



![THREE NEW SUBSPECIES OF Elzunia bomplandii (Guérin-Ménéville [1844]) FROM COLOMBIA (NYMPHALIDAE: DANAIDAE: ITHOMININAE)](/img/en/prev.gif)







 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
In a global context, regarding Aphodiinae beetles, SKELLEY (2008) points out 12 tribes worldwide of which only make presence in the New World: Aegialiini, Aphodiini, Didactyliini, Eupariini, Odontolochini, Proctophanini, Psammodiini, Rhyparini and Stereomerini; represented by approximately 128 genera and 816 species, grouping which, as stated before, is very dynamic and can change, due to frequent revisions of the theme. The taxonomy of the group, on a large scale, still has large gaps (WOODRUFF, 1973). However, recently, a more natural arrangement has been developed for the group, although there are still groups of uncertain location (DELLACASA et al., 2001; SKELLEY, 2008).
In Colombia, Aphodiinae beetles1 form a modestly diversified group, associated with vertebrate manure, organic matter in decomposition and some have associations with other organisms such as ants, termites or are tenants of vertebrate caves (CÁRDENAS & PÁEZ, 2016; GIRALDO et al., 2001; WOODRUFF, 1973).
Although frequent in varíous wild and intervened habitats, this group of beetles go unnoticed because of their cryptic habits (many are fossorial and live on or under the substrate) or their appearance and size. They are small beetles, in their most of 5-8 mm in length and dark coloration, which is confused with the natural environment (BATES, 1887; DELLACASA et al., 2001; GALANTE et al., 2003; STEBNICKA, 2001).
Despite its ecological value is very significant by participating in the degradation of vertebrate manure in livestock systems or develop such work in the wild, in which case, they are part of the body saprophyte. On the other hand, are part of the energetic bridge between the available organic matter and other organisms of the natural environment that prey on them or benefit from their edaphogenic work (WOODRUFF, 1973).
In Colombia, there are few records of this group, being the “checklist of Blackwelder” (BLACKWELDER, 1944) the first work that compiles the species for the Neotropical region, including Colombia, which registered at the time 6 genera and 24 species (BLACKWELDER, 1944 p 212-216). List that, although effective in many of the registries, has shown such a number of systematic changes, that it is not very useful for modern studies (CABRERO et al., 2010; SKELLEY et al., 2007ª; 2008).
In Colombia, a few other records can be found in the works of pioneer entomologists, for example FIGUEROA (1977) mentions Aphodius brasiliensis Laporte (currently Trichaphodiellus brasiliensis) and Aphodius lividus Olivier (currently Labarrus lividus) in human and cattle excrement; the first in Valle del Cauca and the second in Valle del Cauca, Tolima and Cundinamarca. In recent years, GIRALDO et al. (2001) mentioned at least two species of Aphodiinae of great interest in the degradation of dung in silvopastoral systems in the Colombian Andean region, making specific mention of Aphodius brasiliensis (whose new identification we already mentioned). Tests on breeding in the laboratory were carried out for Aphodius granarius (currently Calamosternus granarius [Linnaeus, 1767]) registered by CARDENAS-CASTRO & PAEZ-MARTINEZ (2017).
By supporting the identification of a material sent by a group of researchers, we found great difficulty in grouping and identifying them. A problem that DELLACASA et al. (2001) explain very well when expressing the difficulties of the Aphodini, due to the multiplicity of registered genres, which makes identification difficult, even at that level, which is why we had to carry out an exhaustive bibliographic compilation, in which the complexity of the group and others was evidenced. Aspects are: the little information available in the national literature on the subject, the taxonomic complexity and agroecological value of the group. As well as the desire to promote the systematic and agroecological study of this valuable group of beetles, motivated this investigation, which set out to develop a catalog of species of Aphodiinae for Colombia, to provide taxonomic keys at the level of genera and species useful for the development of local studies.
METHODOLOGY
Initially, we did an exhaustive review of the literature, in which the general context of the group at the level of the Western Hemisphere was raised, for which the pages of the guide for genera of the New World beetles were consulted, especially the documents made by SKELLEY (2007, 2008). From there, multiple bibliographic records were examined to raise the context of the Aphodiinae beetles, in such a way that in each annotated tribe the records present for Colombia were reviewed. It is very difficult to harmonize the taxonomic arrangement, given the persistent disparity of proposals of the different authors. However, the main taxonomic arrangement was made following the SKELLEY (2008) proposal. However, a few changes have been enhanced as reviewed by other authors, in which case the source will be mentioned.
Additionally, some species were reviewed directly from specimens present in the collection of the first author (Collection Familia Pardo-Locarno (CFPL-COL). In past occasions, some copies of the collection were reviewed by Fernando VAZ DE MELLO (durante 2007), others, which served as a comparison were donations made by colleagues Leonardo Delgado Castillo (Inecol, Mexico) in 1993 and Carlos A.H. Flechtmann in 1995.
RESULTS AND DISCUSSION
Initially we will present a general context of the group, followed by the catalog for groups and species registered in Colombia and finally, taxonomic keys (tribes, genera and species).
Basic Aspects of subfamily Aphodiinae Leach, 1815
According to SKELLEY (2007), it has sizes less than 15 mm, many being smaller than 8 mm. In addition, the body is elongate to oval, the clypeus is usually expanded and normally covers the buccal apparatus, and the jaws are reduced. Antennas have nine segments, apically with an expansion of three segments, antennal club with 3-7 segments, first segment simple, not hollowed out. Mesocoxae almost always contiguous, the metatibia usually has two apical spines, the abdomen has six visible sternites, the pygidium is partially or totally covered by the elytra and the tarsus almost always with claws.
The small dung beetles as they are known, alludes to the most frequently observed food habit. However, many taxa are associated with very different substrates, such as organic matter, leaf litter, ant nests, termites (case Aschnarhyparus peregrinus (Hinton, 1934), registered by the author in nest of Coptotermes sp.) and even litter. They make up a group of diversified Scarabaeidae (12 tribes, 280 genera and 3200 species) and they are widely distributed worldwide (HINTON, 1934; SKELLEY, 2008; STEBNICKA, 2001; WOODRUFF, 1973).
Catalogue of Aphodiinae of Colombia2
We expose the species listed for Colombia in the following order: tribe, genus, species, author and year; in each case the annotated locality for the species will be recorded, as far as possible, according to the authors consulted. Some localities were taken from the initial records, reviews or local studies, very few from first author’s collection. According to the attached catalog, Aphodiinae beetles from Colombia preliminarily recorded 20 genera and 56 species, supported by approximately 47 bibliographic records and authors listed in the bibliography (Figures 1, 2).

Figure 1 A. Side view of Stebnickiella zosterixys tomado de Skelley 2007 figure 28; B. Aschnarhyparus peregrinus (From Hinton, 1934); C. Parapsammodius integer (Bates, 1887) after Skelley; D. Pronotum of Leiopsammodius sp. (Figures B, C y D From Skelley, 2007 figure 17).
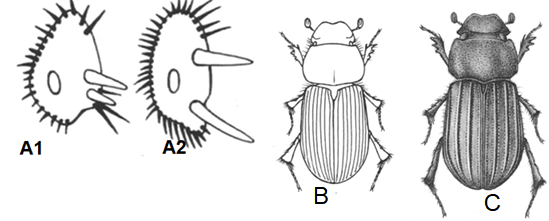
Figure 2 Apical spur of hind tibia A1 Aphodiini A2 Didactyliini; B. Aidophus notatus Harold 1859 (From Dellacasa et al., 2001 Figure 295); C. Xenoheptaulacus tricostatus Harold 1869 (From Dellacasa et al., 2001 figure 927)
Tribu Aphodiini (Sensu DELLACASA et al., 2001)
Xenoheptaulacus tricostatus (Harold, 1869). Magdalena, Santander.
Labarrus pseudolividus3 (Balthasar, 1941). Fonseca, Guajira.
Trichaphodiellus brasiliensis (Laporte, 1840). Popayan (Cauca).
Gordonius rhinocerillusSkelley, Dellacasa & Dellacasa, 2009. Silvia (Cauca).
Neotrichaphodioides caracanus (Balthasar, 1970). San Martin (Meta).
Gonaphodiellus pacatus (Harold, 1880). Fusagasugá (Cundinamarca), Rovira (Tolima).
Gonaphodiellus sexguttatus (Schmidt, 1916). Pichinde (Valle).
Gonaphodioides columbicus (Harold, 1880). Sabana de Bogotá (Cundinamarca), Popayán (Cauca).
Gonaphodioides acutecernans (Balthasar, 1960). Bogotá (Cundinamarca).
Nialaphodius nigrita (Fabricius, 1801). Popayán (Cauca).
Tribu Didactyliini4
Aidophus notatus (Harold, 1859). “Vaterland Columbien”.
Tribu Eupariini
Aphotaenius convexusHarold, 1880. Bogotá, Guasca, (Cundinamarca), La Lucera, Medellín (Antioquia).
Ataenius attenuatorHarold, 1874. Lucera, Medellín (Antioquia).
Ataenius aequalisHarold, 1880. Ambalema, Ibagué (Tolima), Caucathal (Valle), Bogotá (Cundinamarca).
Ataenius carinatorHarold, 1874. “Columbien”.
Ataenius columbicusHarold, 1880. Sabana de Bogotá (Cundinamarca).
Ataenius communisHinton, 1936. Santiago, Norte de Santander (Colombia).
Ataenius complicatusHarold, 1869. Santiago, Norte de Santander (Colombia).
Ataenius gracilisMelsheimer, 1845. “Columbien”.
Ataenius imbricatusMelsheimer, 1845. Cartagena (Bolivar), Santa Marta (Magdalena).
Ataenius moratorHarold, 1869. Leticia (Amazonas).
Ataenius nugatorHarold, 1880. Medellín, (Antioquia), río Meta, Villavicencio (Meta).
Ataenius opatrinusHarold, 1867. Leticia (Amazonas), Sopo 2900 m (Cundinamarca), río Jamundí, Jamundí (Valle).
Ataenius perforatusHarold, 1867. Bogotá, Sopo, 2300 m (Cundinamarca), río Magdalena, Honda (Tolima).
Ataenius platensis (Blanchard, 1846). Bucaramanga (Santander).
Ataenius punctipennisHarold, 1868. Ibagué (Tolima).
Ataenius sculptorHarold, 1868. “Colombia”.
Ataenius scutellarisHarold, 1867. La Estrella 1700 m, Monte Redondo, (Cundinamarca), Leticia (Amazonas.
Ataenius steinheiliHarold, 1874. Barranquilla, (Atlantico), Mompox (Bolivar), “N. Granada”
Ataenius strigicaudaBates, 1887. Cartago (Valle), Ibagué (Tolima), Tame (Arauca), Leticia (Amazonas).
Ataenius tuberculatusSchmidt, 1911. “Colombia”.
Ataenius chinacotaeStebnicka, 2007. Chinacota (Norte de Santander).
Ataenius opacipennis Schmidt, 1910. Colombia.
Ataenius peregrinator Harold, 1887. Colombia.
Ataenius palmaritoensisStebnicka, 2007. Cúcuta (Norte de Santander).
Ataenius cucutaeStebnicka, 2007. Chinacota (Norte de Santander), Honda (Tolima).
Passaliolla aspericeps (Harold, 1876). “La Luzera” (Antioquia).
Passaliolla cancellata (Bates, 1887). Amazonas (Colombia).
Odontolytes huebneri (Petrovitz, 1970). Leticia (Amazonas).
Odontolytes landai (Balthasar, 1963). Ibagué (Tolima).
Odontolytes transversaria (Schmidt, 1909). Honda, Bogotá (Cundinamarca), río Magdalena (Magdalena).
Odontolytes denominatus (Chevrolat, 1864). Colombia.
Odontolytes capitosus (Harold, 1867). Pichinde, Dagua (Valle) “Paramba”
Saprosites breviusculusHarold, 1867. Chinacota (Norte de Santander).
Saprosites dentipesHarold, 1867. Bogotá (Cundinamarca), “Paso del Quinto” (Quindío?).
Saprosites meditansHarold, 1867. Merenberg (Cauca), “La Luzera”, Medellín (Antioquia), Chinacota (Norte de Santander).
Saprosites parallelusHarold, 1867. Chinacota, 2700 m (Norte de Santander.
Saprosites peregrinusRedtenbacher, 1858. “Colombia”.
Saprosites subterraneusPetrovitz 1976. Calarca (Quindío).
Tanyana guyanaensis (Stebnicka, 2003). Leticia (Amazonas).
Batesiana tuberculata (Bates, 1887). Buenaventura (Valle).
Tribu Odontolochini (incertae sedis5)
Stebnickiella zosterixys6 Skelley, 2007. Caparu (Vaupes).
Tribu Psammodiini
Parapsammodius integer (Bates, 1887). Río Frío, Cerro Campana, Santa Marta (Magdalena).
Platytomus gregalis (Cartwright, 1948). Cali, Saladito (Valle).
Platytomus parvulus (Chevrolat, 1864). Río Frío (Magdalena).
Tribu Rhyparini
Aschnarhyparus peregrinus (Hinton, 1934). Santa Ana (Bolivar).
From the previous list, several aspects of the evolution of the Aphodiinae issue in Colombia can be summarized (Figure 3). Examining in a span of 1800 to date, in períods of 20 years, for now the oldest species described would be Nialaphodius nigrita Fabricius, 1801 and the most recent would be Gordonius rhinocerillusSkelley, Dellacasa and Dellacasa, 2009. The species register goes through an initial períod of few discoveries from 1800 to 1860 (from one to five species). The períod between 1860 to 1880 represents the golden age of the discoveries of Colombian Aphodiinae, with 21 species many of them discovered by the famous taxonomist von Harold (Who to date completes at least 25 species). From there, records between 1880 and 1960 fluctuate from 8 to 2 species every two decades. Finally, from 1980 to date, discoveries of new species intensified, highlighting the achievements of entomologist Stebnicka (at least six species) partly supported by the review of specimens often collected by Henry Howden and his wife who made multiple collections in the Andean region, Orinoquia and Amazonia and deposited in their collection which was in turn in a Canadian museum.
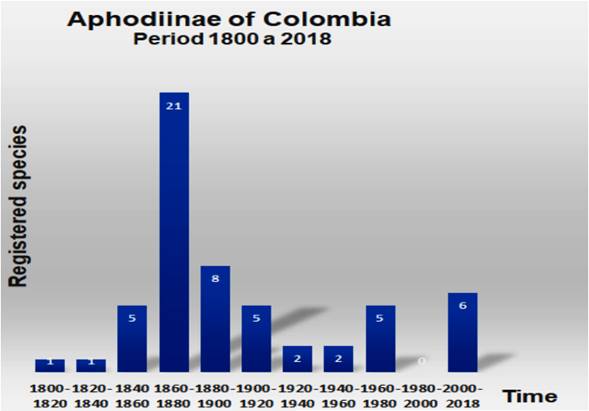
On the other hand, the 56 species registered for Colombia represent a small number for the diversity known worldwide, which according to the authors ranges from 3200 to 3400 species (DELLACASA et al., 2001; SKELLEY, 2008); records for the Aphodiinae of the United States indicate a relatively high diversity: 210 species (RATCLIFFE, 1998), although this records had significant changes with the revision of the Aphodiini tribe for the United States and Canada (GORDON & SKELLEY, 2007). However, this and other studies suggest a great affiliation with cold and seasonal ecosystems of the Arctic regions, although there are multiple endemisms everywhere on all continents and in all habitats (BLACKWELDER, 1944; CABRERO et al., 2010; GORDON & SKELLEY, 2007). In this sense, the figure of Colombian Aphodiinae is lower than what is known for Mexico (111 species according to CABRERO et al., 2010), but it is higher than other nearby continental records (for example the Southern Cone of Chilean Patagonia and Argentina and its margin subantarctic totaled 27 species according to SMITH & SKELLEY (2007). These two studies (CABRERO et al., 2010 and SMITH & SKELLEY 2007), are the most recent and conscientious regional surveys on Aphodiinae for countries or large regions of Central and South America. Another of the few regional studies of Neotropical Aphodiinae raised 35 Aphodiinae species belonging to two tribes and 12 generates Río Grande do Sul and Santa Catarina, Brazil (SILVA, 2015).
Colombian Aphodiinae Key (Modified from DELLACASA et al., 2001; SKELLEY, 2007; STEBNICKA, 2000, 2001A; 2001B, 2002, 2003A; 2003B, 2004, 2005, 2006A; 2006B, 2007).
1. Anteríor clypeal edge flattened, broadly margined (double edged), many with a distinct inwardly projecting spine or tooth at middle; pronotum somewhat swollen anteríorly (tumid); head able to be deflexed at nearly 90o to pronotal surface; when head deflexed, clypeus and anteríor pronotal lobes form cavity for fore leg; posteríor lateral margins of pronotum (usually) emarginate with denticles; mesocoxae widely separated; meso-metasternal suture straight and mesosternum flat; protibial teeth of many close and anteríorly placed. … Odontolochini (at the moment a single species: Broad head, flattened clypeal margin, and anteríorly tumid pronotum with explanate sides and basal constriction. Stebnickiella zosterixys Skelley, 2007 (Figure 1A).
1’. Other combination of characters
2. Pronotum with at least five longitudinal, parallel ridges. Myrmecophilous, termitophilous and/or saprophagous species. Oriental, Ethiopian, Australian and Neotropical regions. Rhyparini Schmidt, 1910 (see AschnarhyparusMakhan, 2006), only at the moment Aschnarhyparus peregrinus (Hinton, 1934). (Figure 1B).
2’. Pronotum without longitudinal ridges.
3. Pronotum either with five transverse ridges, five transverse furrows and posteríor longitudinal furrow (Figure 1D), sometimes also with a pair of accessory swellings between fourth and fifth ridges, or with reduced pronotal structure, derived from these formations; depressed areas in anteríor corners, lateral elevated areas corresponding to fusion of second to fifth ridges, lateral impressions corresponding to ends of third transverse furrow and/or vestiges of transverse furrows and posteríor longitudinal furrow. Exclusively psammophile species. All zoogeographic region. Psammodiini Mulsant, 1842. (Figure 1D).
3’. Pronotum neither with transverse ridges, nor with vestigial formations.
4. Basal margin of elytra never bordered; pygidium not furrowed laterally; femora never grooved at fore or hind margin; transverse carinae of middle and hind tibiae rarely missing; abdominal sternites usually feebly sclerificate and not fused one another but never with a transverse belt of short longitudinal carinae; apical spurs of hind tibiae separated each from other so that the first tarsal segment is articulated in between them; epipharynx with acropariae of different length and size.
4’. Basal margin of elytra bordered, border more or less distinct, sometimes seventh and eighth interstices are basally bordered only; pygidium usually furrowed laterally; femora generally grooved at fore or hind margin; transverse carinae of middle and hind tibiae usually lacking, only rarely existing; abdominal sternites strongly sclerificate, fused one another and with a transverse belt of short longitudinal carinae; apical spurs of hind tibiae attached each other so that first tarsal segment does not raise between them; epipharynx without acropariae. Eupariini Schmidt, 1910. 16.
5. Mostly plumper animals, elytra mostly considerable broader behind. Profemur narrower than metafemur. Basimetatarsomere considerably, asymmetrically widened apically. Parapsammodius Verdu, Stebnicka & Galante, 2006, Parapsammodius integer (Bates, 1887) (Figure 1C).
5’. Mostly slender animals, elytra parallel, subparallel, at most moderately broader behind. Profemur as wide as or wider than metafemur. Basimetatarsomere elongate, subcylindrical, only slightly to moderately, symmetrically widened apically. Platytomus Mulsant, 1842. (Figure 4A, 7C).

Figure 4 Dorsal view of several species, taken and modified from its authors, the citation of each one is mentioned in parentheses. A. Platytomus varíolosus (from Wikipedia); B. Gordonius rhinocerillus (Skelley 2009 Fig. 4); C. Labarrus pseudolividus (Smith & Skelley, 2007 Fig 57); D. Trichaphodiellus brasiliensis (Dellacasa et al., 2001 Pl 894).
6. Median longitudinal metathoracic line strong, complete; slightly smaller species (see CARTWRIGHT, 1948). Platytomus parvulus (Chevrolat, 1864).
6’. Median longitudinal metathoracic line obsolete anteríorly, posteríor half strong and deep, posteríor femoral line strong (see CARTWRIGHT, 1948). Platytomus gregalis (CARTWRIGHT, 1948).
7. Apical spurs of hind tibiae separated each from other, so that first tarsal segment is articulated in between them (Figure 2 A1, A2); abdominal sternites not fuse done another. Aphodiini.
7’. Apical spurs of hind tibiae very closely attached each other so that first tarsal segment does not through them (Figure 2B); abdominal sternites fuse done another. Didactyliini Clypeus distinctly sinuate at middle; genae ciliate; elytra glabrous. Aidophus notatus (Harold, 1859) (Figure 2B).
8. Third, fifth and seventh elytral interstices costiform, other flat (Figure 2C); epipleura wide, distinct till suture apical angle; eyes very large. Reddish-testaceous. Length 3.5-4 mm. Panama, Venezuela, Colombia. XenoheptaulacusHinton, 1934 (Xenoheptaulacus tricostatusHarold, 1869) (Figure 2C).
8’. Not all of the above characters simultaneously present.
9. Basal margin of pronotum bordered. Protibia finely punctate dorsally, ventral inner margin of protibia with 3 widely spaced tubercles; metatibiae apically fimbriate with equal, very short spinules; male with prominent clypeal horn. Length 5.5-6 mm. GordoniusSkelley, Dellacasa & Dellacasa, 2009 (Gordonius rhinocerillus Skelley, Dellacasa & Dellacasa, 2009) (Figure 4B).
9’. Basal margin of pronotum not bordered.
10. Scutellum pentagonal, lateral margins parallel or convergent toward base. Elytra dirty yellowish, usually with a discal brownish spot more or less distinct; head and pronotal disc more or less darkened. Length 3-7 mm. Labarrus Mulsant & Rey, 1869. (Labarrus pseudolividusBalthasar, 1941) (Figure 4C).
10’. Scutellum triangular
11. Pronotum with hind angles obtusely subtruncate.
11’. Pronotum with hind angles obliquely truncate.
12. Head moderately convex with epistome gibbous, nearly smooth, with a more or less distinct central tubercle anteríorly from the arcuate and not tuberculate frontal suture; clypeus feebly sinuate at middle, rounded at sides; genae obtusely rounded, prominent. Pronotum doubly punctured and in males, with small median superficial impression at front margin; shoulder not denticulate. Reddish-testaceous; head and pronotum disc darker; elytra yellowish darker on disc and with preapical indefinite darker spots. Length 6-8 mm. Trichaphodiellus A. Schmidt, 1913 (Trichaphodiellus brasiliensis Castelnau, 1840) (Figure 4D).
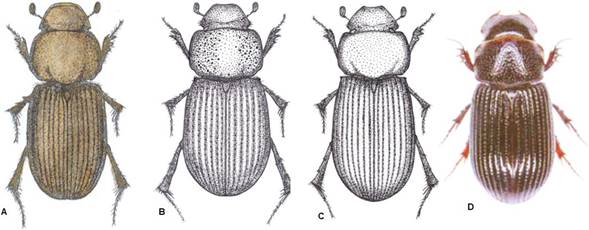
Figure 5 Dorsal view of several species, taken and modified from its authors, the citation of each one is mentioned in parentheses. A. Neotrichaphodioides caracanus (From Dellacasa et al 2010 Fig. 9); B. Gonaphodiellus pacatus (Dellacasa et al., 2012 Fig. 53); C. Gonaphodioides acutecernans (Dellacasa et al., 2012 Fig. 70); D. Aphotaenius convexus Harold (Stebnicka, 2009, Fig 56).
12’. Head with epistome almost flat, sparsely punctulate; clypeus almost semicircular or subtruncate anteríorly, widely rounded at sides, glabrous; genae acutely angulate or obtusely rounded, not to very shortly bristled, more or less strongly protruding; frontal suture finely impressed, mutic. Pronotum transverse, feebly convex, evenly or dually punctured. Piceous, brownish-red or brownish-yellow, sometimes elytra cloudy darkened on disc. Length: 3.5-8.0 mm. NeotrichaphodioidesDellacasa, Dellacasa & Skelley, 2010 (Neotrichaphodioides caracanusBalthasar, 1970) (Figure 5A).
13. Elytra more or less strongly denticulate at shoulder, rarely lacking denticle. First joint of labial palps as long as second; clypeus subtruncate anteríorly or very feebly sinuate at middle, round at sides; elytra oval, more or less broadened posteríorly, finely striate; body feebly or, at most, moderately convex; mostly medium size species (length 5.0-6.0 mm). Taxa more or less dark brownish, with or without paler elytral spots. Gonaphodiellus Schmidt, 1913
13’. Elytra denticulate or not at shoulder. First joint of labial palps longer than second; body more convex; mostly rather small species (length 4-5 mm). Clypeus deeply sinuate at middle, angulate at sides; elytra subparallel-sided or oval elongate, finely striate; body more or less distinctly convex. Taxa chestnut brown or reddish testaceous, rarely blackish. Gonaphodioides Dellacasa, Dellacasa & Gordon, 2012.
14. Genae rounded, not or faintly protruding from the eyes. Elytral interstriae feebly convex, glabrous, preapically alutaceous thus almost dull; truncation of hind angles of pronotum inwardly sinuate. Dark brownish; clypeal margin and fore angles of pronotum reddish; elytra yellowish spotted basally on second and third interstriae and at shoulder, posteríorly with more or less elongate and widened blackish margined yellowish spots on second, fourth, sixth and seventh interstriae. Length 4.5-5 mm. Colombia, Costa Rica, El Salvador, Honduras, Mexico (Chiapas), Panama. Gonaphodiellus sexguttatus (Schmidt).
14’. Genae obtuse, protruding from the eyes. Pronotum on sides dually, somewhat irregularly punctured; clypeal lateral margins arcuate. Brown-yellowish, epistome, pronotal and elytral disc usually darker. Length 5.5-6.0 mm. Colombia, Costa Rica, Panama. Gonaphodiellus pacatus (Harold) (Figure 5B).
15. Elytra subparallel-sided, very elongate. Chestnut-brown. Length 5.5-6.0 mm. Bolivia, Colombia. Gonaphodioides acutecernans (Balthasar) (Figure 5C).
15’. Elytra oval, less elongate. Elytral interstriae distinctly convex. Chestnut-brown. Length 4.0-5.0 mm. Colombia, Venezuela. Gonaphodioides columbicus (Harold) 16. Head very broad. Elytra with irregular tubercles and swellings. BatesianaChalumeau 1983Batesiana tuberculata (Bates) (Figure 7C).
16’. Different. Head less broad, as much as the pronotum or less. Elytra without tubercles
17. Clypeus with broad, inflexed edge extending from gena; side margins of pronotum with two or more denticles and/or angulations; middle coxae widely separated. Odontolytes Koshantschikov, 1916.
17’. Clypeus without broadly inflexed edge, rarely with triangular, inflexed process at middle; side margins of pronotum without denticles, frequently posteríor angles of pronotum denticulate; middle coxae contiguous, subcontiguous or separated.
18. Body moderately convex, surface varíously sculptured; head small to moderate in size; disc of pygidium deeply eroded. … AtaeniusHarold, 1867.
18’. Body strongly convex to deplanate, surface punctate; head moderate in size to broad; disc of pygidium eroded, scabrously punctate or shining, smooth.
19. Body stout, suboval, black, strongly shining, glabrous; clypeal median edge between angulations or denticles triangularly flattened upward. Aphotaenius Cartwright, 1952. Aphotaenius convexus (Harold, 1880) (Figure 6C).

Figure 6 Dorsal view of several species, taken and modified from its authors, the citation of each one is mentioned in parentheses. A. Tanyana guynaensis (Stebnicka, 2009, Fig. 82); B. Odontolytes denominatus (Stebnicka, 2009, Fig. 35); C. Ataenius columbicus (Stebnicka, 2007, Fig. 85); D. Passaliolla aspericeps (Stebnicka, 2007, Fig. 59); E. Odontolytes landai (Stebnicka, 2007, Fig. 40).
19’. Body elongate, parallel-sided.
20. Body strongly deplanate dorso-ventrally; abdominal sternites laterally with large fovea. Passaliolla Balthasar, 1945.
20’. Body convex, not deplanate; abdominal sternites without fovea.
21. Pronotal lateral margin with tooth in posteríor third; meso- and metatibiae flattened, apical spurs seta-like. Tanyana Stebnicka, 2006. … Tanyana guynaensis (Stebnicka, 2003) (Figure 6A).
21’. Pronotal lateral margin without tooth in posteríor third; meso- and metatibiae not flattened, apical spurs spine-like; head broad, strongly gibbose medially; elytral striae usually with coarse punctures; abdominal sternites strongly coalesced, fluted along sutures; disc of pygidium eroded; external sexual dimorphic characters not advanced. SaprositesRedtenbacher, 1858.
22. Colour black; pronotum subquadrate, posteríor angle widely truncate, side steep, not explanate; mesosternum with cordate callosity; metafemur parallel-sided, posteríor marginal line incomplete. Length 6.0-8.0 mm. Odontolytes capitosus (Harold, 1867)
22’. Colour castaneous or piceous; pronotum transverse, posteríor angle right-angled, prominent, side more or less explanate; mesosternum with carina or with semioval callosity.
23. Scutellum smooth or finely punctate; disc of elytra convex. Pronotal base with fine marginal line. Elytral striae in posteríor half delimited on each side by fine lines; intervals usually distinctly punctate, apically carinate, 10th interval with median row of minute granules. Length 6.0 -7.5 mm. Odontolytes denominatusChevrolat, 1864 (Figure 6B).
23’. Scutellum foveate or carinate; disc of elytra slightly deplanate. Pronotal base lacking marginal line. Mesosternum with callosity.
24. Head on each side of median convexity with moderate punctures separated by less than one diameter; mesosternal callosity wide without carina inside. Pronotal punctures on each side of disc same size as those on clypeal disc; elytra slightly widened in posteríor third. Length 4.8-5.2 mm. Odontolytes transversaria (Schmidt, 1909).
24’. Different. Another group of characters.
25. Clypeus truncate anteríorly or median emargination indistinct, surface without transverse carina; elytral preapical umbone indicated; colour piceous. Anteríor angle of pronotum widely rounded, strongly produced forwards, side widely explanate; mesosternum with semioval callosity. Length 4.8-5 mm. Odontolytes huebneri (Petrovitz, 1970)
25’. Clypeus truncate anteríorly or median emargination indistinct, surface without transverse carina; elytral preapical umbone indicated; colour piceous. Anteríor angle of pronotum obtuse, slightly prominent, side narrowly explanate; mesosternum with carina. Elytra shining, discal intervals with distinct rows of punctures, lateral intervals wider than striae with granules or tubercles. Length 3.0- 4.5 mm. Odontolytes landai (Balthasar, 1963) (Figure 6E).
26. Clypeal surface in anteríor third or in anteríor half in most species or sexes with more or less pronounced transverse wrinkles (except some species of Ataenius aequalisplatensis group).
26’. Clypeal surface varíously punctate and/or granulate, not transversely wrinkled (except some species of Ataenius aequalis-platensis group).
27. Body in most species bicoloured; meso- and metafemora along anteríor margin with fringe of hair-like setae, usually without posteríor lines; metatibiae in most species slightly deplanate dorso-ventrally. Ataenius terminalis group (not represented in Colombia).
27’. Body unicoloured, frequently elytra lighter than head and pronotum; mesoand metafemora without fringe of setae along anteríor margin, posteríor lines of metafemora complete, incomplete or absent; metatibiae subcylindrical.
28. Male genitalia in most species with parameres longer than phallobase, lanceolate, frequently toothed apically.
28’. Male genitalia in most species with parameres usually as long as phallobase, rounded or acute apically, without teeth.
29. Body slender, approximately 3.0-4.8 mm in length; side of pronotum and elytra with belt of fine, close punctures extending from anteríor angles of pronotum to Apex of elytra. Ataenius nugator group.
29’. Body stout, approximately 5-7 mm in length; side of pronotum and elytra varíously punctured or smooth, without belt of close punctures Ataenius crenator group (not represented in Colombia).
30. Colour varíous, yellowish brown to black; clypeal surface in most species with transverse wrinkles, in some species punctate or granulate; metatibia apically in most species without accessory spine. Ataenius aequalis-platensis group.
30’. Colour piceous to black; clypeal surface with transverse wrinkles; metatibiae apically in all species with distinct accessory spine.
31. Male genitalia with parameres slender, converging apically. Ataenius strigatus group (not represented in Colombia).
31’. Male genitalia with parameres wide, not converging apically. Ataenius strigicaudus group.
32. Dorsal surface in most species tightly encrusted by argillaceous coating or only a part of body covered with coating; base of pronotum usually without marginal line; intervals of elytra in some species alternately different. Ataenius imbricatus group.
32’. Body not encrusted by argillaceous coating; base of pronotum margined; intervals of elytra not alternately different.
33. Elytral striae in most species bordered on each side by undulate lines, intervals more or less carinately elevated.
33’. Elytral striae not bordered by undulate lines, intervals in some species carinately elevated and/or granulate or swollen.
34. Body large, elongate; clypeal anteríor margin rounded or obtuse on each side of median emargination; scutellum foveate; metatibiae apically with accessory spine. Ataenius perforatus group.
34’. Body small to moderate in size, in most species oblong oval; clypeal anteríor margin rounded or denticulate on each side of median emargination; scutellum without foveae; metatibiae in most species without accessory spine. Ataenius complicatus group.
35. Clypeal surface and middle of head with fine to moderate punctures, intervals of elytra in most species smooth; metatibiae apically in some species with accessory spine. (Ataenius scutellaris group) only Ataenius scutellarisHarold, 1867.
35’. Clypeal surface with granules or umbilicate punctures, middle of head frequently with longitudinal lines of united punctures; intervals of elytra smooth or granulate or swollen; metatibiae apically in all species without accessory spine. Ataenius texanuscarinator group.
36. Parameres of male aedeagus in lateral view narrowed apically. Ataenius nugatorHarold, 1880.
36’. Parameres of male aedeagus in lateral view widened apically. Ataenius communis
37. Clypeal margin widely rounded; terminal spur of protibia in male straight or variably curved. South America, West Indies. Ataenius punctipennisHarold, 1868.
37’. Clypeal margin more or less obtusely angled; terminal spur of protibia in male slightly bent downward or inward.
38. Body elongate, castaneous; pronotum with fine and moderate punctures, the latter widely spaced on disc; elytra shiny; basitarsomere of metatarsus longer tan upper tibial spur. Southern USA to Argentina. Ataenius platensis (Blanchard, 1846)
38’. Body oblong oval, piceous to black; pronotum with fine and moderate to coarse punctures, the latter close on disc; elytra dull; basitarsomere of metatarsus shorter tan upper tibial spur (may be equal in length). Louisiana, Central and South America, West Indies. Ataenius aequalisHarold, 1880.
39. Head strongly alutaceous, often scabrously punctured on each side of median gibbosity; elytral intervals in apical fourth more or less eroded on each side, discal interval 1-4 strongly crenate by punctures. Central and South America, West Indies. Ataenius strigicaudusBates, 1887.
39’. Head microreticulate, finely to moderately punctured on each side of median gibbosity; elytral intervals not eroded apically, discal intervals 1-4 with moderately crenating punctures. Body mostly dark brown to piceous, shining; head strongly gibbose medially, clypeal emargination deep. South America. Ataenius columbicusHarold, 1880 (Figure 6C).
40. Body without coating, opaque; base of pronotum with conspicuous marginal line. Punctures of pronotal disc fine to moderate; meso- and metatibiae without setigerous denticles at inner side. Length 3.8-4.2 mm. Brazil: Bahia, Argentina, Bolivia, Colombia, Ecuador, Guiana, Paraguay, Peru, Suriname, Venezuela, Guadeloupe, Trinidad. Ataenius moratorHarold, 1869.
40’. Body entirely or partially covered with argillaceous coating or oily dirt; base of pronotum without marginal line.
41. Elytral intervals alternately higher. Colombia, Bolivia, Brazil, Guiana, Venezuela. Ataenius tuberculatusSchmidt, 1911.
41’. Elytral intervals not alternately higher. Panama, Argentina, Belize, Brazil, Colombia, Costa Rica, El Salvador, Guatemala, Guiana, Honduras, Mexico, Nicaragua, Paraguay, Suriname, Venezuela, Cuba, Bahamas, Dominican Republic, Puerto Rico, St. Croix, Guadeloupe, Trinidad, Barbados, Canada, Eastern United States to South Dakota and Texas, South Carolina. Ataenius imbricatus Melsheimer, 1844.
42. Pronotal posteríor angle rounded or slightly emarginate, surface punctures medium sized. Ataenius opatrinusHarold, 1867.
42’. Pronotal posteríor angle distinctly excised, surface punctures usually large. Head fine, shallow punctate, irregularly coarsely punctate or punctuation absent. Clypeal side in front of gena slightly emarginate, clypeal surface usually with coarse punctures. Ataenius perforatusHarold, 1867.
43. Dorsum entirely or in part covered with conspicuous, erect setae. Length 5.0-7.0 mm. Central and South America, Lesser Antilles. Ataenius complicatusHarold, 1869.
43’. Dorsum in part covered with minute pubescence or glabrous.
44. Dorsum shiny; elytral intervals only slightly convex at middle, narrowly flattened and punctate on each side. Length 5.6-7.0 mm, USA, Mexico, Colombia, Guatemala, Honduras, El Salvador. Ataenius sculptorHarold, 1868.
44’. Dorsum opaque; elytral intervals narrowly carinate at middle, deplanate and impunctate on each side. Length 3.5-4.0 mm, Colombia, Mexico, Venezuela. Ataenius steinheiliHarold, 1874.
45. Species with clypeal margin denticulate.
45’. Species with clypeal margin rounded.
46. Large species. 4.5-4.7 mm. Cuticle shining, glabrous dark broawm to carbon black. Head broad, moderately gibbose at middle. Ataenius chinacotaeStebnicka, 2007.
46’. Smaller species. 2.8-4.2 mm. Dorsum different.
47. Medium size. 3.4-4.2 mm. Body weakly shining to opaque, dark reddish Brown to piceus. Ataenius attenuatorHarold, 1874.
47’. Small size. 2.8-3.5 mm. Dorsum different.
48. Rank 2.8-3.5 mm. Body moderately to strongly shining. Reddish brown to piceous, glabrous. Ataenius peregrinator Harold, 1887.
48’. Rank 2.8-3.2 mm.
49. Body shagreened weakly shining. Surface of head and pronotum minutely pubescent, elytra distinctly setigerous. Pronotal punctures everywhere fine and close on disc separated by less than one diameter from halfway to sides contiguous and diagonelly confluent toward anteríor angles. Ataenius palmaritoensisStebnicka, 2007.
49’. Body not shagreened weakly shining. Pronotal punctures different. Cuticles castaneous shining. Pronotum subquadrate, relatively long, side margin minutely setigerous, posteríor angles rounded. Ataenius cucutaeStebnicka, 2007.
50. Big species. 2.8-3.5 mm. Head moderately convex, median emargination shallow. Cuticle piceous, moderately to strongly shining, legs redish brown. Punctures of head united in short longitudinal lines; elytral intervals strongly convex. Middle femur with strong complete marginal line; elytral intervals almost carinate. Ataenius gracilis (Melsheimer, 1844).
50’. Small species. 2.2-2.5 mm. Head with small gibbosity. Body dull, reddish Brown to piceous, coverd with minute pale setae. Ataenius opacipennis Schmidt, 1910.
51. Length 3.8-4.0 mm. Head with granules extending from clypeal margin to vertex, frontal suture marked by fine carina, groove above suture distinct. Passaliolla aspericeps (Harold, 1876) (Figure 6D).
51’. Length 2.8-3.2 mm. Head granulate in varíous degrees or without granules, frontal carina lacking, groove weakly marked or invisible. Disc of pronotum with fine and larger punctures, the latter separated by 1-2 their diameter. Clypeal surfasse below median gibbosity with band of transverse granules, clypeal carina lacking; elytra about 2 times as long as pronotum, elytral striae groove deep. Passaliolla cancellata (Bates,1887).
52. Large species. Length 5.0-6.0 mm.
52’. Medium sized and small species. Length 2.4-4.8 mm.
53. Clypeus just above median emargination smooth, without carina, frontal suture not indicated, gena small rounded, weakly produced; apical accessory spines of metatibia small. Length 5.0-5.1 mm, greatest width 1.8-1.9 mm. Saprosites meditansHarold 1867 (Figure 7A).

Figure 7 Dorsal view of several species, taken and modified from its authors, the citation of each one is mentioned in parentheses. A. Saprosites meditans (Stebnicka, 2009, Fig. 72); B. Tanyana guyanaensis (Stebnicka, 2009 Fig 82); C. Batesiana tuberculata (Stebnicka, 2009 Fig 20); D. Platytomus micros (From Generic guide to New World Scarab Photo by P Skelley 2008).
53’. Clypeus just above median emargination with transverse carina, frontal suture elevated, gena right-angled, strongly prominent; apical accessory spines of metatibia strong. Head in male almost impunctate, in female coarsely granulate; disc of elytra not deplanate, strial punctures not very coarse, lateral striae narrower than intervals. Length 5.0-6.0 mm, greatest width 2.0-2.2 mm. Saprosites dentipesHarold, 1867.
54. Medium sized species. Length 3.5-4.8 mm.
54’. Small species. Length 2.4-2.8 mm.
55. Lateral marginal line of pronotum thick, grooved, with row of large punctures inside groove. Pronotum not everywhere, not uniformly punctate, punctures mixed fine and larger, varíously spaced; elytral striae with moderate punctures, lateral striae narrower than intervals. Larger punctures on pronotal disc separated by 1-3 diameters, closer at anteríor and posteríor angles. Gena right-angled, exceeding eye; posteríor angles of pronotum truncate, base slightly sinuate. Length 4.2-4.8 mm, greatest width 1.5-1.6 mm. Saprosites parallelusHarold, 1867.
55’. Lateral marginal line of pronotum thin, without groove and punctures. Pronotal sides everywhere finely uniformly punctate without group of larger punctures at anteríor and posteríor angles. Length 3.0-4.0 mm, greatest width 1.1-1.3 mm. Saprosites breviusculusHarold, 1867.
56. Body elongate; pronotal sides parallel; elytra parallel, disc slightly deplanate. Length 2.4-2.6 mm, greatest width 0.8-0.9 mm. Saprosites peregrinus Redtenbacher, 1857.
56’. Body suboval; pronotal sides converge posteríorly; elytra slightly arcuate, disc convex. Pronotum moderately convex, not tumid anteríorly, posteríor angles widely truncate to base, base sinuate; mesosternum punctate without callosities. Length 2.5-2.8 mm, greatest width 0.8-0.9 mm. Saprosites subterraneusPetrovitz, 1976.
CONCLUSIONS
The study of the Aphodiinae beetles of Colombia, a fertile field for many developments, is just beginning and is expected to change rapidly, both in the species register and in its distribution, as new studies provide data. It is expected that this contribution will support or encourage several authors.
The first group of species that can present more changes or additions would be that of cosmopolitans and euriphagas; in this sense, records such as Haroldiellus sallei Harold, 1863, Calamosternus (Linnaeus, 1767) etc., could emerge very soon for Colombian geography. However, it is expected that new and unsuspected registries will join the list, since the registries of neighboring and bordering countries encourage such a hypothesis.

