











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La subtribu Pronophilina (Lepidoptera: Nymphalidae: Satyrinae) (Lamas, 2003; Viloria 2007; Viloria, Pyrcz y Orellana 2010), es uno de los grupos más diversos dentro de las mariposas diurnas en los paisajes de alta montaña en la región Neotropical (Adams, 1973; Álvarez-Hincapié et al., 2017; Pyrcz et al., 2016), en el cual se reconocen aproximadamente 45 géneros y más de 600 especies (Lamas, 2003; Pyrcz et al., 2011, 2020) de las cuales el 95% se encuentran principalmente en los Andes tropicales entre Venezuela y el norte de Argentina (Pyrcz, 2004; Viloria et al., 2010). En Colombia, se conocen cerca de 205 especies de pronofilinas, siendo el segundo país con la mayor riqueza de especies de este grupo, solo superado por Perú con un total de 284 especies (Pyrcz y Rodríguez, 2007, Mahecha-Jiménez et al., 2011).
Estas mariposas presentan patrones de distribución restringida especialmente en macizos montañosos aislados, además de numerosos complejos de especies crípticas que incluyen notables endemismos en las diferentes formaciones montañosas de la región Neotropical, mas especialmente en la región Andina tropical (Adams 1973; 1985, 1986; Greeney et al., 2009; Pyrcz et al., 2009; Viloria et al., 2010; Pyrcz et al., 2016; Mahecha et al., 2019). En adición, la mayoría de las especies de este grupo se restringen a lo largo de gradientes de altitud y bandas de elevación estrechas y bien definidas (< 300 metros) donde pueden encontrarse especies parapátridas (Pyrcz y Wojtusiak, 2002; Pyrcz y Rodríguez, 2007; Pyrcz et al., 2009; Viloria et al., 2010; Pyrcz y Garlacz, 2012; Pyrcz et al., 2016). La mayor cantidad de especies de Pronophilina se encuentra relacionada con ecosistemas templados húmedos o ecosistemas fríos tales como bosques andinos nublados, altoandinos y páramos, donde hay buena disponibilidad de sus plantas huéspedes (para el desarrollo de los estadios inmaduros) y nutricias (fuente de alimento) conocidas comúnmente como chusque o bambú de montaña (Chusquea serrulata y Chusquea scandens) y otras gramíneas (Montero y Ortiz, 2012, 2013).
El acelerado aumento de las actividades humanas que transforman y reducen los bosques naturales en la región andina colombiana por el incremento de cultivos, ganadería, urbanización (Clavijo-Giraldo et al., 2020), incluyendo las afectaciones ambientales como la pérdida de hábitat por el establecimiento de cultivos ilícitos y la presencia grupos armados sumado a la minería ilegal producto del conflicto armado en Colombia (Ríos-Málaver, 2019), están generando una constante reducción de la biodiversidad, incluyendo las especies nativas de mariposas en las regiones montañosas Neotropicales (Marín et al., 2014; Ríos-Málaver, 2019; Ríos-Málaver, Olarte-Quiñonez y Viloria 2020). Sin embargo, en la región nororiental andina colombiana se ha demostrado que algunas alteraciones antrópicas dentro de la estructura del paisaje, favorecen la heterogeneidad ambiental y con ello, la diversidad de especies de mariposas en ecosistemas de alta montaña (Olarte-Quiñonez et al., 2016). Más recientemente Acevedo et al. (2018) identificaron zonas de concentración de especies de mariposas y ranas entre las unidades biogeográficas, en los departamentos de Santander y Norte de Santander, a través de modelos de distribución potencial, resaltando la importancia de las áreas de alta montaña como un continuo de condiciones biogeográficas para mantener puntos de diversidad de estos grupos biológicos.
El estudio de los patrones de distribución y diversidad de mariposas es importante ya que refleja los cambios ecológicos que sufre una comunidad de insectos a medida que incrementa un gradiente a nivel altitudinal, mostrando distribuciones restringidas y endémicas en las montañas altoandinas (Pyrcz et al., 2009; Meléndez-Jaramillo et al., 2019). Por tal motivo, el objetivo del presente trabajo fue conocer la estructura, composición y los patrones de diversidad del ensamblaje de mariposas de la subtribu Pronophilina, en un gradiente altitudinal en el Cerro de Tierra Negra, municipio de Pamplona, Norte de Santander, de la Cordillera Oriental de Colombia.
Materiales y Métodos
Área de estudio
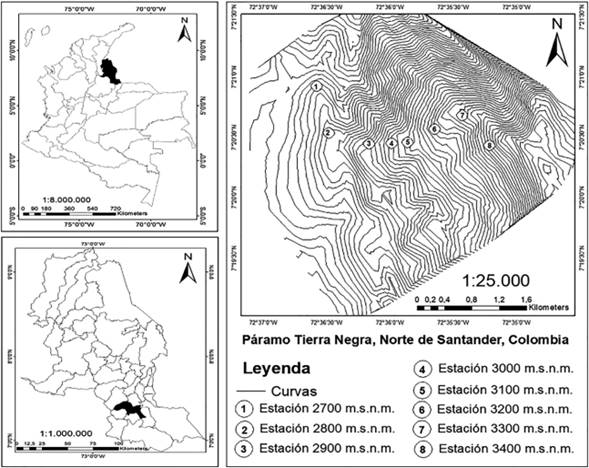
El Cerro de Tierra Negra se encuentra en el complejo de páramos de Santurbán, entre los departamentos de Norte de Santander y Santander (Rangel-Ch., 2000). Esta zona cuenta con una franja altitudinal de 2.678 a 3.600 msnm, con coordenadas 7°28’0” N y 72°34’60” O, con temperaturas medias entre 0,3 y 12°C y precipitación anual entre 600-2.500 mm (Morales et al., 2007). El área presenta fragmentos de bosque altoandino con dominancia de estados sucesionales secundarios en las zonas de 2.600 a 3.000 msnm (Holdridge, 2000; Rangel-Ch., 2000; Rodríguez et al., 2006) y presencia abundante de bambú de montaña Chusquea spp. En las zonas de páramo de 3.100 a 3.600 msnm presenta vegetación arbustiva con bosque bajo (Rodríguez et al., 2006; Morales et al., 2007) (Figura 1).
Recolección y determinación de las mariposas
El muestreo de campo se realizó a lo largo de seis meses, entre marzo y septiembre de 2013. Se recolectaron los adultos de mariposas diurnas a lo largo de un gradiente altitudinal entre 2.700 y 3.400 msnm (Pyrcz et al., 2009; Mahecha et al., 2019). Dentro del gradiente de elevación se establecieron ocho estaciones altitudinales fijadas mediante GPS (Garmin Rino 530HCX Handheld), altímetro calibrado y mapa topográfico del área de estudio del Instituto Geográfico Agustín Codazzi, distanciadas entre sí por un desnivel de 100 metros. Para la captura de individuos se instalaron 4 trampas tipo Van Someren-Rydon (TVS) (Rydon, 1964, Freitas et al., 2014; 2021) por estación altitudinal para un total de 32 trampas, se utilizó como atrayente pescado en descomposición (trucha arcoíris Oncorhynchus mykiss, comúnmente comercializada en la ciudad de Pamplona). Estas trampas fueron distancias 50m entre sí a una altura entre 1 y 1.5m del suelo (Freitas et al., 2006, 2014, 2021). La recolección de ejemplares fue realizada por una persona mediante el método activo de red entomológica manual y pasivo mediante el uso de trampas tipo Van Someren-Rydon (TVS) (con cebos atrayentes de pescado en descomposición).
La captura manual con red entomológica se realizó en dos transectos de 100 x 4 metros por estación, con recorridos diarios entre las 9:00 y las 16:00 horas para aprovechar la mayor cantidad de horas luz y cubrir diferentes hábitos de vuelo en los diferentes grupos de mariposas (Ríos-Málaver, 2007; Carrero-S. et al., 2013; Van Swaay et al., 2015; Checa et al., 2018); para un esfuerzo de muestreo de ocho horas diarias y un total de 256 horas durante 32 días.
Los individuos recolectados fueron depositados en sobres de papel pergamino de 90 g, en los cuales se registraron los datos de recolección con la fecha, altitud, hora y método empleado en la captura según las especificaciones de Neild (1996) y Chacón y Montero (2007).
El material recolectado se depositó en el Laboratorio de Entomología de la Universidad de Pamplona bajo el permiso de recolección de material biológico con fines de investigación científica no comercial, resolución 200 del 17 de abril de 2015, expedida por La Corporación Autónoma Regional de la Frontera Nororiental (CORPONOR). La determinación taxonómica se realizó con ayuda de descripciones originales y obras ilustradas: D’Abrera (1988); Pyrcz y Viloria (2007) y por medio de la comparación con ejemplares tipo de la base de datos en línea Butterflies of America (Warren et al., 2016) y muestras de la colección de referencia depositadas en la Colección Entomológica de la Universidad de Pamplona-CEUP.
Análisis de datos
Se obtuvieron los porcentajes de abundancia y riqueza por cada estación altitudinal, adicionalmente se realizó análisis de completitud para determinar el esfuerzo de muestreo mediante el índice propuesto por Chao y Jost (2012) con el paquete estadístico iNEXT-RStudio (Hsieh et al., 2013). Se realizaron los perfiles de diversidad alfa con los valores de diversidad de orden 0 (Q0), o riqueza de especies, diversidad de orden 1 (Q1), el cual representa el exponencial de la entropía del índice Shannon (Moreno, 2001; Marín et al., 2014) y la diversidad de orden 2 (Q2), que es el inverso del índice Simpson (Pyrcz et al., 2009; Moreno et al., 2011; Rös et al., 2012; Marín et al., 2014; Olarte-Quiñonez et al., 2016; Casas-Pinilla et al., 2017; Ríos-Málaver et al., 2020), así como índices de Inequidad para determinar qué tan diferentes fueron las estaciones de muestreo (Rös et al., 2012), estos análisis se realizaron mediante el software estadístico R-Project 3.1.3. Para estimar el número de comunidades efectivas, se calcularon los valores de diversidad beta, utilizando los índices de diversidad de Shannon y Simpson (Jost et al., 2011; Jost y Gonzalez-Oreja, 2012; Calderón-Patrón et al., 2012), mediante la fórmula de diversidades alfa y beta propuesta por Jost (2007). Se realizaron curvas de rango de abundancia para determinar gráficamente la estructura de las comunidades a lo largo del gradiente altitudinal entre 2.700 y 3.400 msnm. Adicionalmente, se realizaron análisis de agrupamiento mediante el índice de Bray-Curtis para determinar la similaridad de las estaciones de muestreo mediante la composición y abundancia de las especies con el paquete VEGAN del software R-Project 3.1.3 (Pyrcz et al., 2009; Casas-Pinilla et al., 2017).
Resultados
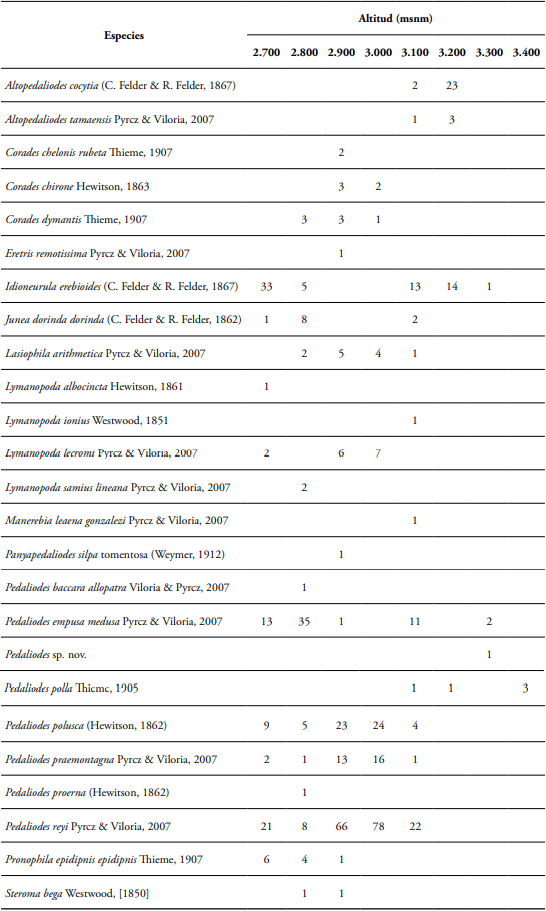
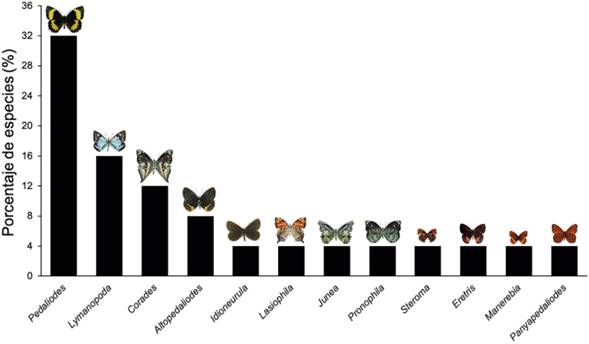
Se recolectaron 539 individuos de mariposas adultas, representados en 12 géneros y 25 especies pertenecientes a la subtribu Pronophilina (Nymphalidae: Satyrinae), donde el género más abundante fue Pedaliodes, con un 69% que equivale a 372 individuos del total de los individuos encontrados (Figura 2). Dentro de este género se determinaron ocho especies, siendo Pedaliodes reyi Pyrcz & Viloria, 2007 con 195 individuos la especie más abundante, seguida de Pedaliodes polusca (Hewitson, 1862) con 65 y Pedaliodes empusa (C. Felder & R. Felder, 1867) con 62 respectivamente (Tabla 1). El siguiente género más abundante fue Idioneurula, con un solo taxón, Idioneurula erebioides (C. Felder & R. Felder, 1867) representando el 12%, equivalente a 66 individuos recolectados, seguido por el género Altopedaliodes con el 5,4%, representado en 29 individuos, donde la especie con mayor abundancia fue Altopedaliodes cocytia (C. Felder & R. Felder, 1867) con 25 individuos. Finalmente, el género Lymanopoda contó con el 3,5% de los individuos registrados, especies como Lymanopoda lecromi Pyrcz & Viloria, 2007 con 15 individuos fue la más abundante del género, seguida de Lymanopoda samius Westwood, 1851 con 2 individuos (Figura 2).
Tabla 1 Listado de las especies de mariposas Pronophilina (Nymphalidae: Satyrinae) registradas en un gradiente altitudinal del Cerro de Tierra Negra, Norte de Santander Colombia.
Fuente: elaboración propia.
Fuente: elaboración propia.
Figura 2 Porcentaje de especies de los géneros pertenecientes a la subtribu Pronophilina del Cerro de Tierra Negra, Norte de Santander, Colombia.
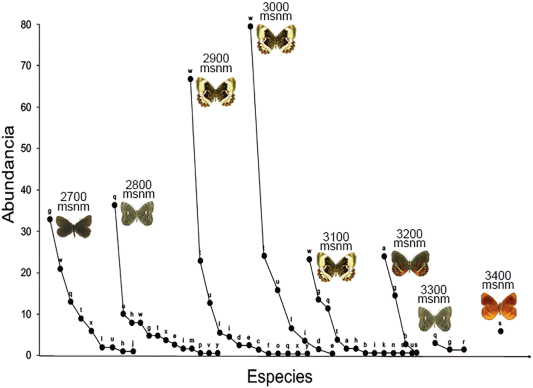
La distribución de las abundancias mostró que la especie con mayor presencia en el rango altitudinal de 2.700-3.100 msnm es P. reyi (letra w). En el rango altitudinal de 3.200 msnm, las especies con mayor presencia fueron A. cocytia e I. erebioides (a y g, respectivamente). A 3.300 msnm, la especie con mayor presencia fue P. empusa (letra q) y a 3.400 msnm, la única especie encontrada fue Pedaliodes polla Thieme, 1905 (letra s) (Figura 3).
Fuente: elaboración propia.
Figura 3 Estructura de los ensamblajes de mariposas de la subtribu Pronophilina en los diferentes niveles de elevación, a través de las curvas de distribución de abundancia del Cerro de Tierra Negra, Norte de Santander, Colombia. Altopedaliodes cocytia (a), Altopedaliodes tamaensis (b), Corades chelonis (c), Corades chirone (d), Corades dymantis (e), Eretris remotissima (f), Idioneurula erebioides (g), Junea dorinda (h), Lasiophila arithmetica (i), Lymanopoda albocincta (j), Lymanopoda ionius (k), Lymanopoda lecromi (l), Lymanopoda samius (m), Manerebia leaena gonzalezi (n), Panyapedaliodes silpa tomentosa (o), Pedaliodes baccara (p), Pedaliodes empusa (q), Pedaliodes sp. nov. (r), Pedaliodes polla (s), Pedaliodes polusca (t), Pedaliodes praemontagna (u), Pedaliodes proerna (v), Pedaliodes reyi (w), Pronophila epidipnis (x), Steroma bega (y).
La completitud del muestreo a nivel general arrojó un valor de 98,52% a lo largo de todo el gradiente altitudinal, recolectando la mayoría de las especies esperadas para la zona (25 especies observadas de 35 especies esperadas) (Figura 4).
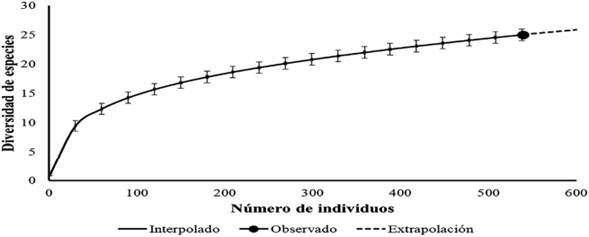
Figura 4 Curva de completitud para el ensamblaje de especies basada en enrarecimiento y extrapolación, según el método de Chao y Jost (2012), Cerro de Tierra Negra, Norte de Santander, Colombia.
La diversidad de orden 0 (Q0), que representa la riqueza potencial de especies, evidenció que las estaciones altitudinales de 2.800 y 2.900 msnm, pertenecientes a bosque altoandino, fueron los sitios con la mayor riqueza de especies efectivas observadas (Q0= 13), seguido de la estación a 3.100 msnm, con 12 especies. Asimismo, se consideró que el resto de las estaciones pertenecientes al bosque altoandino tiene cada una, una diferencia no mayor a dos especies efectivas. Por consiguiente, se encontró que las estaciones restantes tienen, a 2.700 msnm nueve especies, y a 3.100 msnm siete especies efectivas. En las estaciones altitudinales pertenecientes al páramo, la diferencia entre el número de especies no fue superior a tres especies efectivas, donde la estación a 3.200 msnm tiene cuatro especies, a 3.300 msnm tres especies, y a 3.400 msnm sólo una especie efectiva (Figura 5). Para la diversidad de orden 1 (Q1), representada por las especies más frecuentes o comunes, se encontró que 2.800 msnm, perteneciente al bosque altoandino, es la estación con mayor número de especies efectivas, seguido de 3.100 msnm con valores de Q1= 7,3 y 6,21 respectivamente (Figura 5).
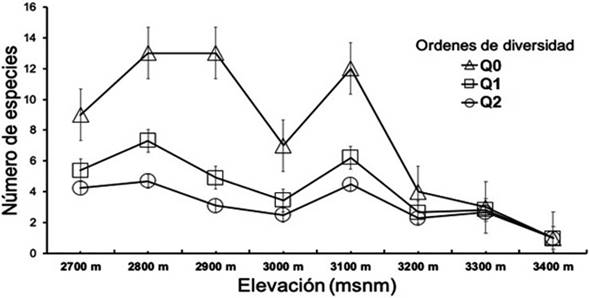
Fuente: elaboración propia.
Q0: riqueza de especies, Q1: especies comunes (exp H’); Q2: especies dominantes (inverso de Simpson 1/D).
Figura 5 Perfiles de diversidad alfa para la comunidad de mariposas registradas en las estaciones altitudinales 2.700-3.400 msnm en el Cerro de Tierra Negra, municipio de Pamplona, Norte de Santander.
Finalmente, con la medida de diversidad de orden 2, basada en la dominancia de especies y que indica el número de especies muy abundantes, se observaron tendencias muy similares a lo observado con la diversidad de orden 1 (Q1), donde la estación de 2.800 msnm que pertenece a bosque altoandino, sigue siendo el valor más alto con 4,69 especies efectivas, seguido de 3.100 msnm con 4,47.
Al comparar los hábitats, las diversidades Q0, Q1 y Q2 registraron valores similares de riqueza y diversidad, donde se pudo observar una disminución de estos a medida que se ascendió en el gradiente altitudinal de 2.700-3.400 msnm.
Los perfiles de diversidad beta entre pares de altitudes, mostraron el recambio de especies dentro del ensamblaje, con el incremento de la altitud, un valor intermedio de 1,36 y un valor alto de diversidad beta de 2,0, que representa el equivalente a dos comunidades efectivas para la diversidad de orden 0 (Q0).
En la diversidad de orden 1 (Q1), la comparación de 2.900 vs 3.000 msnm, mostró un valor más bajo de 1,02 comunidades efectivas (7 especies comunes), entre 3.100 y 3.200 msnm, se registró un recambio intermedio de 1,47 comunidades efectivas (4 especies comunes). El par de elevaciones con el más alto valor de diversidad beta para Q1, fue en 3.300 vs 3.400 msnm, con un valor de 2,00, en el cual no se registraron valores de especies comunes entre estos niveles altitudinales.
El valor de Q2, varió entre los pares de elevaciones a medida que se aumentó el gradiente altitudinal, con valores medios y altos entre 1,31 y 2,00 comunidades efectivas, donde se observó que los niveles altitudinales con menor recambio fueron entre 2.900 y 3.000 msnm, reflejando que, de las especies abundantes, sólo 3 son consideradas como muy abundantes: Pedaliodes polla, P. praemontagna Pyrcz & Viloria, 2007 y P. reyi. (Thieme, 1905). El valor medio/alto de 1,56 comunidades efectivas en el perfil beta, refleja valores altos de recambio de especies en los niveles altitudinales entre 3.100 y 3.200 msnm, donde se registraron especies como I. erebioides y A. cocytia, como las más abundantes entre este par de altitudes. Por último, los niveles altitudinales de 3.300 vs 3.400 msnm mostraron que hay especies no compartidas entre los pares de altitudes, con P. empusa como la más abundante a 3.300 msnm y P. polla como la más abundante a 3.400 msnm. Esto refleja que el recambio de especies mediante los órdenes de diversidad Q0, Q1 y Q2 sucede a medida que se aumenta la elevación en las franjas altitudinales estudiadas (Figura 6).
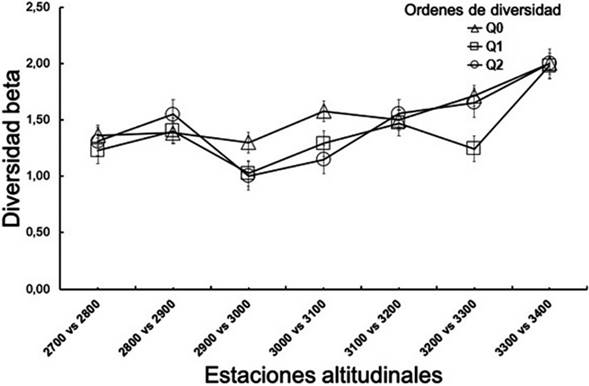
Fuente: elaboración propia.
Figura 6 Perfiles de diversidad beta, para los diferentes niveles altitudinales entre 2.700 y 3.400 msnm en el Cerro de Tierra Negra, Norte de Santander, Colombia. Se evidencia la riqueza de especies (Q0), especies comunes (Q1, exponencial de Shannon) y especies dominantes (Q2, inverso de Simpson).
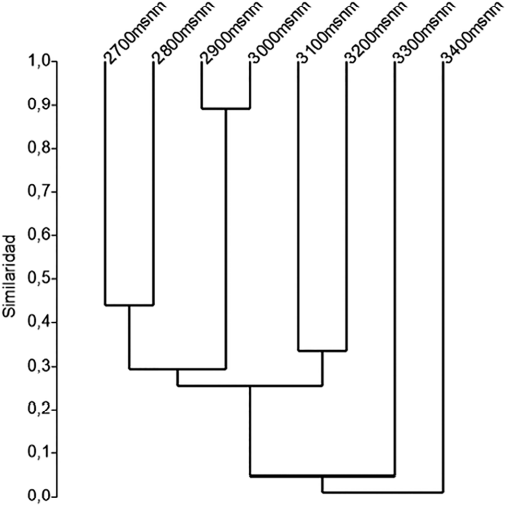
El análisis de similitud de la composición de pronofilinos a lo largo del gradiente altitudinal 2.700-3.400 msnm, mediante el uso del índice de Bray Curtis, mostró el agrupamiento de especies entre estaciones altitudinales pertenecientes al bosque altoandino (franja 2.700-3.000 msnm) y páramo (3.100-3.400 msnm). La franja altitudinal de 2.700-3.000 msnm refleja valores entre el 45% y 90% de similitud en cuanto a composición de especies, con especies propias de este par de altitudes, como Corades chelonis Hewitson, 1863, Corades chirone Hewitson, 1863, Corades dymantis Thieme, 1907, Eretris remotissima Pyrcz & Viloria, 2007, Lymanopoda albocincta Hewitson, 1861, Lymanopoda lecromi Pyrcz & Viloria, 2007, L. samius, Panyapedaliodes silpa tomentosa (Weymer, 1912), Pedaliodes baccara Thieme, 1905, Pedaliodes proerna (Hewitson, 1862), Pronophila epidipnis Thieme, 1907 y Steroma bega Westwood, [1850] [Figuras 8 y 9 (Anexo)]. Asimismo, la franja de páramo presenta especies propias como, Altopedaliodes cocytia, A. tamaensis Viloria & Pyrcz, 2007, Manerebia leaena gonzalezi Pyrcz & Viloria, 2006, Pedaliodes sp. nov. y P. polla, con un procentaje de similitud variante del 10% con respecto a la franja de bosque altoandino. También se encontró una relación entre las dos franjas altitudinales (franja altoandina y páramo), en las cuales se presentaron algunas especies compartidas como I. erebioides, Junea dorinda (C. Felder & R. Felder, 1862), P. empusa, P. polusca, P. praemontagna y P. reyi (Figura 7).
Discusión
Las 25 especies de mariposas de la subtribu Pronophilina observadas en este estudio a lo largo del gradiente altitudinal, representan el 70,12% de las especies de mariposas de este grupo esperadas para la zona, por lo cual se considera que el muestreo es representativo para todo el gradiente de 2.700 a 3.400 msnm, con una completitud de 0,98.
Trabajos como el de Pyrcz y Wojtusiak (1999) reportaron 44 especies de Pronophilina distribuidas en un gradiente altitudinal en la Cordillera Occidental colombiana con numerosos endemismos propios del Chocó Biogeográfico. Del mismo modo para esta región de la Cordillera Occidental, Pyrcz y Viloria (1999) registraron siete nuevos pronofilinos en los orobiomas altoandinos y de páramos en Colombia. Otros estudios realizados en parte de la Cordillera de Mérida, Venezuela, Pyrcz y Wojtusiak (2002) registran 22 especies de pronofilinos, Pyrcz y Viloria (2007) reportaron 56 especies de Pronophilina para la región del Tamá (frontera de Venezuela y Colombia), relacionando niveles de riquezas similares y confirmando el alto grado de endemismo que este grupo de mariposas tiene en la Cordillera Oriental, donde se encuentran especies como L. samius, P. praemontagna, A. tamaensis y A. cocytia que también fueron encontradas en el presente estudio.
En este tipo de ecosistemas en Ecuador, Pyrcz et al. (2009) reportan un total de 48 especies en un gradiente altitudinal desde 1.500 a 2.600 msnm, en el que, las mariposas del género Pedaliodes representaron el 82,11% del total de la muestra, corroborando la dominancia de dicho género en los diferentes niveles de altitud. El estudio de Pyrcz y Garlacz (2012) se reportaron 22 especies de Pronophilina para dos localidades cercanas y ecológicamente similares en la cordillera de Mérida en Venezuela Monte Zerpa, y el Baho. Al igual que en nuestro estudio, encontraron diferencias en la estructura y composición de los ensamblajes de Pronophilina especialmente marcadas por la ausencia de Pedaliodes ornata Grose-Smith & W. F. Kirby, 1895 en el sector del Baho. Estos resultados muestran que en las zonas de alta montaña del Nororiente colombiano la riqueza promedio de especies de Pronophilina puede oscilar entre 22 y 25 especies, conteniendo tanto especies raras y endémicas como especies no descritas para la ciencia. Viloria et al. (2010) encontraron un total de 23 especies de Pronophilina en la Cordillera de la Costa en Venezuela, en las que el género Pedaliodes representa el 34,78% del material estudiado, valores cercanos en el presente trabajo muestran un 32% en términos de riqueza. Mahecha-Jiménez et al. (2011), reportaron 13 especies de alta montaña en un bosque al suroriente de Bogotá, siendo bajo en comparación con las 25 especies reportadas en este trabajo. Marín et al. (2014) contabilizaron 38 especies de esta subtribu en el valle de Aburrá, lo cual es indicativo de la gran riqueza que puede albergar este tipo de ecosistemas de bosques secundarios de la alta montaña. Las 25 especies de Pronophilina encontradas en este estudio, son un aporte importante en relación comparativa al sistema montañoso de la Cordillera de los Andes, ya que en trabajos como el de Henao-B. y Stiles (2018), se hallaron 13 especies de pronofilinas en dos reservas altoandinas de la Cordillera Oriental. Recientemente Clavijo-Giraldo et al. (2020) reportaron 38 especies de Pronophilina en la reserva forestal El Romeral, cerca de Medellín, Antioquia, las cuales en su mayoría pertenecen al género Pedaliodes, (36,84% del total).
Al comparar la abundancia proporcional de pronofilinas mediante los datos publicados por Álvarez-Hincapié et al. (2017) en la Cordillera Central colombiana, se evidencia que cinco especies representan más del 50% de los individuos registrados. Para el nororiente colombiano, los datos publicados por Olarte-Quiñonez et al. (2016) registraron un patrón similar donde tres especies aportan más del 50% de los individuos registrados en las localidades pertenecientes a los complejos de páramos Almorzadero, Santurbán y Tamá, éstas especies fueron P. reyi (21,27%), A. cocytia (20,34%) e I. erebioides (12,95%). En el presente trabajo, cuatro especies contribuyen el 71% de los individuos, P. reyi (36,18%), I. erebioides (12,24%), P. polusca (12,06%) y P. empusa (11,5%), encontrándose similaridad taxonómica entre las especies determinadas en estos dos estudios de pronofilinas de la región nororiental, por cuanto ambas áreas pertenecen biogeográficamente al complejo de páramos de Santurbán, y albergan especies endémicas de la Cordillera Oriental (Olarte-Quiñonez et al., 2016).
La diversidad alfa, evidenció un mayor número de especies en las estaciones altitudinales bajas, fenómeno que indica que la diversidad disminuye a medida que aumenta la altitud, posiblemente en relación a la estructura del paisaje y a la variación proporcional de la temperatura ambiental (Olarte-Quiñonez et al., 2016; Casas-Pinilla et al., 2017).
La diversidad de orden 1 (Q1) presentó valores elevados a altitudes medias, esta observación sustentaría la hipótesis de dominio medio y efecto Rapoport (Pyrcz y Wojtusiak, 2002; Mahecha-Jiménez et al., 2011; Carrero-S. et al., 2013). Con esto se comprueba el cambio de diversidad a medida que aumenta la altitud en el gradiente, debido al recambio de especies en la transición de zonas bajas hacia zonas altas (Pyrcz et al., 2009) y también a la plasticidad ecológica que tienen los insectos para adaptarse a cambios en la composición del hábitat (Pyrcz, 2004; Rös et al., 2012). La diversidad de orden 2, que es el inverso del índice de Simpson, presentó similaridad en el mismo patrón, disminuyendo sus valores en la zona alta del gradiente (3.200- 3.400 msnm), evidenciando mayor diversidad a bajas altitudes.
Olarte-Quiñonez et al. (2016) registraron que los valores de diversidad disminuyen proporcionalmente en comparación con las coberturas vegetales y en las diferentes escalas altitudinales. Los valores de diversidad en los diferentes órdenes (Q0, Q1 y Q2) se reducen en la comparación de coberturas que varían desde arbustos y matorrales (2.607-2.880 msnm), pasando por bosque natural denso (2.620-3.177 msnm), a páramo y subpáramo (2.780-3.741 msnm).
El patrón de diversidad a diferentes escalas a lo largo del gradiente altitudinal ascendente, mantiene en aspectos generales una disminución, lo que se relaciona paralelamente con la disponibilidad de plantas huéspedes, alimenticias y de percha (Pyrcz et al., 2009; Carrero-S. et al., 2013). Las estaciones con alta riqueza de especies (Q0), son zonas con baja equidad, lo que indica que hay una mayor cantidad de especies raras o poco comunes que presentaron cambios en la composición (Jost et al., 2010; Rös et al., 2012).
La diversidad beta muestra un patrón de recambio de especies, diferenciando dos comunidades dentro del gradiente altitudinal. Tal diferencia se refleja en la comparación del intervalo 3.000-3.100 msnm (Figura 6), en la cual se observa un valor de 1,58 comunidades efectivas y se reduce la cantidad de especies compartidas, siendo cuatro especies (Lasiophila arithmetica Pyrcz & Viloria, 2007, Pedaliodes polusca, P. praemontagna y P. reyi) las que se comparten del total de 15 registradas entre el par de elevaciones. Estas diferencias muy posiblemente se deben al cambio de vegetación (cobertura vegetal) entre una franja de bosque altoandino a 3.000 msnm y el páramo a 3.100 msnm (Pyrcz y Garlacz, 2012), esto puede deberse a la transformación del hábitat y su uso en actividades de pastoreo, siembra de cultivos y minería que se adelantan en la zona de estudio, estos factores posiblemente tienden a simplificar la estructura del ecosistema y por consiguiente a aumentar el número de especies compartidas, principalmente en hábitats abiertos (Tobar-L., 2004; Rös et al., 2012; Olarte-Quiñonez et al., 2016; Acevedo et al., 2018).
La similitud entre las estaciones de muestreo mediante el índice de Bray-Curtis, evidenció dos grupos, el primer grupo en las estaciones entre 2.700 y 3.000 msnm, y el segundo en aquellas entre 3.100 y 3.400 msnm, grupos que coinciden con bosques altoandinos y páramo, respectivamente. La determinación de estos agrupamientos a lo largo del gradiente está regulada principalmente por la gran representatividad de la subtribu Pronophilina que domina estas franjas altitudinales (Pyrcz y Viloria, 2007; Pyrcz et al., 2009; Pyrcz y Garlacz 2012) y también por la heterogeneidad del paisaje y las variaciones microclimáticas que se reflejan en el cambio en las coberturas vegetales mencionadas por Olarte-Quiñonez et al. (2016).
El presente estudio es un importante complemento para el conocimiento de los ensamblajes de mariposas diurnas en ecosistemas altoandinos y páramo en el departamento Norte de Santander, Colombia, en el cual se registran especies adicionales a las ya publicadas por Olarte-Quiñonez et al., (2016), como P. epidipnis, C. dymatis, C. chirone, L. albocincta, Lymanopoda ionius (Westwood, 1851), P. silpa tomentosa y P. proerna.
También se evidenció un cambio en la composición de mariposas en relación al incremento de altitud, reflejando la transición entre el ecosistema de bosque altoandino hacia páramo. Es importante impulsar la conservación de los ecosistemas de alta montaña por su notable diversidad y alto endemismo de especies de mariposas, además de la importancia ecosistémica en los procesos ecológicos que dan origen al recurso hídrico del departamento de Norte de Santander. Del mismo modo, conocer los patrones de diversidad de las mariposas de alta montaña, nos permite concebir un panorama sobre la composición actual de sus comunidades como posible herramienta para evaluar las amenazas ante un escenario de cambio climático y sus consecuencias sobre las especies potencialmente vulnerables a la extinción.



