











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
A fundamental quest of ecological research for more than half a century has been unveiling the processes structuring biological diversity patterns at different scales (HUBBELL, 2001; HUTCHINSON, 1991; LEIBOLD, 2008; WHITFIELD, 2002; WHITTAKER, 1972). This quest is particularly relevant for understanding and coping with the effects of human-induced ecosystem fragmentation. Such is the case of tropical forest fragmentation, which is a major threat to global biological diversity (HADDAD et al., 2015; TURNER, 1996). Before fragmentation a continuous forest cover over a wide geographic extent, containing a diverse biota, is the dominant feature. After fragmentation, the continuous forest cover is converted to a mosaic of discrete patches within a matrix of exotic non-forest vegetation (GARDNER et al., 2009), with a consequent reduction of habitat size and increase of patch isolation, both of which conduce to a reduction in species richness (BIERREGAARD JR. et al., 1992). Fragmentation can induce a reduction of biological diversity in terms of mainly: (1) local extinction (HARRIS, 2013; SAUNDERS et al., 1991); (2) homogenization of regional biotas by the increase of invasive and/or synanthropic species (NOSS, 1983; TAKKIS et al., 2018); and (3) both processes acting together either independently or synergistically.
The extinction of native species is considered an unquestioned outcome of tropical forest fragmentation as several studies have shown a conspicuous decline in species richness over time within fragments of forest (BLANDÓN et al., 2016; DODSON & GENTRY, 1991; HADDAD et al., 2015; KATTAN et al., 1994; LECK, 1979). In addition, although there is always some level of ecosystem resistance to invasion, during the fragmentation process there is also a strong tide of colonization by invasive (JANZEN, 1983; JOSHI et al., 2015; SIMBERLOFF, 1992) and/or synanthropic species (TAKKIS et al., 2018). This colonization is promoted in part by a greater frequency of disturbances occurring in and around forest fragments (CHAPMAN et al., 2015; LAURANCE, 1991). Species invasion can be so overwhelming that in some cases an increment of species richness within a forest patch may be observed, but by an increase of invasive (NOSS, 1983) and/or of synanthropic (TAKKIS et al., 2018) species. Thus, at the scale of a single forest patch, the particular outcome of fragmentation in terms of species richness and composition depends on a compromise between extinction and immigration of species. Thus, the immediate effect of a fragmentation process is an addition of species to the recently formed forest fragments, most of which would be species that were already present in the landscape. But the species that were specific to the areas of forest that were subjected to a land cover transformation may vanish. The result is that while each forest fragment may gain species, the region as a whole will become less diverse (ANDREN, 1992).
The point in the fragmentation process at which the biological integrity of a regional pool of species declines abruptly depends on the guild of organisms considered, since it will be a function of parameters such as dispersal ability, sensitivity to isolation, and life span (i.e., lag between fragmentation and species loss may take far more time in some organisms than in others due to their longer life spans). An important additional factor would be the distribution pattern of individuals of different species across the region. Although contagion may constitute the most widespread distribution pattern in natural ecosystems, explained mainly by the assumption that organisms exhibit some habitat specificity (TUOMISTO et al., 2003), there is a considerable amount of variance in the degree of contagion, ranging from almost random, to completely clumped. This variability, which is scale dependent, is dictated by both the niche and the dispersal ability of each species. On one end of this continuum there are species tolerant to a wide array of environmental characteristics and with high dispersal abilities. Thus, they will appear as with a highly widespread or near random distribution within a geographic region. On the other end of the continuum there are species highly sensitive to environmental gradients, with lower dispersal rates, and which are then constrained to areas within a geographic region that exhibit only the most suitable environmental characteristics for their development, thus appearing with a clumped distribution. In many instances, when the environmental conditions are optimal, these species are highly competitive and resistant to invasion, but under disturbance, their competitive ability may be diminished.
Here we propose a procedure that can be used to evaluate the effects of tropical forest fragmentation on regional tree biodiversity based on two premises. First, if most of the species of a regional pool exhibit a clumped distribution within a human induced fragmented landscape (i.e., low inter-sample floristic similarity) then species composition prior to fragmentation could still be maintained. On the contrary, if most of the species of the regional pool show a random or near-random distribution (i.e., high inter-sample floristic similarity), then the fragmentation event could have resulted in a point in which there was a switch from a native species dominated landscape to one dominated by invasive and/or synanthropic species. Second, a fundamental concept of geographic analysis is the principle of distance-decay, which states that nearby environments are generally more similar to each other than distant ones (FOTHERINGHAM, 1981; FOTHERINGHAM et al., 2003; ROBERTSON, 1987). Thus, there is an inverse correlation between ecosystem similarity and distance among samples. In the case of floristic similarity this relationship also holds, although the slope of this relation is influenced by geographic scale and by species’ dispersal ability (SOININEN et al., 2007; TUOMISTO et al., 2003). Therefore, it is expected that a landscape dominated by invasive and/or synanthropic species should have a strong distance-decay function, since these species show a strong distance-decay. On the contrary, a less significant distance-decay function with lower slopes would be more a characteristic of a landscape dominated by native species (NEKOLA & WHITE, 1999). This paper presents the results of a vegetation survey performed in a highly fragmented lowland moist forest on the eastern foothills of the Eastern Andes of Colombia, and uses these premises (i.e., randomness of species distributions and distance-decay in floristic similarity) to evaluate the degree of disturbance, in terms of species composition, generated by forest fragmentation.
STUDY AREA
The study area is located in the eastern foothills of the Eastern Cordillera, Department of Casanare, Colombia, comprising portions of Charte, Cravo Sur and Payero river basins (5°30'N, 72°30'W; Figure 1). Elevation ranges between 300 and 800 m. At 300 m (Yopal weather station) mean annual rainfall and temperature are 2,508 mm and 26.4°C, respectively. There is a dry season between December and March when mean monthly rainfall is less than pan-evaporation. Following these temperature and precipitation regimes, the vegetation of the study area is classified as Lowland Tropical Moist Forest based on Holdridge’s life zone system (HOLDRIDGE, 1967). The area is geologically composed by sedimentary outcrops of the Tertiary and by alluvial deposits of the Quaternary (GOOSEN, 1971). Top soils are acidic (pH < 5.5), with exchangeable Al ranging from 2 to 4 meq/100 g. They show low contents of organic matter (< 2 %) and nitrogen (0.1-0.2 %), but normal C/N ratios (ca. 10). Available phosphorus is very low (ca. 5.7 ppm) and sodium values are below the salinity level (< 1 meq/100 g).
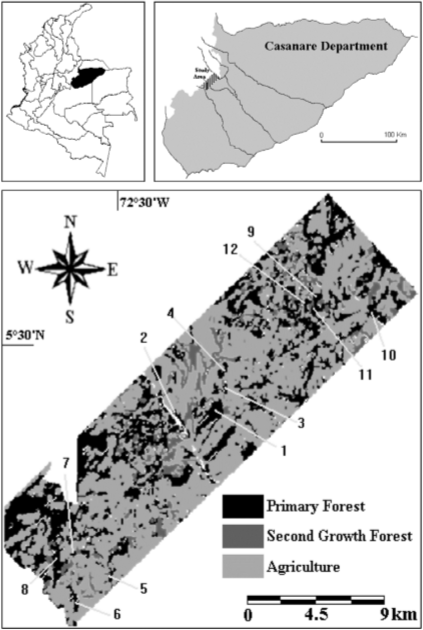
Figure 1 Map of the study area (270 km2 in the eastern foothills of the Eastern Cordillera of the Northern Andes, Department of Casanare, Colombia) showing the main land cover types and the location of each of the twelve forest patches studied (after VIÑA & CAVELIER, 1999). Numbers in the figure represent each of the forest patches evaluated (see Table 1 for a description of the structure of each patch).
The lowland Andean forests of the region once occupied a continuous narrow belt between the savannas of the Amazon and Orinoco river basins and the montane forests along the eastern Andes of Colombia and Venezuela. Therefore, the biota of the area may have some degree of similarity to that of the Amazon regions of Colombia, Ecuador, and Peru, and may also have some similarities with the foothills of the Venezuelan Cordillera of Merida (ZULUAGA-RAMÍREZ et al., 1994). Nevertheless, this conclusion is based on observations done to the north (Department of Arauca, Colombia) given that the biota of the study area has been little studied.
The study area has suffered an extensive deforestation process that has reduced the forest cover by more than 50% (VIÑA & CAVELIER, 1999). The continuous forest cover was replaced by a mosaic of forest patches of different sizes and shapes, surrounded by a matrix of pastures, croplands and secondary forests of different ages. While forest patches in the study area exhibit a high human influence, some of them have been maintained through several years after the fragmentation process, as could be seen on a time series of aerial photographs used to report the deforestation process in the region (VIÑA & CAVELIER, 1999). Thus, the study area constitutes a suitable place for studying tropical forest fragmentation and its effects on tree species composition.
METHODS
Forest structure
Forest structure and tree species composition were measured in twelve forest patches ranging from 4 to 120 ha and located in the Charte, Cravo Sur and Payero river basins (i.e., four patches per basin; Figure 1). Tree sampling was performed using the point-centered quarter method (COTTAM & CURTIS, 1956; COTTAM et al., 1953; ELLENBERG & MUELLER-DOMBOIS, 1974). This method was applied along transects of variable length, depending on forest patch size, and starting from a random point located 30 m in the interior of each forest patch, to avoid over-sampling edge species. Sampling points were located every 15 m along each transect, and for each of the four quadrants, the distance from the sampling point, the diameter at breast height (dbh) and a botanical sample of the nearest tree (> 2.5 cm dbh) were obtained. Vouchers were deposited at the herbarium of the Alexander von Humboldt Institute, Colombia. Unidentified taxa were classified as morpho-species. The distance between sampling points was deemed appropriate due to the tree density observed (shorter distances would produce an overlap between sampling points, while larger distances would leave many trees unsampled). To estimate tree density per forest patch, all distances from trees to sampling points along transects were averaged and squared to obtain the mean area occupied by each tree (ELLENBERG & MUELLER-DOMBOIS, 1974).
Numerical Analyses
Rarefaction curves show the relationship between a determined number of individuals sampled and the number of species expected for that sample size (HURLBERT, 1971; SANDERS, 1968). But an important conceptual restriction is that rarefaction assumes that species in a sample (e.g., forest patch) follow a random distribution (SMITH & GRASSLE, 1977). Thus, if the species have a clumped distribution, the observed number of species will be significantly lower than the expected number obtained by the rarefaction curve (HECK JR. et al., 1975; SMITH & GRASSLE, 1977). Therefore, we calculated a rarefaction curve using all stems sampled in all forest patches combined (i.e., as if the study area was not fragmented) and then compared the expected number of species obtained by this rarefaction curve against the observed number of species in each forest patch. The rarefaction curve (HURLBERT, 1971; SANDERS, 1968) and its variance (HECK JR. et al., 1975) were calculated following the expressions (1) and (2):
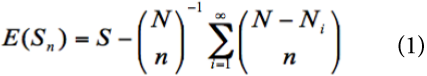
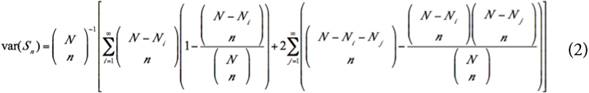
Where, E(S n ) is the expected number of tree species obtained in n individuals sampled, S is the total number of species observed, N is the total number of individuals sampled, N i is the number of individuals of species i, var (S n ) is the variance of the expected number of tree species, and N j is N i+1 .
To evaluate floristic distance-decay, inter-patch floristic similarity was calculated using the Jaccard index for presence-absence data (3) (LÔBO et al., 2011; OLDEN & ROONEY, 2006; QIAN & GUO, 2010):
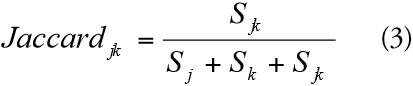
Where S jk is the number of species co-occurring in patches j and k, S j the number of species present in patch j, and S k the number of species present in patch k. The inter-patch Jaccard index was then plotted against the inter-patch geographic distance (on a logarithmic scale).
To simulate a landscape dominated by invasive and/or synanthropic species, both methods (i.e., rarefaction and plot of inter-patch similarity vs. inter-patch geographic distance) were applied after excluding rare species, or species found only within a single forest patch. Observed and simulated species distributions were then compared to evaluate the effects of fragmentation on species occurrence and distribution.
RESULTS
A total of 1308 individuals (> 2.5 cm dbh), belonging to 369 species and morphospecies in 59 families, were sampled (see Appendix). The most important family was Leguminosae with 73 tree species, followed by Euphorbiaceae (23 spp.), Rubiaceae (23 spp.), Myrtaceae (18 spp.), Lauraceae (14 spp.), Melastomataceae (14 spp.), Flacourtiaceae (13 spp.) and Moraceae (12 spp.). About 67% of the species were found only in a single forest patch (Figure 2; Appendix), with the notable exceptions of Urera caracasana (Jack) Griseb found in nine forest patches and Tabebuia rosea (Bertol) D.C. found in eight forest patches. Structural features of each of the forest patches studied are shown in Table 1. Average stem density was 1438 individuals per hectare and average basal area was 32.1 m2 per ha (Table 1).
Table 1 Structural characteristics of the forest patches sampled. The numbers in the ‘Forest Patch’ column correspond to the numbers shown in Figure 1.
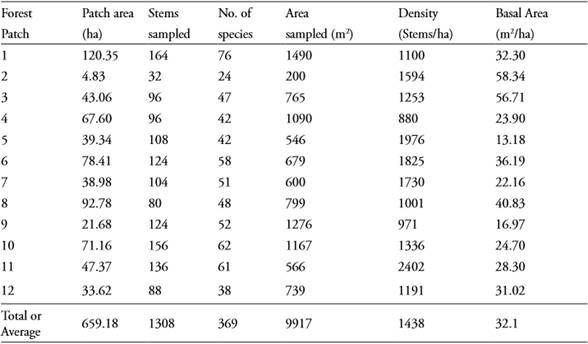
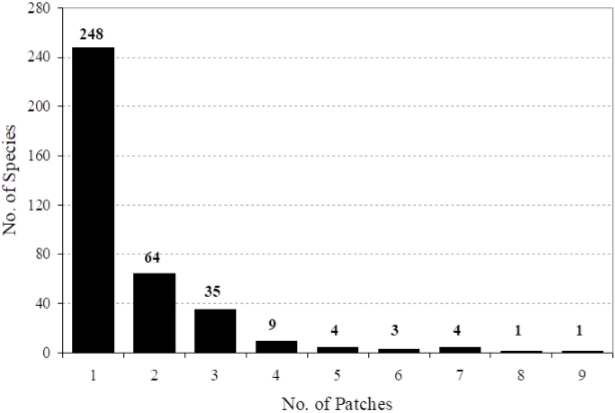
Figure 2 Frequency distribution of the number of tree species occurring within one or more forest patches. For instance, 248 species were only found within a single forest patch, while 64 were found within two forest patches.
The expected number of tree species per number of stems sampled, based on a rarefaction curve obtained for all the individuals sampled in all the forest patches combined, is shown in Figure 3a. The actual number of tree species obtained per forest patch is also shown. For any number of stems sampled, the observed number of tree species is lower in all forest patches than the expected by the rarefaction curve (Figure 3a). Thus, the tree community present in the twelve forest patches studied is dominated by species with a clumped distribution as opposed to the random distribution that would be expected if inter-patch floristic similarity was higher. Figure 3b shows the rarefaction calculated on the same data, but excluding 248 rare species or species found only within a single forest patch. In this case, the number of tree species observed per number of stems sampled is within the confidence intervals established by the expected number of tree species, showing that under this circumstance the tree community would be dominated by species with a more random distribution, thus having a higher inter-patch floristic similarity.
Distance-decay plots of floristic similarity for the entire dataset, and for the data excluding rare species (i.e., those found only in single forest patch) are shown in Figure 4. Both models are significant (p < 0.05), and explained 54-56% of the observed variance, but the slope of the distance-decay function obtained excluding rare species is almost two times steeper, thus exhibiting a stronger distance-decay.
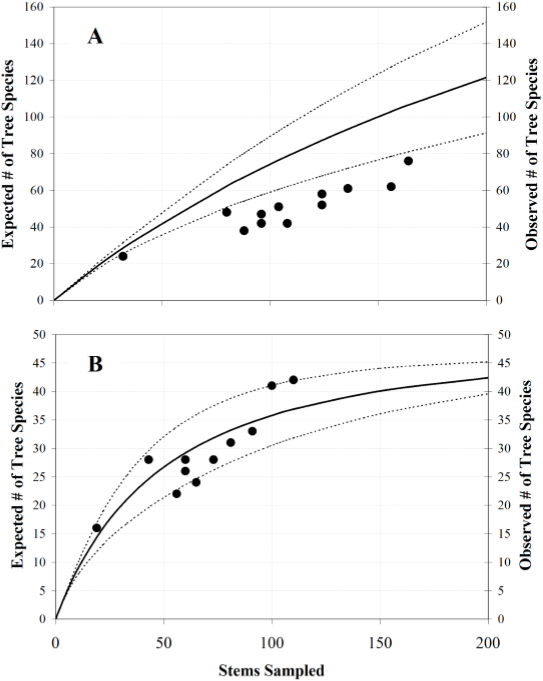
Figure 3 A) Rarefaction curve (equation 1) obtained for all the stems sampled in all the forest patches combined (continuous line), and the observed number of species sampled per patch (black dots). (B) Similar to Figure 2A, but calculated excluding 248 rare species (i.e., those occurring only in one forest patch). Dotted lines in both figures represent the rarefaction variance (equation 2).
Figure 4 Distance-decay plots of floristic similarity (Jaccard Index; equation 3) for both the entire dataset (black dots; continuous line; y = -0.0379Log(x) + 0.2058; r2 = 0.555; p < 0.05) and the dataset excluding 248 rare species (i.e., occurring only in one forest patch; white squares; dotted line; y = -0.0585Log(x) + 0.3377; r2 = 0.539; p < 0.05).
DISCUSSION
An important ecological change that might be expected via the transformation of a once continuous tropical forest into a patchwork of forest fragments is the regional tree species homogenization (BIERREGAARD JR. et al., 1992; LIU et al., 2018; NOSS, 1983). While richness per patch may increase due to the colonization of invasive and/ or synanthropic species (TAKKIS et al., 2018), floristic similarity among patches is also expected to increase. Thus the overall species diversity of the entire region would be reduced. This outcome was expected in our study area as its forest patches are the result of a deforestation process that has influenced the entire study region (VIÑA & CAVELIER, 1999). When the number of tree species was reduced so that only the species that occurred in two or more forest patches were included (i.e., 121 spp. or about one third of the total number of species), the results of both, rarefaction and distance-decay of floristic similarity correspond to the regional homogenization of species composition that would be expected for a fragmented ecosystem. But without the reduction of rare species the study area showed the characteristics of a landscape dominated by native species, i.e., with most tree species sampled displaying a clumped distribution, and a low inter-patch floristic similarity.
These results suggest that pre-fragmentation tree species composition may still be maintained, and an equilibrium point in which all extinction-prone species become regionally extinct may have not yet been reached (MCCOY, 1982). However, the point in a fragmentation process at which the biological integrity declines abruptly is usually not known, and in the case of tree species the lag between fragmentation and species loss may take long periods of time due to their longer life spans. In the study area, two thirds of the tree species sampled were considered rare, since they were found only within a single forest patch among the twelve evaluated. Thus, some of these tree species may be prone to regional extinction if forest fragmentation is maintained or increased. This may be the case of dioecious (i.e., male and female flowers on different plants) or of self-incompatible hermaphroditic (i.e., bisexual flowers) tree species, as has been reported for riparian tropical rain forests in Belize (MEAVE & KELLMAN, 1994) and for tropical dry forest patches in Costa Rica and Nicaragua (GILLESPIE, 1999). This could mean that the study area may still lose several of its tree species given that severely fragmented tropical forests are considered 'living dead' because many of their tree species may eventually become extinct as co-adapted pollinators and seed dispersers, which are quite sensitive to forest fragmentation, may become locally extinct (DE PAULA MATEUS et al., 2018; JANZEN, 1986). However, more research on this regard is needed to evaluate the species composition of these co-adapted wildlife species in the study region.
The analysis of inter-patch floristic similarity shown in this study constitutes a practical way to evaluate the effects of tropical forest fragmentation on biodiversity at regional scales, and is applicable to many other fragmented forest areas around the tropics. However, the premises used in this study to evaluate the effects of forest fragmentation on tree species composition (i.e., randomness of species distributions and distancedecay in floristic similarity) assume that geographic space is homogeneous, while geographic barriers may contribute to a discontinuous occurrence of species, although depending on how effective these barriers are in reducing species dispersal. Thus, the expansion of invasive and/or synanthropic woody species could be diminished or even prevented by the presence of extensive areas of pastures and other non-forest land covers that may constitute important dispersal barriers. However, as several species exhibited a broad distribution within the study area [e.g., Urera caracasana (Jack) Griseb, Tabebuia rosea (Bertol) D.C.], these non-forest areas may not yet constitute important dispersal barriers. In fact, the study area exhibits significant forest regeneration, as second growth forests constitute an abundant land cover type [ca. 15% of the entire study area was under this land cover class (VIÑA & CAVELIER 1999); Figure 1]. This also supports the notion that distance to seed sources may not yet constitute important barriers to forest regeneration in the study area, as opposed to what has been observed in other highly fragmented forests of the Neotropics (NEPSTAD et al., 1990). Therefore, the study of the dynamics of forest regeneration becomes a priority for conservation and restoration programs in the region. Thus, future assessments of the number and composition of tree species colonizing these areas of forest regeneration are crucial for understanding long-term effects of tropical forest fragmentation on tree species composition, as has been suggested by some authors (BROWN & LUGO, 1990; CHAZDON, 1999; LUGO, 1992). These tropical secondary lowland forests have been shown to carry out a very important function as preservers of soil moisture and nutrients in deforested areas, as regulators of biogeochemical cycles, and as refuges of animal and plant species, among many others (BROWN & LUGO, 1990; CHAZDON, 1999; DEL PLIEGO et al., 2016).
A major limitation of this study was the use of only a few forest patches to evaluate the effects of forest fragmentation on tree species occurrence and distribution, therefore the conclusions drawn may need to be taken as preliminary. Nevertheless, the study presents useful information that will hopefully stimulate further research on the linkages between deforestation and fragmentation of tropical forests and their effects on biodiversity not only within the study region but throughout the Tropics.


