











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Los estudios de abejas en todo el mundo tienden a centrarse en zonas de alta diversidad biológica, sin embargo, la influencia del hombre sobre los ecosistemas es cada vez mayor y existe la necesidad de evaluar diferentes aspectos de la biología de las abejas en zonas urbanas con el fin de proponer alternativas que eviten la desaparición de las especies nativas (NATES-PARRA et al., 2006a).
El desarrollo urbano afecta directamente la fauna local debido a que se produce una pérdida de hábitat y fragmentación del paisaje, esto tiene serias consecuencias para la diversidad local pues se generan cambios permanentes en el medio ambiente (MARKOVCHICK-NICHOLLS et al., 2008). En el caso de las abejas, dependiendo de las especies estos ambientes pueden beneficiarlas, pues generan nuevos sitios de nidificación o afectarlas al invadir su hábitat o por efecto de la contaminación atmosférica (NATES-PARRA et al., 2006a; MARTÍNEZ et al., 2017).
lLas abejas cumplen un papel ecosistémico muy importante debido a que son los insectos que están más implicados en la polinización y pueden presentar preferencia en cuanto a los recursos que colectan como el polen, el néctar o diferentes tipos de sustancias resinosas (OTERO et al., 2014); sumado a esto, las abejas sociales son más eficientes facilitando la polinización de plantas que las abejas solitarias, debido a su sistema de comunicación, el alto número de individuos por colonia y una mayor efectividad en el acopio del alimento (NATES-PARRA 1991a; NATES-PARRA & RODRÍGUEZ 2011; OTERO & SANDINO, 2003).
Las abejas angelitas (Apidae: Meliponini) son abejas sociales, distribuidas entre el nivel del mar y los 2000 m (NATES-PARRA, 1990), se distinguen porque tienen su aguijón atrofiado no funcional, alas con la venación reducida, y porque nidifican en gran variedad de sustratos, ya sea en cavidades o de forma expuesta. En Colombia puede encontrarse aproximadamente unas 100 especies distribuidas en todas las regiones naturales (NATES-PARRA, 2001). Sin embargo, y aunque las abejas meliponinas son quizá uno de los grupos más diversos y abundantes entre las abejas sociales de Colombia (NATES-PARRA, 1991a), muchas especies están amenazadas, debido entre otras cosas a la presencia de abejas africanizadas, al uso de pesticidas y a la desaparición de zonas boscosas (NATES-PARRA, 1991b).
En ambientes naturales como bosques tropicales, sabanas, bosques subtropicales, bosques de araucarias y también en comunidades modificadas con jardines, especies ornamentales, árboles frutales entre otros, algunas especies de meliponinas toman recursos de especies dentro de las familia Myrtaceae, Melastomataceae, Solanaceae y de la subfamilia Mimosoidea, debido a que pueden llegar a producir abundante polen y néctar, supliendo así sus necesidades nutricionales (RAMALHO et al., 1989); sin embargo, los hábitos alimenticios generalistas y la explotación oportunista de alimentos pueden conducir a una utilización generalizada de los recursos en zonas con baja oferta floral, en parches de bosques o variar dependiendo de los períodos de floración (RAMALHO et al., 1989).
En Colombia se han realizado trabajos en los que se busca conocer las preferencias de los tipos de polen recolectado por parte de algunas especies de meliponinos. OBREGÓN (2011) estudió el origen botánico de la miel y el polen de Meliponae eburnea Friese 1900, y Tetragonisca angustula (LATREILLE, 1811) en Fusagasugá (Cundinamarca), encontrando que M. eburnea colecta en gran medida polen de especies vegetales de las familias Myrtaceae, Melastomataceae, Solanaceae y Euphorbiaceae; además que presentan una preferencia por especies nativas pese a la presencia de plantas foráneas en la zona. RODRÍGUEZ y NATES-PARRA (2006) encontraron que Melipona fasciata Latreille 1811, colecta principalmente Myrtaceas (Psidium guajava L.), Melastomataceae, Solanaceae y Caesalpinaceae en el municipio de Acacias (Meta). Sin embargo, los estudios a este grupo taxonómico se centran a investigaciones dirigidas por la Universidad Nacional de Colombia en Bogotá, a cargo del laboratorio de abejas (LABUN) (NATES-PARRA & GONZALES, 2000). Muchos de los estudios en el Valle del Cauca han priorizado los censos y distribución geográfica del material colectado (NATES-PARRA, 1984, 1990, 1991a). Por lo que resulta de gran importancia generar conocimiento en el departamento del Valle acerca de los hábitos de forrajeo en las especies de abejas nativas.
El conocer las plantas de las cuales dependen estas abejas como fuente de recursos, resulta fundamental para mantener las interacciones planta-insecto, pues brinda herramientas para su conservación y valoración como polinizador nativo (OBREGÓN, 2001). Así pues, el objetivo del presente trabajo es aportar información que permita reconocer las plantas utilizadas como fuentes de polen y los hábitos en cuanto a la recolección de recursos polínicos por parte de Nannotrigona mellaria (Smith, 1862), una especie común en el departamento y que se encuentra fácilmente en zonas urbanas; con ello se espera brindar datos que faciliten su conservación o su uso en la meliponicultura, además de aumentar su valor como polinizador.
MATERIALES Y MÉTODO
Sitio de estudio
El estudio se realizó en la ciudad de Cali, Valle del Cauca, en el campus de la Universidad del Valle Sede Meléndez, 3°22′ N -- 76°31′ W, altura aproximada de 995 m., zona de vida de bosque seco tropical según el sistema de clasificación de HOLDRIDGE (1967). La vegetación incluye alrededor de 4250 árboles y unas 182 especies (HERRERA-HURTADO, 2009). A dicha vegetación deben sumarse las especies herbáceas, epífitas y arbustivas, una representación muy valiosa del paisaje local y todas fuentes posibles de néctar y polen para las abejas que son objeto de estudio. También se realizará colectas en el campus de la Universidad Nacional sede Palmira en las coordenadas geográficas 3°30’45” N 76°18’29.9” W, a una altura aproximada de 965 m.
Se reportaron un total de 242 colonias de N. mellaria (Figura 1) en la Universidad del Valle de las cuáles se seleccionaron tres (Nido A, Nido B y Nido D) ubicadas a más de 500 m entre ellas; en la Universidad Nacional de Colombia sede Palmira se reportaron un total de 21 colonias y se seleccionó una de forma aleatoria (Nido C); la especie fue determinada y depositada en la colección del laboratorio de investigaciones en abejas (LABUN) con los números de catálogo 25274, 25275, 25276 y 25277. Se recolectó el polen corbicular de 20 individuos que regresaban al nido luego del pecoreo, durante 24 semanas una vez a la semana por nido, entre las 7 am y las 10 am, debido a que este es el horario de mayor actividad de estas abejas. Las recolecciones se realizaron desde junio de 2013 hasta enero de 2014.

Figura 1 Nannotrigona mellaria en la entrada de uno de los nidos utilizados para la recolección de muestras.
El material colectado se guardó en tubos Eppendorf individuales de 1,5 mL con ácido acético glacial para su posterior montaje. Adicional al polen corbicular, se realizaron colectas de polen de plantas en flor a 250 m de radio de las colonias estudiadas; el material se conservó en ácido acético glacial hasta su posterior tratamiento en laboratorio. Las plantas de las que fue obtenido el polen fueron identificadas por comparación en el herbario Luis Sigifredo Espinal Tascón (CUVC) de la Universidad del Valle.
Tratamiento en laboratorio
El polen recolectado de las abejas fue centrifugado en ácido acético a 3000 rpm durante 4 min, se descartó el sobrenadante y se lavó en tubos Eppendorf de 1,5 mL con 1,4 mL de agua destilada; luego se centrifugó nuevamente y se repitió el lavado con el fin de limpiar el ácido acético previo al montaje. En cada fase de lavado se adicionó una gota de etanol absoluto con el fin de facilitar el hundimiento de los granos de polen. Finalmente, luego del lavado el material se secó en horno a 40 °C durante al menos dos horas.
El polen colectado de las plantas fue acetolizado según el método presentado por FONNEGRA (1989), con algunas modificaciones. Se realizó las preparaciones con un volumen máximo de 1,5 mL; una vez tratado, el polen fue montado con gelatina glicerinada la cual fue preparada según las especificaciones de FONNEGRA (1989) con las cantidades para la Universidad de Antioquia; las placas fueron selladas con parafina y para el polen acetolizado se montó al menos dos placas por muestra (OTERO et al., 2014).
Análisis de polen
El polen obtenido de las corbículas de las abejas, fue identificado por comparación con el material recolectado de las plantas alrededor de las colonias, mediante el uso de un microscopio óptico Nikon E-200. Adicional a esto, se comparó con láminas y descripciones de los libros Pollen morphology and plant taxonomy: Angiosperms (ERDTMAN, 1986) y Pollen and spores of barro colorado (ROUBIK & MORENO, 1991). Los datos que se guardaron corresponden al número de veces que se reporta el tipo de polen de una especie vegetal en el material obtenido semanalmente por nido.
Se estimó la riqueza de especies vegetales colectadas por las abejas en los diferentes nidos y la eficiencia de muestreo mediante el uso de curvas de acumulación de especies, con los estimadores de riqueza Chao 1, Chao 2, Jack 1 y Jack 2; se utilizaron estos estimadores debido a que presentan un mejor desempeño (WALTHER & MOORE, 2005) para estimar el número de especies vegetales utilizadas por Nannotrigona mellaria como fuentes de polen dentro del área de estudio. Estos análisis se realizaron mediante el uso del software informático EstimateS Win910 (COLWELL, 2013).
Se realizó un análisis de agrupamiento UPGMA con índice de similitud Bray-Curtis de los datos obtenidos durante las 24 semanas por nido, sumando las abundancias por especie vegetal del material colectado, con el fin de determinar si las colonias estudiadas presentan polen de especies vegetales afines y que tan similares son las colonias entre sí. Estos análisis se realizaron mediante el uso del programa estadístico PAST 3.0.1 (HAMMER et al., 2001).
Finalmente, se realizó un análisis de especie indicadora mediante el uso del programa estadístico PC ORD 6 (MCCUNE & MEFFORD, 2011), para lo cual se sumaron los datos obtenidos cada cuatro semanas por nido, con el fin de interpretar más fácilmente los resultados (6 datos en total por nido). Este análisis se realizó para determinar cuáles son las especies específicas para cada nido estudiado.
RESULTADOS
Polen recolectado por Nannotrigona mellaria
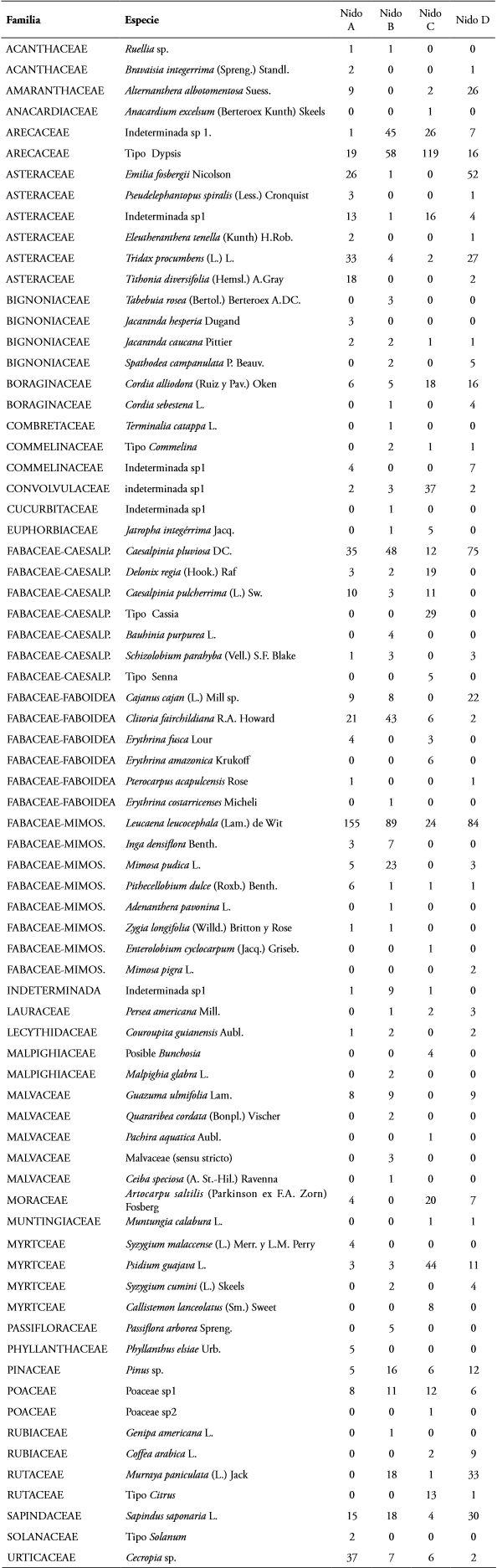
Se encontró que N. mellaria utiliza al menos 73 especies vegetales como fuente de polen para los cuatro nidos estudiados, tales especies se encuentran dentro de 29 familias, siendo Fabaceae la más representada; además, se reportó un tipo polínico que no pudo ser identificado ni siquiera a nivel de familia (Figura 2). El polen de Leucaena leucocephala (Lam.) de Wit, fue el que se encontró más frecuentemente durante el tiempo de estudio. En la tabla 1 se presentan las veces que fue reportada cada especie para cada nido. Los tipos de polen más representativos se presentan en la figura 3.
Tabla 1 Tipos de polen por familia y especie vegetal recolectados por N. mellaria en los nidos estudiados.
Eficiencia de muestreo
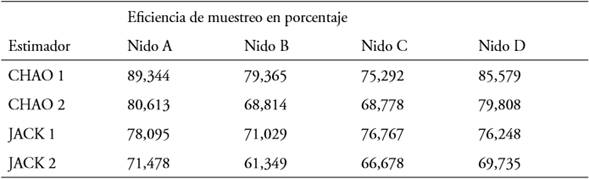
En el nido B se encontró un mayor número de taxa, 47 tipos diferentes de polen y el nido C fue en el que se encontró el menor número, 38 tipos. Los muestreos realizados presentan una eficiencia mayor para el nido A (hasta el 89% Chao 1) y una menor eficiencia para el nido C (hasta 76,767% Jack 1) (Tabla 2).
Similitud entre los nidos estudiados
El análisis de agrupamiento indica que las obreras en los nidos de la Universidad del Valle (A, B, D) recogen tipos de polen más similar entre sí, que con el nido C, que se encontraba en la Universidad Nacional. Entre los nidos de la Universidad del Valle se comparte más del 50% en similitud, mientras que estos con el nido de la Universidad Nacional comparten alrededor del 30% (Figura 4).
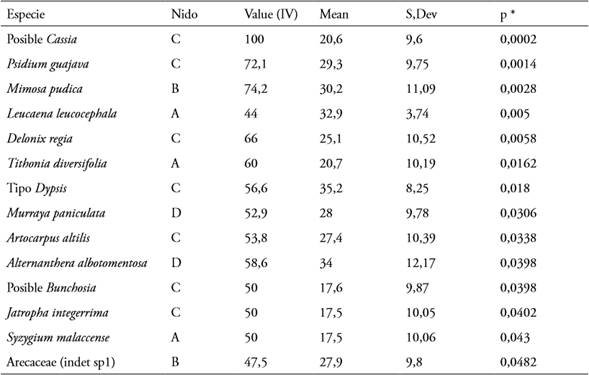
En cuanto a los tipos de polen, 14 de las especies vegetales encontradas mostraron un valor indicador significativo (<0,05) para los nidos estudiados; tres especies resultaron indicadoras para el nido A, dos especies para el nido B, siete especies para el nido C y dos especies para el nido D; los resultados del análisis se presentan en la tabla 3.
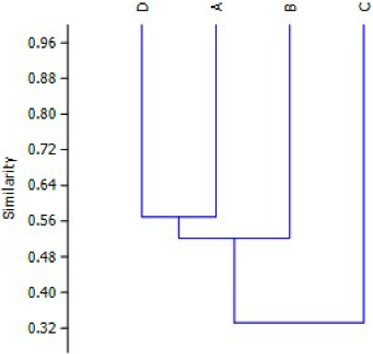
Figura 4 Análisis de agrupamiento UPGMA con índice de similitud Bray Curtis para los cuatro nidos estudiados.
DISCUSIÓN
La especie Nannotrigona mellaria exploró un amplio rango de plantas en el pecoreo de polen; según las muestras obtenidas, utilizó 73 especies pertenecientes 29 familias durante el tiempo de estudio, sin embargo, explotan principalmente unas pocas especies entre las que se destacan Leucaena leucocephala y polen de Arecaceae tipo Dypsis con el 18% y el 11% de las presencias en el material colectado, respectivamente. Algunos estudios realizados con otras especies de meliponinos muestran que estas abejas colectan en gran medida polen de árboles (ROUBIK et al., 1986; WILMS et al., 1996; RAMALHO, 2004), entre los que se pueden resaltar Eucalyptus sp., Eucaliptus globulus Labill., polen del tipo Myrcia (MALAGODI-BRAGA & KLEINERT, 2009; OBREGÓN, 2011); en nuestro caso particular esta tendencia se mantiene pues N. mellaria recolectó polen principalmente de árboles dentro de la familia Fabaceae debido posiblemente a que estos árboles ofrecen una gran cantidad de inflorescencias en un área relativamente pequeña por lo que las abejas no tienen que desplazarse mucho y son más eficientes en el acopio de recursos.
En cuanto a la especie arbórea, Leucaena leucocephala hay algunas características que podrían explicar el por qué es una fuente tan importante de recursos nutricionales para N. mellaria durante el tiempo de muestreo; para empezar esta una especie relativamente abundante en los sitios de estudio por lo que es muy accesible a las obreras de estas abejas, adicional a esto el periodo de máxima floración de esa especie va desde julio hasta octubre (PÉREZ et al., 2008), tiempo en el cuál se estaba realizando la recolección del polen corbicular, finalmente el alto número de inflorescencias y flores por inflorescencia puede explicar el por qué el polen de esta especie fue tan frecuente en el material recolectado para los cuatro nidos. Esto datos contrastan con las preferencias de otros Apidae como los Euglossinos, puesto la mayoría de las fuentes de polen son plantas con anteras poricidas como solanáceas y melastomatáceas (OTERO et al., 2014), familias que también son muy colectadas por meliponinos (WILMS & WIECHERS, 1997; OBREGÓN, 2011; VIT et al., 2013).
Muchos visitantes y polinizadores suelen ser atraídos a flores que presentan algunas características particularmente atrayentes tales como una coloración llamativa (MENZEL & SHMIDA, 1993; GUMBERT, 2000) o fragancias (WHITTEN & WILLIAMS, 1992; GERLACH, 2010), debido a que estas plantas pueden ofrecer algún tipo de beneficio o recompensa, es decir, son “magnet species” (THOMPSON, 1978). Sin embargo, la presencia de éstas puede favorecer la visita a plantas con flores menos atractivas que se encuentren cercanas u asociadas (THOMSON, 1978; FEINSINGER, 1987; JOHNSON et al., 2003; MOLINA et al., 2008).
En estudios realizados por VARGAS (2012), en bosque seco del Valle del Cauca indica que la familia Fabaceae es una de las más diversas con 37 géneros; del mismo modo, en la Universidad del Valle esta familia es una de las más abundantes, con un total de total de 35 especies solamente en árboles (VARGAS et al., 2011). En general, muchas de las plantas dentro de esta familia presentan inflorescencias muy vistosas y llamativas que resultan muy atractivas a los polinizadores, además ofrecen tanto polen como néctares que son los principales recursos alimenticios en Apidae (VIT et al., 2013); estas características atractivas para las abejas explicarían el que las Fabaceae sean tan importantes en las recolecciones de polen de N. mellaria.
Los resultados reportados en la eficiencia de muestreo indican que este fue adecuado pese a que las recolectas de polen se realizaron en un periodo de tiempo que corresponde aproximadamente a medio año; probablemente muestreos realizados en el periodo entre enero y junio permitan obtener una mayor aproximación al comportamiento en cuánto de pecoreo de polen por N. mellaria. Sin embargo, los resultados obtenidos en cuanto a la riqueza de especies son satisfactorios, teniendo en cuenta que trabajos realizados con Nannotrigona testaceicornis Lepeletier en Brasil, reportaron un total de 107 tipos de polen durante un año de estudios (MALAGODI-BRAGA & KLEINERT, 2009) y en Colombia, estudios con Apis melífera L., reportan el uso de 85 especies vegetales (MONTOYA, 2011), y en meliponinos con M. eburnea y T. angustula 93 y 81 especies vegetales respectivamente (OBREGÓN, 2011).
El análisis UPGMA con índice Bray-Curtis agrupa los nidos A, B, D con una similitud entre sí mayor al 50% y separados del nido C, esto indica durante el tiempo de estudio existe una preferencia específica por parte de N. mellaria a algunas especies vegetales como fuentes de polen, como ya se ha reportado en otros trabajos con meliponinos (WILMS & WECHERS, 1997; MALAGODI-BRAGA & KLEINERT, 2009; OBREGÓN, 2011; RECH & ABSY, 2011). Sin embargo, la recolección por parte de estas abejas también se ve influenciada por la riqueza y abundancia de dichas plantas en su medio ambiente circundante.
RAMÍREZ & MONTENEGRO (2004) reportaron que, en condiciones de abundancia de recurso, algunas abejas permanecen cerca de sus colmenas visitando las plantas que son más abundantes en el área. Es posible que las obreras de N. mellaria, en los nidos estudiados, utilizaran principalmente las plantas más cercanas a sus colonias como fuentes de polen, prefiriendo las plantas con flores más atractivas como se mencionó anteriormente. Esto explicaría los resultados obtenidos en el análisis de similitud.
Respecto al análisis de especie indicadora, los resultados obtenidos para el nido C pueden deberse a que éste se encontraba en condiciones ambientales que son diferentes, tanto en abundancia como riqueza de especies vegetales a los otros tres nidos. Así pues, estos factores pueden ser determinantes para N. mellaria a la hora de recolectar sus recursos más eficientemente (MONTOYA, 2011).
Para el nido A se reportó polen de L. leucocephala, Tithonia diversifolia y S. malaccense como especies indicadoras, vale la pena mencionar que todas ellas se encontraban a no más de 200 m de distancia del nido de manera abundante, de hecho, durante los muestreos se observó que T. diversifolia tenía una importante población en las cercanías del nido A, algo que no ocurría cerca de ninguno de los otros nidos, lo que parece indicar que las colonias muestreadas no necesitan recorrer grandes distancias en el pecoreo de recursos y que colectan principalmente en las cercanías, siempre que existan recursos suficientemente abundantes en la zona.
En conclusión, Nannotrigona mellaria es una especie nativa con gran potencial como polinizador en zonas urbanas del Valle de Cauca debido a que tolera la presencia humana alrededor de su nido, puede utilizar edificaciones como refugios para nidificar (NATES-PARRA et al., 2006b; MARTÍNEZ et al., 2017) y visita una gran cantidad de especies vegetales muy comunes en este tipo de zonas. De acuerdo con los resultados obtenidos se puede concluir que la familia Fabaceae es de gran importancia como fuente de polen para esta especie en la zona de estudio, probablemente debido a su abundancia, a su floración que es muy exuberante y atractiva visualmente a los Apidae y a que puede ofrecer néctares además de polen como alimento para la especie.

