 59,37% (Chao2=32) del total de géneros esperados para el área de estudio. Por su parte, la curva de géneros observados se encuentra muy pronunciada con una tendencia alejada a lograr una asíntota. En cuanto, a los géneros Uniques (7) presentan una alta cantidad para toda el área, mientras que los géneros Duplicates tienden a desaparecer en los últimos muestreos (Figura 3). En la curva de acumulación para cada uno de los fragmentos, los géneros de Reduviidae observados para la RCM (Chao2=14,94 ICE=16,58) y L.E (Chao 2=14,94 ICE=16,29) fueron de 13 cada uno. Según la estimación de la diversidad para estos dos fragmentos, se esperaría que aproximadamente tres géneros fueran adicionados en la riqueza de estas dos áreas (Fig. 4A y B) y para P.Col. se registran 6 géneros, de los cuales se esperaría un género adicional en este fragmento (Chao2=6,48 ICE=7,14) (Fig. 4C).
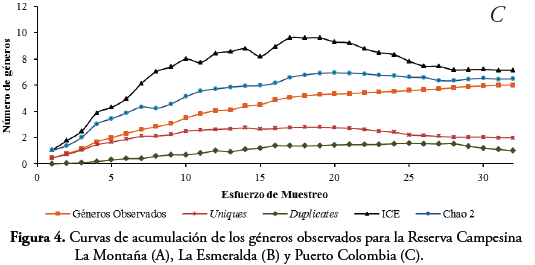
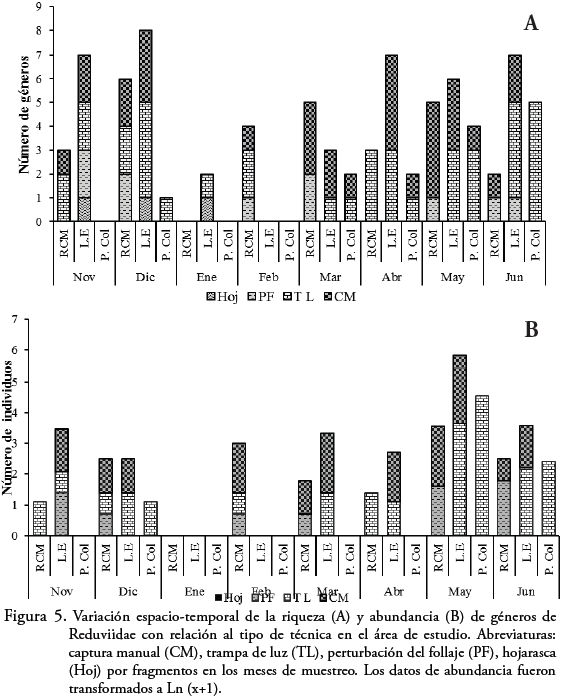
Teniendo en cuenta las técnicas de captura, Phasmatocoris Breddin, Rasahus Amyot & Serville y Melanolestes St'l fueron capturados con TL, mientras Leogorrus, uno de los géneros más abundantes solo fue capturado con trampa activa (CM) y Apiomerus con las cuatro técnicas (Tabla 1). La mayor riqueza (9 géneros) y abundancia (184 individuos) de Reduviidae se registró en las trampas de luz (TL), mientras que en manual (CM) se registraron 11 géneros y 44 individuos respectivamente (Figura 5B). La técnica de cernido de hojarasca (Hoj), fue la que presentó la menor efectividad en todos los fragmentos con tres géneros y tres individuos durante noviembre, diciembre y enero respectivamente. El uso de las cuatro técnicas fue efectivo para obtener en L.E ocho géneros en noviembre, al igual que en diciembre en L.E y en la RCM (Figura 5).
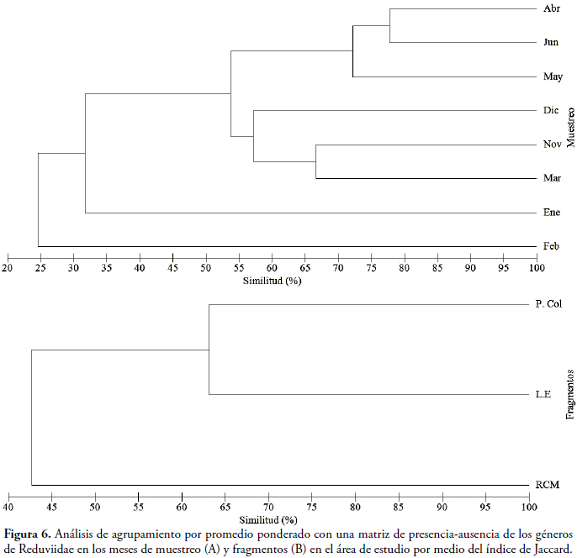
En el caso de los fragmentos de bosque, solo tres géneros son compartidos (Leogorrus, Oncerotrachelus Stål, Panstrongylus). L.E y P.Col comparten tres géneros, mientras L.E y RCM solo dos; siendo esta última la que presentó el mayor número (6) de géneros exclusivos. Por otro lado, entre los meses de muestreo, se forman cinco grandes grupos (Figura 6A). El primero se encuentra constituido por abril, mayo y junio con una similitud de 72,22%. El segundo está conformado por noviembre y marzo con el 66,67% y el tercero por diciembre con un valor inferior al 60%, muestreo en el cual empiezan a decaer los valores de precipitación en la zona (Tabla 1). Los dos últimos grupos están conformados por enero y febrero, los cuales presentan valores de similitud menores al 35%, caracterizándose por ser los meses más secos (0mm) (Figura 6A y Tabla 1). Con respecto a la variación de la composición de Reduviidae por fragmentos, se forman dos agrupamientos. Uno formado por P.Col y L.E, con una similitud del 63,16 %, mientras que el otro grupo lo conforma el fragmento de RCM (Figura 6B).
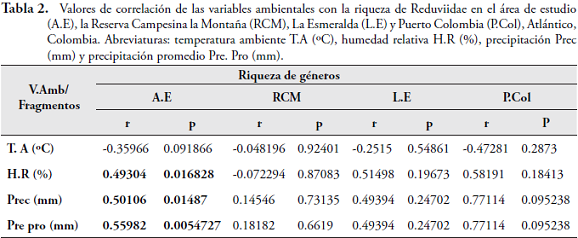
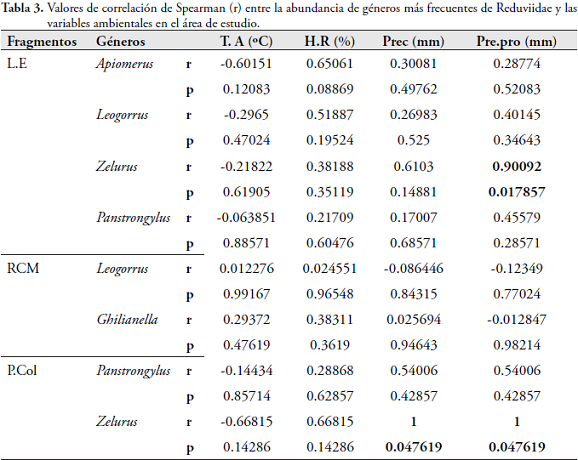
Con el análisis de correlación de Spearman, se demostró que la riqueza de géneros de Reduviidae es directamente proporcional y significativa con la humedad relativa (r=0,49304; p=0,016828), precipitación (r=0,50106; p=0,01487) y promedio (r=0,55982; p=0,0054727) (Tabla 2). Al relacionar las variables ambientales con la abundancia de los géneros más representativos en el área de estudio, se encontró que Zelurus fue el único que presentó una correlación significativa y directamente proporcional con las precipitaciones para los fragmentos de L.E (r = 0,90092; p = 0,017857) y P.Col (r = 1; p = 0,047619) (Tabla 3).
DISCUSIÓN
Teniendo en cuenta el número de subfamilias (9) de Reduviidae encontradas para el Atlántico, la composición representa el 60% con respecto al número que se encuentran para Colombia (16) (FORERO, 2004). Lo anterior representa el 43% para el Neotrópico con 21 subfamilias conocidas (FROESCHNER & KORNILEV, 1989; MALDONADO, 1990; GIL-SANTANA & COSTA, 2003; FORERO, 2004). Asimismo, el número de géneros reportados para el Atlántico, representan un 10% de los 201 géneros conocidos para el Neotrópico, mientras que para Colombia (95) es el 20% de los reportados por FORERO (2004). Estos resultados constituyen los primeros aportes al conocimiento sobre la diversidad de géneros de Reduviidae para el Atlántico en un ecosistema como el BST. Se registran para el departamento 18 nuevos reportes de géneros, de los cuales es muy poca la información que se tenía tanto de sus métodos de captura, distribución y comentarios de su biología en campo; de los cuales algunos han sido reportados para la región Caribe (FORERO, 2006).
La mayor riqueza de Harpactorinae se puede atribuir a que sus individuos se encuentran habitualmente en diversos microhábitats tales como vegetación baja, en hojarasca o árboles caídos y durante el día en lugares soleados (BRAILOVSKY et al., 2007); lo que facilita de cierta manera su búsqueda y captura en este tipo de hábitats que son muy comunes en el área de estudio; además de ser la subfamilia más grande de Reduviidae con 51 géneros para la región neotropical (MALDONADO, 1990, FORERO et al., 2008), que se encuentra constituida principalmente de depredadores generalistas (READIO, 1927, LOUIS, 1974). La alta abundancia de la subfamilia Reduviinae se atribuye a que es considerada una de las más heterogéneas debido a su distribución (MELO, 2007). Adicional a lo anterior, este resultado se debe a que Zelurus presentó un incremento del número de individuos en solo dos muestreos, lo que aportó de manera significativa en la abundancia de Reduviidae que se tenía hasta el momento. La estrecha relación de la emergencia de los adultos con la llegada de las primeras lluvias en L.E y P.Col, estarían explicando la mayor abundancia de Zelurus en mayo (NANNI et al., 2011). Además, la abundancia de este género parece estar fuertemente influenciada por fuentes de luz durante la noche, lo cual concuerda con los resultados obtenidos por PROUS (2005). En el caso de aquellos géneros capturados con pocos individuos, se puede atribuir a que se hace necesaria una búsqueda más exhaustiva en algunos de los microhábitats más específicos que atienden a la biología de estos géneros (FORERO, com. pers).
La mayor riqueza en diciembre puede estar relacionada con los cambios en la precipitación y la humedad relativa en el área de estudio. Durante este mes se pasó de un periodo de altas lluvias a uno seco, cuando el bosque presentó un cambio brusco en la vegetación, perdiendo gran parte de su follaje y un aumento en la hojarasca. Lo anterior puede constituir un aumento en ciertos microhábitats preferidos por algunos géneros de Reduviidae, lo cual también se relaciona con un aumento de presas detritófagas. Caso contrario se evidenció en abril, en donde a pesar de que se observó nuevamente un aumento en la riqueza con la llegada de las primeras lluvias, se presentó en los tres fragmentos un rebrote de la vegetación y un incremento en la aparición de fitófagos, los cuales son un recurso alimenticio para este grupo de insectos.
La presencia de Apiomerus durante todos los muestreos se puede atribuir a que tenga varias generaciones en el año sin importar las condiciones imperantes (ambientales o disponibilidad de presas). Por otro lado, la mayor abundancia de este género en la L.E, está relacionada con la presencia de árboles de Mangifera indica, que proporcionan una resina utilizada por estos Reduviidae para capturar las presas. Esto coincide con lo propuesto por MILLER (1956), BÉRENGER & PLUOT-SIGWALT (1997), quienes describen que este género emplea la resina de las plantas como método de captura de las presas, aplicándosela al cuerpo como trampa adhesiva (ADIS, 1984; GIL-SANTANA & FORERO, 2010). Adicional a lo anterior, este género usa la resina de las plantas para el cuidado parental, cubriendo sus huevos y protegiéndolos contra la depredación y la desecación, además de ayudarle a las ninfas de primeros instares en la captura de sus presas (EISNER, 1988; CHOE & MOHO, 2007; FORERO et al., 2011). Este comportamiento, muy similar dentro de la tribu Apiomerini, fue observado en individuos del género Callioclopus, registrándose una hembra con cinco ninfas de diferentes estadios escondidos en un árbol de M. indica. La hembra mostraba un comportamiento de defensa al pie del árbol al momento de la captura (obs. pers.). Esto se complementa con lo reportado por BÉRENGER & PLUOT-SIGWALT (1997), en los llanos orientales de Colombia, quienes encontraron ninfas de este género cubiertas de resina y por FORERO (2004), quien describe por primera vez el uso de resinas para este grupo de hemípteros.
A manera de observación de campo, se registró un Apiomerus sp depredando un escarabajo de la familia Tenebrionidae en la base de un árbol de Hura crepitans. Lo anterior coincide con lo descrito por GIL-SANTANA (2002) y MARQUES et al. (2003), quienes reportan a Apiomerus spp. capturando insectos de diferentes órdenes, incluyendo a Coleoptera e Hymenoptera (abejas) (WEAVER et al., 1975; GONZALES-BUSTAMANTE, 1994; SILVA & GIL-SANTANA, 2004; BERNIKER et al., 2011); razón por la cual se comporta como generalista en el área de estudio.
Con respecto a la especie P. geniculatus, reportada para el Atlántico, se destaca su alta presencia y distribución en los tres fragmentos estudiados; lo cual coincide con lo descrito para otros sitios del departamento por D'ALESSANDRO et al. (1981; 1985) y BARRETO & BARRETO (1984). La especie P. geniculatus se ha registrado para los municipios de Luruaco, Piojó, Tubará, Puerto Colombia, Barranquilla y Soledad (MAESTRE & EYES, 2012). En el presente trabajo se registra la especie para el municipio de Juan de Acosta (RCM) y se amplía la distribución para la vereda La Esmeralda, en Tubará; así como para Puerto Colombia (km 6, vía puerto Colombia). Esta información es importante al momento de realizar seguimientos de distribución de vectores en el departamento, debido a que se han dejado de lado las investigaciones de la dinámica poblacional de vectores silvestres enfocándose fuertemente en los estudios en la parte domiciliaria (MOJICA et al., 2003; PARRA et al., 2004). Para esta especie de hábitos netamente silvestres se hace relevante su estudio tanto en ambientes domiciliarios como silvestres (TONN et al., 1976; LORENZO et al., 1998; VÁSQUEZ et al., 2007; ANGULO & ESTEBAN, 2011); debido a su importancia como vector transmisor de la enfermedad de Chagas y su exitosa adaptación a una amplia variedad de biomas y ecosistemas (PATTERSON et al., 2009).
Además del monitoreo del vector, se hace necesario conocer el estado actual de los fragmentos de BST, ya que la reducción a la cual están siendo sometidos, traen consigo cambios como la domiciliación de especies que antes eran netamente silvestres como P. geniculatus. Según ANGULO (2000), esta especie visita frecuentemente el domicilio aledaño a áreas donde aún existen relictos de bosque primario o secundario; debido a la disminución o cambios del área del bosque por la ampliación de la zona agrícola o urbana. Lo anterior conlleva al alejamiento de los mamíferos y aves silvestres de los cuales se alimentan especies silvestres como P. geniculatus, determinando cambios en su biotopo, causando que estas busquen vertebrados domésticos en el interior de las viviendas e infectan en forma esporádica a la población humana (VEGA et al., 2006).
Los resultados de la curva de acumulación en el área de estudio indican que se hace necesario realizar más muestreos, con el fin de tener una riqueza de géneros más cercana a la real y poder establecer con propiedad un número más representativo para el Atlántico. Lo correspondiente a la trayectoria de la curva de acumulación de géneros Uniques, indica una posible estacionalidad de estos géneros. Es probable que si se realizan más muestreos, con un mayor esfuerzo en ciertas épocas del año, se podría tener información más precisa sobre la biología de aquellos Reduviidae con bajo número de individuos. En lo que se refiere a la curva de acumulación para los tres fragmentos, aún no se obtienen el total de géneros posibles para el área y probablemente al invertir más días en cada muestreo y mejor resolución taxonómica podría obtenerse mejores resultados. Esto se sustenta también en la tendencia de la curva de géneros Uniques, la cual no se estabilizó, influyendo en los valores estimados. Es posible que estos géneros Uniques se presenten por la falta de técnicas más específicas para su captura que estén de acorde con su biología. Además, probablemente la población de estos individuos se encuentre en una baja densidad, razón por lo cual se hace necesario un monitoreo más exhaustivo para estos taxones.
La mayor riqueza con CM se puede explicar debido a la biología que exhiben muchos de los géneros de esta familia. Estos insectos presentan hábitos como estar siempre cubiertos por tierra (PROUS, 2005), debajo de los troncos podridos y de la corteza de los árboles o entre las grietas de las rocas (BRAILOVSKY et al., 2007), razón por la cual con la técnica mencionada se logra revisar este tipo de microhábitats. De esta manera se logra reconocer algunos de los sitios preferidos por esta familia, logrando obtener un mayor número de géneros con esta técnica. La mayor abundancia con trampa de luz, se debe a que algunos de los géneros presentan foto tactismo positivo y son fácilmente atraídos hacia el estímulo sin limitar su número, mientras que otros simplemente se ven atraídos por la presencia de potenciales presas. La TL por ser una trampa que se deja por largas horas con atrayente (luz), logra ser mucho más efectiva que una trampa activa (CM), la cual en la mayoría de los casos depende directamente del recolector, lo que podría de cierta manera limitar su eficiencia. En lo que concierne a la alta abundancia con la técnica de TL, puede estar relacionado con la gran cantidad de individuos capturados de Zelurus, el cual probablemente tiene hábitos nocturnos o crepusculares.
Cabe resaltar que sin la complementariedad de las técnicas no se hubiese podido capturar a Phasmatocoris, Rasahus y Melanolestes, que solo fueron atraídos por trampa de luz. En el caso de Leogorrus, solo fue capturado manualmente entre la corteza o la base de los árboles, debajo de troncos podridos o grietas en el suelo, por esta razón esta técnica es eficiente para este taxón; mientras que Apiomerus, fue recolectado con la utilización de las cuatro técnicas de captura. Lo anterior demuestra que para tener un valor más exacto sobre la riqueza y composición de Reduviidae, se hace necesario utilizar el mayor número de técnicas y de esta manera enriquecer el inventario de estos insectos y su dinámica en el bosque seco. Con respecto al uso de la técnica perturbación de follaje, se hizo más efectiva en los muestreos en donde la vegetación arbustiva presente en los fragmentos era más abundante, a diferencia de los meses en los cuales el bosque perdía gran parte de la vegetación, característica propia de los BST. La captura de Phasmatocoris con trampa de luz no concuerda con lo descrito por FORERO (2006); quien ha capturado ejemplares manualmente, con pitfall y trampas Malaise; razón por lo cual esta información complementa los datos sobre el uso de técnicas que se deben usar para el estudio de este género en el bosque seco. En cuanto a los géneros Rasahus y Melanolestes, no se encontraron reportes de ser capturados únicamente con trampas de luz. Los estudios realizados sobre estos dos géneros se basan en revisiones, taxonomía, sistemática y descripción de nuevas especies (MCPHERSON et al., 1991; COSCARÓN & CARPINTERO, 1993; COSCARÓN et al., 1994; MORRONE & COSCARÓN, 1996, 1998; ZHANG & WEIRAUCH, 2011), dejando de lado el estudio de la biología de estos géneros.
La variación espacio-temporal observada en la composición de géneros de Reduviidae, en la que posiblemente se pueden encontrar géneros específicos para época seca y de lluvia, puede estar respondiendo a características propias de cada género o a la biología de estos. Sin embargo, se observó que los meses donde se iniciaron las primeras lluvias (marzo-junio) fue donde apareció el mayor número de géneros compartidos. Probablemente esto se debe a que la presencia y abundancia está estrechamente sincronizado con el aumento en los valores de precipitación durante la temporada de lluvias (MAASS et al., 2002). Adicionalmente, se puede asumir que con la llegada de las lluvias se presenta un aumento en la cantidad de insectos fitófagos que les sirven como alimento a los Reduviidae; así como una mayor proporción de refugios necesarios para su existencia y condiciones microclimáticas que favorecen la emergencia de estos chinches depredadores.
El menor número de géneros de Reduviidae compartidos entre la RCM y P.Col. puede estar relacionado con que existen diferencias marcadas entre estos dos fragmentos como la distancia. La RCM se encuentra ubicada en el centro del Atlántico, el cual presenta relieve ondulado con pendientes inclinadas, lo que conlleva al poco acceso de asentamientos humanos y por lo cual el fragmento mantiene parte de su condición original. En el caso de P.Col., se encuentra en el noroccidente del departamento y presenta un terreno plano poco ondulado, con un acelerado proceso de urbanización, lo que implica la fragmentación de los parches de bosques ubicados en esta área, pérdida de conectividad y cambios bruscos en sus condiciones ambientales. Además, la fenología del bosque es distinta para cada fragmento, ya que en la RCM no se da la pérdida completa del follaje durante los meses del año y se puede observar árboles que mantienen hojas aun en la época seca. También hay que mencionar que esta se caracteriza por tener árboles con dosel >10m, característica que beneficia a los Reduviidae y les proporciona mayor disponibilidad de refugio y microhábitats, independientemente de la época. En el caso de P.Col, la vegetación pierde totalmente su follaje durante la época seca, lo que podría representar una característica adversa para que algunos Reduviidae colonicen este lugar. Además, existe un deterioro de la calidad del bosque debido a la frecuente extracción de madera para la elaboración de carbón, así como muchos caminos que son frecuentemente utilizados por los colonos. Según KLEIN (1989) y GIBBS & STANTON (2001), las alteraciones en el microclima después de la fragmentación de un hábitat debido a la disminución del área y el aumento de los bordes, permite que la zona reciba mayor radiación solar y la aridez se incremente, lo que proporciona condiciones adversas para el desarrollo de estos insectos. Esto sugiere una simplificación de la complejidad estructural del fragmento de P.Col; mientras que RCM es uno de los pocos fragmentos que podría ser considerado área protegida. Estas diferencias implican variaciones características tanto ambientales como en la composición vegetal entre los fragmentos; lo que de una u otra manera podría explicar las diferencias en la diversidad de Reduviidae entre las dos localidades.
En el caso de los géneros Leogorrus, Oncerotrachelus y Panstrongylus, los cuales se encontraron en los tres fragmentos, posiblemente se estén comportando como géneros euritípicos, las cuales no responden a un patrón de distribución específico, sino que se han adaptado a distintos hábitats, siendo capaces de tolerar las variaciones microclimáticas en cada fragmento objeto de estudio.
Durante abril, mayo y junio el incremento en el número de individuos recolectados coincide con la llegada de las primeras lluvias a la zona. Esto también coincide con el aumento en la abundancia de Zelurus, donde se observó que después de seis meses de no aparecer, se reporta por primera vez en mayo con muchos individuos. Esto puede estar fuertemente relacionado con la estacionalidad bien marcada que presenta este género con la llegada de las lluvias y la emergencia de sus adultos, según lo expresado por PROUS (2005). Este mismo comportamiento también se observó con Ghilianella, el cual fue observado entre el follaje y arbustos, en contraste con los meses secos (enero y febrero), cuando no fue posible su captura. En general, la mayoría de los individuos sincronizan sus apariciones en épocas donde se presenta un mayor aprovechamiento de recursos (presas) y garantizan su permanencia, alimento y éxito reproductivo.
Con respecto a la similitud mostrada entre noviembre y marzo, su relación se debe principalmente a que son meses en los cuales hay un periodo de transición entre épocas. Esto proporciona microhábitats para refugio y alimentación para muchos insectos detritófagos; recurso alimenticio que es aprovechado por algunos de los géneros de Reduviidae. En abril, el paso de una época seca a lluvias marca otro punto de cambio en los fragmentos, cuando el bosque seco reverdece y presenta un aumento en la oferta de presas fitófagas. Durante enero y febrero (meses secos), los artrópodos disminuyen su abundancia, razón por la cual es muy baja la probabilidad de alimento por escasez de fitófagos, ya que se observa pérdida parcial del follaje, una de las principales adaptaciones fisiológicas de las plantas ante el déficit de agua en BST (IAVH, 1998). La alta similitud entre los fragmentos de P.Col y L.E puede estar relacionada con alguna semejanza en la pérdida de la cobertura y estructura de la vegetación entre estos dos sitios, a diferencia de lo obtenido con RCM; donde se observa una estratificación vegetal, con una cobertura arbórea más densa en comparación con los otros dos fragmentos, lo cual proporciona una gran variedad de microhábitats. Esto demuestra una variación espacio-temporal de la composición de Reduviidae en el Atlántico, expresando probablemente una característica de algunos géneros de la familia a ser dependientes de regímenes climáticos y producción de recursos vegetales en el BST.
La incidencia de la precipitación y la humedad relativa en la riqueza de Reduviidae demuestra que este factor juega un papel importante en el ciclo biológico y la estacionalidad de algunos géneros. Esto concuerda con lo sugerido por BLINN & RUNCK (1989) y AMBROSE (1999), quienes explican que la disponibilidad de agua es una de las condiciones que necesitan los huevos de heterópteros depredadores para sobrevivir; determinándose como un factor importante en la dinámica ecológica de estos insectos en los fragmentos de bosque seco. Por otro lado, en este estudio se demostró la importancia que tienen las precipitaciones sobre algunos de los géneros más representativos como Zelurus, debido a que dentro de este tipo de bosques existen especies que tienen sincronizada su emergencia y periodo de actividad con las lluvias en la zona; razón por la cual se hace necesario tener datos experimentales en campo para demostrar cómo incide esta variable en los Reduviidae del BST.
AGRADECIMIENTOSA la comunidad campesina de la vereda Los Torombolos (RCM) por facilitar la estancia y permisos en este lugar durante la realización del trabajo de campo, en especial a los señores Santiago y Rafael Conrado y demás familiares, por acogernos en su hogar y colaborarnos con la alimentación. Al señor Sigifredo, su esposa y resto de familiares por siempre ser una gran compañía, apoyo y acogernos durante todo los muestreos en Puerto Colombia. A la señora 'Alejandra' por abrirnos un campito dentro de su hogar durante los muestreos en Corrales de San Luis. Al Dr. Igor Dimitri Forero por su colaboración en la identificación de los Reduviidae y por abrirnos las puertas de su laboratorio en la Universidad Javeriana (Bogotá-Colombia). A los miembros del semillero de investigación NEOPTERA del programa de biología, por su colaboración en el trabajo de campo y la Facultad de Ciencias Básicas de la Universidad del Atlántico por facilitar los permisos y las instalaciones del Laboratorio de Zoología para el procesamiento e identificación de las muestras.
ADIS, J., 1984.- Eco-entomological observations from the Amazon. V. Feeding habits of Neotropical "bee killers" and "tresin bugs" (Apiomerinae: Reduviidae: Hemiptera). Revista de Biología Tropical, 32: 151-153. [ Links ]
ANGULO, V.M., 2000.- Aspectos ecológicos de la enfermedad de Chagas en el oriente de Colombia. Mvz-cordoba, 5(1): 64-68. [ Links ]
ANGULO, V.M. & ESTEBAN, L., 2011.- Nueva trampa para la captura de triatóminos en hábitats silvestres y peri-domésticos. Biomédica, 31: 264-8. [ Links ]
BARRAZA, J., MONTES, J., MARTÍNEZ, N. & DELOYA. C., 2010.- Ensamblaje de escarabajos coprófagos (Scarabaeidae: Scarabaeinae) del Bosque Tropical Seco, Bahía Concha, Santa Marta, Colombia. Revista Colombia de Entomología, 36 (2): 285-291. [ Links ]
BARRETO, M. & BARRETO, P., 1984.- Aves y pitos en Colombia. Cespedesia, 13: 93-97. [ Links ]
BÉRENGER, J. & PLUOT-SIGWALT, D., 1997.- Relations privilegiées de certains Heteroptera Reduviidae prédaterus avec les végetaux. Premier cas connu d'un Haprpactorinae phytophage. Comptes Rendus Académie des Sciences Paris, Sciences de la vie, 320: 1007- 1012. [ Links ]
BERNIKER, L., SZERLIP, S., FORERO, D. & WEIRAUCH, C., 2011.- Revision of the crassipes and pictipes species groups of Apiomerus Hahn (Hemiptera: Reduviidae: Harpactorinae). Zootaxa, 2949: 1-113. [ Links ]
BLINN, D.W. & RUNCK, C., 1989.- Substratum requirements for oviposition, seasonal egg densities, and conditions for egg eclosion in Ranatra montezuma (Heteroptera: Nepidae). Annals of the Entomological Society of America, 82: 707-711 [ Links ]
BRAILOVSKY, H., MARIÑO, R. & BARRERA, E., 2007.- Cinco especies nuevas de Pselliopus (Hemiptera: Heteroptera: Reduviidae: Harpactorinae: Harpactorini) para México. Revista Mexicana de Biodiversidad, 78: 85- 98. [ Links ]
CANO, E.B. & SCHUSTER. J.C., 2007.- International commission on tropical biology and natural resources - Beetles as Indicators for Forest Conservation in Central America. Tropical Biology and Conservation Management. [ Links ]
CANTILLO-BARRAZA, O., GÓMEZ-PALACIO, A., SALAZAR, D., MEJÍA-JARAMILLO, A., CALLE, J. & TRIANA, O., 2010.- Distribución geográfica y ecoepidemiológica de la fauna de triatominos (Reduviidae: Triatominae) en la Isla Margarita del departamento de Bolívar, Colombia. Biomédica, 30: 382-389. [ Links ]
CASTRO-HUERTAS, A., 2010.- Sistemática del género Acanthischium Amyot & Serville, 1843 (Hemiptera: Reduviidae: Harpactorinae). Tesis de Maestría no publicada. Universidad Nacional, Bogotá D.C. Colombia. [ Links ]
CEBALLOS, G., 1995.- Vertebrate diversity, ecology, and conservation in Neotropical dry forest. En: Bullock, S., E. Medina & H. A. Mooney (Eds). Tropical deciduous Forest Ecosystem. Cambridge Univ. Press, Cambridge. 195-222. [ Links ]
CHAVERRA-RODRÍGUEZ, D., FORERO, D., JARAMILLO, N. & TRIANA, O., 2010.- New record and ecological notes of Phimophorus spissicornis (Hemiptera: Heteroptera: Reduviidae: Phimophorinae) in Colombia. Revista Colombiana de Entomología, 36 (1): 176-178. [ Links ]
CHOE, D.H. & MOHO, K.R., 2007.- Use of plant resin by a bee assassin bug, Apiomerus flaviventris (Hemiptera: Reduviidae). Annals of the Entomological Society of America, 100 (2): 320-326. [ Links ]
CLARKE, K.R. & WARWICK, R.M., 2001.- Change in marine communities: An approach to statistical analysis and interpretation. Natural Environment Research Council, UK. 144 p. [ Links ]
COLWELL, R.K., 2009.- ESTIMATES: Statistical Estimation of Species Richness and Shared Species from Samples, Version 8.2. Available at http://viceroy.eeb.uconn.edu/estimates [ Links ]
COLWELL, R.K. & CODDINGTON, J., 1994.- Estimating terrestrial biodiversity through extrapolation. Phil. Trans. R. Soc. Lond, 345: 101-118. [ Links ]
COSCARÓN, M.C. & CARPINTERO, D., 1993.- Revision of the genus Melanolestes Stål (Heteroptera: Reduviidae: Peiratinae) Ent. Scand, 24 (4): 361-381. [ Links ]
COSCARÓN, M.C., DÍAZ, N.B. & POVEL, G.D.E., 1994.- Taxonomic congruence between external morphology and male and female genitalia characters of members of Rasahus Amyot & Serville (Heteroptera: Reduviidae: Peiratinae). Zool. Med. Leiden, 68(10): 97-108. [ Links ]
DÍAZ, J.M., 2006.- Bosque seco tropical en Colombia. Cali: Banco de Occidente. [Última consulta: 27 Noviembre 2015]. Disponible en: http://www.imeditores.com/banocc/seco/creditos.htm [ Links ]
DIRZO, R., YOUNG, H.S., MOONEY, H.A. & CEBALLOS, G., 2011.- Seasonally Dry Tropical Forests ecology and conservation. Island Press, Washington, DC 2009, USA. [ Links ]
D' ALESSANDRO, A, BARRETO, P. & THOMAS, M., 1981.- Nuevos registros de triatominos domiciliarios y extradomiciliarios en Colombia. Colombia Med, 12:75-85. [ Links ]
D'ALESSANDRO, A. & BARRETO, P., 1985.- Colombia. En: Carcavallo, R.U, J.E. Ravinovich, R.J. Tonn. (Eds.) Factores biológicos y ecológicos de la enfermedad de Chagas. Tomo II. Buenos Aires: Centro Panamericano de Ecología Humana y Salud, Organización Panamericana de la Salud/Organización Mundial de la Salud. 377-99. [ Links ]
EISNER, T., 1988.- Insekten als fursorgliche Eltern. Verhandlungen der Deutschen Zoologischen Gesellschaft, 81: 9-17. [ Links ]
FORERO, D., 2004.- Capítulo 5. Diagnóstico de los géneros neotropicales de la familia Reduviidae (Hemiptera: Heteroptera) y su distribución en Colombia (excepto Harpactorinae). Pp 129-275. En: FERNÁNDEZ, F., ANDRADE, M. & AMAT, G. (Eds.) Insectos de Colombia. Universidad Nacional de Colombia (sede Bogotá) Facultad de Ciencias, Instituto de Investigación de Recursos Biológicos, Humboldt. 3: 602p. [ Links ]
FORERO, D., 2006.- New records of Reduviidae (Hemiptera: Heteroptera) from Colombia and other Neotropical countries. Zootaxa, 1107: 1-47. [ Links ]
FORERO, D., 2008.- The systematics of the Hemiptera. Revista Colombiana de Entomología, 34 (1): 1-21. [ Links ]
FORERO, D., GIL-SANTANA, H.R. & VAN DOESBURG, P.H., 2008.- Redescription of the Neotropical genus Aristathlus (Heteroptera, Reduviidae, Harpactorinae), p. 85-103. In: GROZEVA, S. & SIMOV, N., (Eds.). Advances in Heteroptera Research, Festschrift in Honour of 80th Anniversary of Michail Josifov. Sofia-Moscow, Pensoft Publishers, 417p. [ Links ]
FORERO, D., CHOE, D.H. & WEIRAUCH, C., 2011.- Resin Gathering in Neotropical Resin Bugs (Insecta: Hemiptera: Reduviidae): Functional and Comparative Morphology. Journal of Morphology, 272 (2): 204 - 229. [ Links ]
FROESCHNER, R.C. & KORMILEV, N.A., 1989.- Phymatidae or ambush bugs of the world: a synonymic list with keys to species, except Lophoscutus and Phymata (Hemiptera). Entomography, 6:1- 76. [ Links ]
GARCÍA, H., CORZO, G., ISAACS, P. & ETTER, A., 2014.- Distribución y estado actual de los remanentes del bioma de bosque seco tropical en Colombia: insumos para su gestión. En PIZANO, C. & GARCÍA, H (Editores). El bosque seco tropical en Colombia, pp. 229-251 I. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Bogotá-Colombia. [ Links ]
GIBBS, J. & STANTON, E., 2001. Habitat fragmentation and arthropod community change: carrion beetles, phoretic mites and flies. Ecological Applications, 11: 79-85. [ Links ]
GIL-SANTANA, H.R., 2002.- Predação de Lagria villosa Fabricius, 1783 (Coleoptera: Lagriidae) por Apiomerus nigrilobus Stål, 1872 (Hemiptera: Reduviidae: Apiomerinae) em Cabo Frio, Estado do Rio de Janeiro, Brasil. Entomología y Vectores, Rio de Janeiro. 9(2): 201-208. [ Links ]
GIL-SANTANA, H.R. & COSTA, L.A., 2003.- A new genus and species of Peiratinae from Nova Friburgo, Rio de Janeiro, Brazil (Hemiptera, Heteroptera, Reduviidae). Revista Brasileira de Zoologia, 20: 3-8. [ Links ]
GIL-SANTANA, H.R. & FORERO, D., 2010.- Taxonomical and biological notes on Neotropical Apiomerini (Hemiptera: Heteroptera: Reduviidae: Harpactorinae). Zootaxa, 2331: 57-68. [ Links ]
GONZALES-BUSTAMANTE, L.E., 1994.- Apiomerus sp. (Hemiptera-Reduviidae) mimic of Trigona trinidadensis (Hymenoptera-Apidae). Revista Peruana de Entomología, 37: 115-116. [ Links ]
GUHL, F., AGUILERA, G., PINTO, N. & VERGARA, D., 2007.- Actualización de la distribución geográfica y ecoepidemiología de la fauna de triatominos (Reduviidae: Triatominae) en Colombia. Biomédica, 27(1): 143- 162. [ Links ]
GUHL, F., PINTO, N. & AGUILERA, G., 2009.- Sylvatic triatominae: a new challenge in vector control transmission. Memorias Instituto Oswaldo Cruz, Rio de Janeiro, 104 (1): 71-75. [ Links ]
HAMMER, Ø., HARPER, D.A.T. & RYAN, P.D., 2001.- PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4 (1). Disponible en: http://palaeo-electronica.org/2001_1/past/issue1_01.htm [ Links ]
HIJMANS, R.J., CAMERON, S.E. & PARRA, J.L., 2004.- Worldclim, versión 1.3. Disponible en: http://www.worldclim.org/. [ Links ]
INSTITUTO GEOGRÁFICO AGUSTÍN CODAZZI, (IGAC)., 1994.- Atlántico: Características geográficas. Bogotá, Colombia. 157 pp. [ Links ]
INSTITUTO ALEXANDER VON HUMBOLDT, (IAVH)., 1998.- El Bosque seco Tropical (Bs-T) en Colombia. Programa de Inventario de la Biodiversidad. Grupo de Exploraciones y Monitoreo Ambiental, GEMA. 1-74. [ Links ]
KIM, D.S. & RIEDL, H., 2005. Effect of temperature on development and fecundity of the predaceous plant bug Deraeocoris brevis reared on Ephestia kuehniella eggs. BioControl, 50: 881-897. [ Links ]
KLEIN, B., 1989.- Effects of forest fragmentation on dung and carrion beetles communities in Central Amazonia. Ecology, 70: 1715-1725. [ Links ]
LENT, H. & WYGODZINSKY, P., 1979.- Revision of the Triatominae (Hemiptera: Reduviidae) and their significance as vector of Chagas disease. Bulletin of the American Museum of Natural History, 163(3): 123-520. [ Links ]
LORENZO, M.G., REISENMAN, C.E. & LAZZARI, C.R., 1998.- Triatoma infestans can be captured under natural climatic conditions using yeast-baited traps. Acta Tropica, 70: 277-284. [ Links ]
LOUIS, D., 1974.- Biology of Reduviidae of cocoa farms in Ghana. Am. Midi. Nat. 91: 68-89. [ Links ]
MAASS, J.M., JARAMILLO, V., MARTÍNEZ-YRÍZAR, A., GARCÍA-OLIVA, F., PÉREZ-JIMÉNEZ, L.A. & SARUKHÁN, J., 2002.- Aspectos funcionales del ecosistema de selva baja caducifolia en Chamela, Jalisco En: NOGUERA, F.A., VEGA, J.H., GARCÍA-ALDRETE, A.N. & QUESADA, M., (eds.). Historia Natural de Chamela, pp. 525-542, Instituto de Biología, UNAM, México, D.F., México. [ Links ]
MAESTRE, R. & EYES, M., 2012.- Actualización de la presencia y distribución de triatominos en el departamento del Atlántico-Colombia: 2003-2010. Boletín de Malariología y Salud Ambiental, 2(1):125-128. [ Links ]
MALDONADO, C.J., 1990.- Systematic catalogue of the Reduviidae of the World. Caribbean Journal of Science. Special publication, University of Puerto Rico, Mayagüez, Puerto Rico. 1: 694 pp. [ Links ]
MARTÍNEZ, N., ACOSTA, J. & FRANZ, N., 2009.- Structure of the beetle fauna (Insecta: Coleoptera) in forest remnants of western Puerto Rico. J. Agric. Univ. P.R, 93(1-2): 83-100. [ Links ]
MARQUES, O.M., GIL-SANTANA, H.R., MAGALHÃES, A.C.A. & CARVALHO, A.A.L., 2003. Predação de Apiomerus lanipes (Fabricius, 1803) (Hemiptera: Reduviidae) sobre Apis mellifera (Linnaeus, 1758) (Hymenoptera: Apidae), no Estado da Bahia, Brasil. Entomología y Vectores, 10: 419-429. [ Links ]
MARTÍNEZ, N., SALCEDO, G., SIERRA, K. & BARRAZA, J., 2012.- Escarabajos coprófagos (Coleoptera: Scarabaeinae) asociados a excrementos de mamíferos en un fragmento de bosque seco tropical en el Departamento del Atlántico, Colombia. Ecología Austral, 22: 203-210. [ Links ]
MCPHERSON, J.E., KEFFER, S.L. & TAYLOR, S.J., 1991.- Taxonomic status of Melanolestes picipes and M. abdominalis (Heteroptera: Reduviidae). Florida Entomologist, 47(3): 397-403. [ Links ]
MELO, M.C., 2007.- Revision of the neotropical genus Leogorrus Stal (Hemiptera : Reduviidae). Insect Systematics & Evolution, 38: 51-92. [ Links ]
MILES, L., NEWTON, A.C., DEFRIES, R., RAVILIOUS, C., MAY, I., BLYTH, S., KAPOS, V. & GORDON, J.E., 2006.- A global overview of the conservation status of tropical dry forests. Journal of Biogeography, 491-505. [ Links ]
MILLER, N.C.E., 1956.- The biology of the Heteroptera. London. Leonard Hill, p. 162 [ Links ]
MOJICA, M.T., CUERVO, L.A., ARIZA, K., CHACÓN, E., CHACÓN, R., DIB, J.C. & GUHL, F., 2003.- Distribución de Triatoma maculata e infestación domiciliaria en Santa Marta, Colombia. Biomédica. Instituto Nacional de Salud, 23: suplemento No.1. [ Links ]
MOLINA, J., GUALDRÓN, L.E., BROCHERO, H.L., OLANO, V.A., BARRIOS, D. & GUHL, F., 2000.- Distribución actual e importancia epidemiológica de las especies de triatominos (Reduviidae: Triatominae) en Colombia. Biomédica, 20: 344-360. [ Links ]
MORRONE, J.J. & COSCARÓN, M.C., 1996.- Distributional patterns of the American Peiratinae (Heteroptera: Reduviidae). Zool. Med. Leiden, 70(1): 1-15. [ Links ]
MORRONE, J.J. & COSCARÓN, M.C., 1998.- Cladistics and biogeography of the assassin bug genus Rasahus Amyot & Serville (Heteroptera: Reduviidae: Peiratinae). Zool. Med. Leiden, 72(6): 73-87. [ Links ]
NANNI, A.S., MAGNANO, A.L. & CARPINTERO, D.L., 2011.- Los Heteroptera (Insecta: Hemiptera) edáficos del INTA Delta del Paraná (Partido de Campana, Buenos Aires): Variación espacial y temporal del elenco de las especies en distintos usos de la tierra. Rev. Mus. Argent. Cienc. Nat, 13(1): 27-39. [ Links ]
PANZA, E., TORRES, M., DE LA RODA, P., BERDUGO, I., TORRES, A., DE LA ROSA, P., PASO, J. & ARENILLA, M., 2001.- Esquema de ordenamiento territorial municipio de Juan de Acosta. Alcaldía Municipal de Juan de Acosta 2001-2009. Juan de Acosta. Departamento del Atlántico. República de Colombia [ Links ]
PARRA, J.G., RESTREPO, M., RESTREPO, B. & DOMÍNGUEZ, J., 2004.- Estudio de Tripanosomiasis americana en dos poblados indígenas de la Sierra Nevada de Santa Marta. Colombia. Revista CES Medicina, 18(1): 43-50. [ Links ]
PATTERSON, J.S., BARBOSA, S.E. & FELICIANGELI, M.D., 2009.- On the genus Panstrongylus Berg 1879: Evolution, ecology and epidemiological significance. Acta Tropica, 110:187-99. [ Links ]
PARRA-HENAO, G., ANGULO, V., JARAMILLO, N. & RESTREPO. M., 2009.- Triatominos (Hemiptera: Reduviidae) de la Sierra Nevada de Santa Marta, Colombia. Aspectos epidemiológicos, entomológicos y de distribución. Revista CES medicina, 23(1): 17-26. [ Links ]
PROUS, X., 2005.- Entradas de cavernas, interfaces de biodiversidade entre ambientes externos e subterrâneos: Distribuição dos artrópodes da Lapa do Mosquito, Minas Gerais. Dissertação de mestrado apresentada no programa de Pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre. Universidade Federal de Minas Gerais. Tese de Mestrado. [ Links ]
READIO, P.A., 1927.- Studies on the biology of the Reduviidae of America north of Mexico. Kansas Univ Sci Bull, 17: 1-291. [ Links ]
SCHUH, R. & SLATER. J., 1995.- True bugs of world (Hemiptera, Heteroptera). Classification and natural history. Cornell University, Ithaca. 336 pp. [ Links ]
SILVA, A.C. & GIL-SANTANA, H.R., 2004.- Predation of Apiomerus pilipes (Fabricius) (Hemiptera, Reduviidae, Harpactorinae, Apiomerini) over Meliponinae bees (Hymenoptera, Apidae), in the State of Amazonas, Brazil. Revista Brasileira de Zoologia, 21: 769-774. [ Links ]
SOTO-VIVAS, A., LIRIA, J. & DE LUNA, E., 2009.- Morfometría geométrica y filogenia en Rhodniini (Hemiptera, Reduviidae) de Venezuela. Acta Zool. Mex, 27(1): 87-102. [ Links ]
TONN, R.J.; CARCAVALLO, R. & ORTEGA, R., 1976.- Notas sobre la biología, ecología y distribución geográfica de Rhodnius robustus Larrouse, 1927 (Hemiptera, Reduviidae). Bol. Dir. Malariol. Saneam. Amb, 16:158-162. [ Links ]
VÁSQUEZ, L.R., JARAMILLO, E. & MOLINA, E.J.A., 2007.- Triatominos del municipio de Mocoa, departamento del Putumayo. Revista Facultad de Ciencias de la Salud Universidad del Cauca, 9(3): 46-50. [ Links ]
VEGA, S., MENDOZA, A. & CABRERA, R., 2006.- Primer caso de enfermedad de Chagas aguda en la Selva Central del Perú: investigación de colaterales, vectores y reservorios. Rev. perú. med. exp. salud pública, 23(4) 288-292. [ Links ]
WEAVER, E.C., CLARKE, E.T. & WEAVER, N., 1975.- Attractiveness of an assassin bug to stingless bees. J. Kans. Entomol. Soc, 48: 17-18. [ Links ]
ZHANG, G. & WEIRAUCH, C., 2011.- Matching dimorphic sexes and immature stages with adults: resolving the systematics of the Bekilya group of Malagasy assassin bugs (Hemiptera: Reduviidae: Peiratinae). Systematic Entomology, 36: 115-138. [ Links ]

