











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
El orden Diptera comprende más de 153.000 especies distribuidas en aproximadamente 180 familias y más de 4.000 géneros (Brown et al., 2009). Cerca de 21.000 especies se encuentran en Norteamérica (Courtney y Cranston, 2015) y se estima que hay alrededor de otras 31.000 en el Neotrópico, agrupadas en 118 familias (Amorim, 2009). Dentro del orden, la familia Chironomidae reúne 11 subfamilias (Aphroteniinae, Buchonomyiinae, Chironominae, Chilenomyiinae, Diamesinae, Orthocladiinae, Podonominae, Prodiamesinae, Tanypodinae, Telmatogetoninae y Usambaromyiinae), que comprenden al menos 10.000 especies descritas en más de 400 géneros, de las cuales cerca de 900 están distribuidas en el Neotrópico (Cranston, 1995; Saether, 2000), aunque este es sólo un estimativo, ya que, por ejemplo, para el caso particular de Colombia aún no existe un registro consolidado y actualizado de los quironómidos.
Cabe destacar que la fauna de quironómidos sigue siendo poco conocida a nivel mundial, e incluso en la mayoría de los trabajos limnológicos, las larvas de la familia Chironomidae se identifican a nivel de familia o subfamilia (Prat y Rieradevall, 2011). En algunos casos se pierde gran parte de la información ecológica que se podría obtener de este grupo (Ferrington, 2008; Serra et al., 2017), lo cual se debe probablemente al complejo trabajo que conlleva realizar las preparaciones microscópicas y a la dificultad de determinar taxonómicamente los individuos hasta el nivel de género o especie (Villamarín, 2012).
En Colombia, en particular, los estudios sobre los Chironomidae se han realizado principalmente en la región andina, los cuales consisten en el desarrollo de claves taxonómicas ilustradas para la Sabana de Bogotá (Ospina et al., 1999) y el departamento de Caldas (Hoyos y Días, 2020). En el centro-occidente del país, se destacan 15 nuevos registros de géneros de la familia Chironomidae para la cuenca del río Alvarado, de los cuales ocho pertenecen a la subfamilia Chironominae, cuatro a Orthocladiinae y tres a Tanypodinae (Rojas-Sandino et al., 2018). Por otro lado, Oviedo-Machado y Reinoso-Flórez (2018) estudiaron la composición de la comunidad de larvas de Chironomidae en la cuenca del río Opia (Tolima) y su relación con algunas variables ecológicas. Montaño-Campaz et al. (2019) evaluaron diferentes tipos de variaciones en la morfología de las piezas bucales y en la reducción de las alas de Chironomus columbiensis, expuesto a diversas acciones antrópicas (la minería de oro, la ganadería y los cultivos de café), como una herramienta para estimar los efectos de contaminación en las fuentes hídricas. En la región Caribe, se destaca el trabajo realizado por Barragán et al. (2017), quienes registraron el género Podonumus en el río Guatapurí (Sierra Nevada de Santa Marta), convirtiéndose en el primer registro a nivel de género para la región. Por lo tanto, es fundamental realizar trabajos taxonómicos que permitan evaluar la diversidad de los quironómidos a nivel de género y especie en zonas poco estudiadas, como es el caso de la Sierra Nevada de Santa Marta.
Recientemente, Tamaris-Turizo et al. (2023) realizaron una revisión de los patrones altitudinales registrados para diferentes órdenes y familias de macroinvertebrados acuáticos en Colombia. En dicha revisión, se encuentra que dípteros y la familia Chironomidae siguen un patrón de disminución lineal de la riqueza desde las zonas bajas a las altas que está asociado a características bióticas y abióticas de cada piso altitudinal. Así pues, se espera que la mayor diversidad se encuentre en la parte alta del río y que disminuya en la medida en que se descienda en el gradiente altitudinal.
Métodos
El estudio se realizó en la cuenca del río Gaira, ubicada en la vertiente occidental de la Sierra Nevada de Santa Marta, en el departamento de Magdalena, Colombia. La longitud de la cuenca es de 32,5 km desde su nacimiento hasta la desembocadura del río en el mar Caribe. Esta es una cuenca rectangular que posee zonas muy conservadas en los tramos alto y medio, mientras que en el tramo bajo se desarrollan importantes actividades agropecuarias y de recreación que afectan negativamente la calidad del agua (Tamaris-Turizo et al., 2013). La selección de los sitios de muestreos se realizó considerando el fácil acceso y la seguridad en cada zona (Figura 1).
Sitios de estudio
La Cascada: se localiza a los 1.700 m de altitud, entre los 11°07’39’’ N y los 74°03’14’’ W. El sitio presenta una formación vegetal de bosque muy húmedo subtropical (Bmh-ST), de acuerdo con la clasificación propuesta por Espinal y Montenegro (1963). Dicha localidad es un área que presenta buen estado de conservación y cerca al sitio de muestreo no se realizan actividades agropecuarias extensivas ni otras acciones de origen antrópico que generen impactos negativos sobre los ecosistemas (Figura 1).
La Victoria: se localiza entre los 11°07’44.2” N y los 74°05’35.8” W, a 900 m de altitud. El sector presenta una formación vegetal de bosque húmedo subtropical (Bh-ST), según las categorías mencionadas. En esta zona se desarrollan actividades como la caficultura y en algunos sectores la producción ganadera a baja escala (Figura 1).
Puerto Mosquito: se ubica a 60 m de altitud, entre los 11°10’26’’ N y los 74°10’37’’ W. De acuerdo con la clasificación de las zonas de vida de Holdridge adaptada para Colombia por Espinal y Montenegro (1963), el sector presenta un ecosistema de monte espinoso tropical (Me-T). Aguas arriba del sitio de muestreo se encuentra una captación de agua y río abajo hay varias zonas utilizadas por los turistas y residentes como balnearios. Esto, junto a los impactos de la ganadería y la agricultura, convierten a este sector en el de mayor intervención antrópica entre los sitios seleccionados (Figura 1).

Muestreos
Se realizó la recolección de las larvas de quironómidos en los tres sitios escogidos del río Gaira, así: parte alta (La Cascada: LA), sector medio (La Victoria: LV) y zona baja (Puerto Mosquito: PM). Estos lugares se muestrearon en ocho ocasiones, en los meses de abril, marzo y mayo del 2018 y febrero, mayo, junio y julio del 2019. Esta ventana temporal cubrió los periodos de lluvias y sequía característicos de la región Caribe colombiana. En cada sitio de muestreo se estableció un tramo recto del cauce de 50 m aproximadamente, de acuerdo con las consideraciones espaciales de Frissell et al. (1986), con el criterio de obtener una suficiente heterogeneidad geomorfológica de los hábitats acuáticos. Se tomaron muestras en sustratos como hojarasca, roca, grava y sedimento. Tras seguir parcialmente el método propuesto por Zúñiga y Cardona (2009), los organismos se colectaron con una red Surber (250 μm de diámetro de malla y 0,09 m2 de área) en los sustratos de hojarasca, grava (2-24 cm) y sedimentos (0,2-2 cm), mientras que en el sustrato roca (>25 cm) se realizó una colecta manual con pinceles y pinzas durante 30 minutos. Se colectaron cuatro réplicas por cada sustrato y posteriormente se integraron en una sola muestra para caracterizar la diversidad de cada una de las estaciones. El material recolectado en los muestreos se almacenó en bolsas de polietileno y se conservó en una solución de etanol al 70%.
En cada sitio y durante cada muestreo se midieron in situ la temperatura del agua (°C), el pH (unidades), la conductividad eléctrica (μS/cm) y el oxígeno disuelto (mg/L), empleando un equipo multiparamétrico (OHAUS ST-300).
Análisis en laboratorio
Montaje e identificación de los organismos
Las muestras colectadas en los sustratos de sedimento y grava se tamizaron (250 μm de poro) y se lavaron suavemente con abundante agua, para evitar dañar los organismos. Cada fracción recogida del tamiz se depositó en bandejas, donde se separaron los quironómidos con pinzas o pinceles y se preservaron con etanol al 70% en frascos debidamente etiquetados.
Los especímenes se separaron en subfamilias y morfogéneros teniendo en cuenta caracteres macromorfológicos, como la forma de la cabeza, el tamaño de los seudópodos y la longitud total. Posteriormente, se procedió a preparar los montajes en láminas portaobjetos con el fin de identificarlos (Andersen et al., 2013). Los organismos completos se colocaron individualmente en tubos Eppendorf de 2 mL, con una solución de hidróxido de potasio al 10% y se sometieron al baño de María a 85°C durante 20 minutos, con el fin de degradar la materia orgánica y transparentar las estructuras morfológicas que sirven para la determinación taxonómica. Una vez puestos en el baño maría, los organismos se lavaron durante dos minutos en soluciones de etanol al 70% y 90%. Finalmente, a cada organismo se le diseccionó la cabeza y el cuerpo para fijarlos individualmente en Euparal®. La cabeza se ubicó en posición ventral en una gota de Euparal® y posteriormente se dispuso un cubreobjeto (de 0,5 x 0,5 mm) y se presionó levemente hasta que las mandíbulas quedaran abiertas. A un lado de este montaje, se empleó otra gota de Euparal® y se colocó el cuerpo en posición lateral. Finalmente, los montajes se dejaron durante 24 horas a temperatura ambiente. Cada lámina se etiquetó con la información de colecta.
Se tomaron fotografías a las estructuras morfológicas de los individuos, empleando una cámara digital Leica a ICC50 HD, acoplada a un microscopio óptico (Leica, DM500, EUA) y a partir de estas se realizaron descripciones microscópicas. Finalmente, las fotografías de cada una de las larvas se tomaron con un microscopio-estereoscopio (Leica M205A) acoplado con cámara integrada Leica DFC450. Las imágenes de los micropreparados se utilizaron para identificar a los organismos hasta el nivel taxonómico de género mediante el empleo de claves y descripciones (Andersen et al., 2013; Epler, 2001; Heiri, 2013; Laurindo da Silva et al., 2018; Moreno et al., 2000; Ospina et al., 1999; Paggi, 2001; Prat et al., 2014; Trivinho-Strixino, 2011).
Análisis de los datos
La diversidad se estimó mediante los números efectivos de géneros basados en la serie de números de Hill ( 0 D, 1 D y 2 D); estas medidas permiten integrar datos de abundancia y cumplen con una serie de atributos matemáticos de acuerdo con la interpretación intuitiva del concepto de diversidad del orden q ( q D) (Jost, 2006). De esta manera, 0 D indica la riqueza de géneros; 1 D es el número efectivo de taxones igualmente comunes, el cual se calculó a partir del exponente de entropía de Shannon-Weanner (eH) y 2 D es el número de efectivo de géneros dominantes, que se computó a partir del inverso del índice de entropía de Simpson (1/λ). Estas estimaciones se realizaron con el programa iNEXT Online software, que permite la interpolación y extrapolación de la diversidad de especies (Hsieh et al., 2016) con un nivel de confianza del 95%. Los análisis se realizaron en el programa R, versión 1.3.1 y el paquete Vegan (R Core Team, 2021).
Resultados
Caracterización ambiental
La menor temperatura se observó en el sitio de muestreo La Cascada (17,8°C) y la mayor se registró en Puerto Mosquito (24,2°C). El pH en las estaciones La Cascada y La Victoria presentaron valores levemente por encima de la neutralidad (7,14 y 7,07 respectivamente), mientras que en Puerto Mosquito el pH fue ligeramente ácido (6,09). Por otra parte, el mayor registro de oxígeno disuelto se observó en La Victoria (6,24 mg/L) y el menor (4,82 mg/L) en Puerto Mosquito (Tabla 1).
Tabla 1 Promedio y desviación estándar de las variables físicas y químicas tomadas in situ en cada sitio de muestreo.
| Variable | La Cascada 1.700 msnm | La Victoria 900 msnm | Puerto Mosquito 60 msnm |
| Temperatura (°C) | 17,8 ± 1,4 | 18 ± 1,0 | 24,2 ± 0,7 |
| pH (Unidades) | 7,14 ± 0,2 | 7,07 ± 0,3 | 6,09 ± 0,6 |
| Oxígeno disuelto (mg/L) | 5,38 ± 1,7 | 6,24 ± 1,3 | 4,82 ± 1,8 |
Fuente: Elaboración propia.
Diversidad de Chironomidae
En total se recolectaron 1.319 larvas agrupadas en 3 subfamilias y 14 géneros. Todos los géneros son primeros registros para la cuenca del río Gaira y para la Sierra Nevada de Santa Marta: Ablabesmyia, Chironomus, Corynoneura, Cricotopus, Cryptochironomus, Djalmabatista, Endotribelos, cf. Manoa, Onconeura, Parametriocnemus, Polypedilum, Riethia, cf. Saetheria y Stenochironomus. La subfamilia Orthocladiinae presentó la mayor abundancia con 776 individuos (59% de la abundancia total), seguida de Chironominae con 499 individuos (38%) y Tanypodinae con 44 individuos (3%).
La tabla 2 resume la abundancia y diversidad de quironómidos encontradas en cada sitio. En La Victoria se hallaron 12 géneros, cinco igualmente abundantes ( 1 D) y uno dominante ( 2 D, Cricotopus), siendo este el sitio con la mayor abundancia (708) y riqueza. En Puerto Mosquito se registraron 390 individuos de 11 géneros diferentes, de los cuales cerca del 40% de los especímenes fueron del género Polypedilum. Por su parte, La Cascada presentó las menores abundancia (221 individuos) y riqueza ( 0 D=8), con Parametrionecmus como el taxón más abundante.
Tabla 2 Resumen de la abundancia y diversidad de larvas de la familia Chironomidae encontrada en cada uno de los sitios. La abundancia (N) se estimó como el número total de larvas encontradas en las muestras y la diversidad se definió por medio de los primeros tres números de Hill: 0D=riqueza genérica, 1 D=géneros igualmente comunes y 2D=géneros dominantes.
| Sitio | N | 0 D | 1 D | 2 D |
| Puerto Mosquito | 390 | 11 | 6,06 | 1,29 |
| La Victoria | 708 | 12 | 5,79 | 1,32 |
| La Cascada | 221 | 8 | 5,03 | 1,31 |
Fuente: Elaboración propia.
El muestreo realizado fue representativo de la diversidad que puede encontrarse en los tres sitios de estudio a lo largo del río Gaira, ya que se observó una cobertura de taxones mayor al 90% (Figura 2A) y una variación baja de la cobertura del muestreo respecto a la riqueza de géneros (Figura 2B). Adicionalmente, al corregir la riqueza de géneros por el número de larvas encontradas, se ve que las comparaciones entre los sitios son consistentes: la mayor diversidad de larvas se observó en Puerto Mosquito y La Victoria, y la menor en La Cascada (Figura 2C).

Fuente: elaboración propia.
Figura 2 Perfiles de la diversidad observada y estimada, y estimación de la cobertura de los muestreos en función de la abundancia y la diversidad. A. Curva de completitud de muestras. B. Cobertura de los muestreos respecto a la diversidad de géneros. C. Curvas de rarefacción y extrapolación de muestreos basadas en el tamaño de la muestra (diversidad).
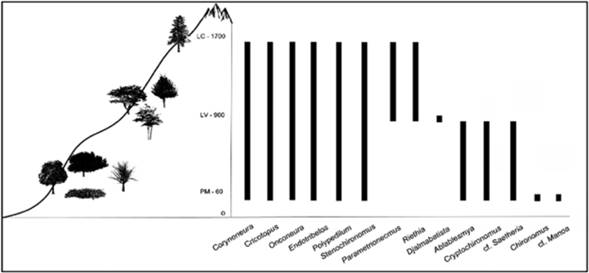
Los resultados sugieren que los géneros Corynoneura, Cricotopus, Endotribelos, Onconeura, Polypedilum y Stenochironomus tienen una amplia distribución altitudinal en la cuenca del río Gaira (desde los 60 hasta los 1700 m), mientras que las larvas de Parametriocnemus y Riethia parecen estar restringidas a la media montaña, los 900 y los 1700 m. Por otro lado, los géneros Ablabesmyia, Cryptochironomus y cf. Saetheria podrían estar limitados a la parte baja de la montaña (entre los 60 y los 900 m). Cabe destacar que Chironomus y cf. Manoa sólo se encontraron en la parte baja del río - Puerto Mosquito (Figura 3).
Discusión
En general, los 14 géneros encontrados en este estudio demuestran que la cuenca del río Gaira alberga una importante riqueza de géneros de Chironomidae. La composición observada en la familia Chironomidae coincide con lo reportado por Rojas-Sandino et al. (2018) y Oviedo-Machado y Reinoso-Flórez (2018) en las cuencas de los ríos Alvarado, Anchique y Opia en el departamento del Tolima (Colombia). Así mismo, Medina y Paggi (2004) demostraron que Chironominae fue la subfamilia con mayor riqueza genérica y Orthocladiinae la de menor abundancia en el río Grande, Argentina. De acuerdo con Ashe et al. (1987), la subfamilia Chironominae en el río Gaira presentó similitudes con la descrita para climas tropicales y subtropicales, y corresponde a taxones euritérmicos cálidos. Los resultados de este trabajo también muestran que a lo largo del río Gaira se distribuyen géneros que presentan amplia distribución altitudinal y otros que exhiben una distribución restringida, como se observó en el género Chironomus, el cual es típico para zonas bajas, de acuerdo con lo registrado previamente por Armitage et al. (1983). La variabilidad de estos géneros en el gradiente altitudinal se da en términos de riqueza y composición, pero no de abundancia. No obstante, Acosta y Prat (2010) encontraron que la mayor diversidad la presentó la subfamilia Orthocladiinae en la cuenca del río Cañete (Perú), en un rango altitudinal de 2552 a 4425 m, donde Cricotopus fue el taxón dominante.
Los taxones más abundantes en el río Gaira fueron Cricotopus, Onconeura, Parametriocnemus y Polypedilum. Estos registros coincidieron parcialmente con lo observado en los ríos Carcarañá y Chocancharava y en ríos de la región de las Yungas en Argentina (Príncipe et al., 2007, 2008; Tejerina y Molineri, 2007; Peroet al. 2023) y con los datos de Acosta y Prat (2010) en los Andes peruanos. Al parecer, los taxones mencionados tienen una alta preferencia por hábitats con sustratos rocosos y corrientes de flujo rápido. Estos géneros, además, tienen una distribución amplía a lo largo de la cuenca del río Gaira, lo que concuerda con lo registrado tanto en ecosistemas de zonas bajas como en arroyos de montaña con alta velocidad de corriente (Epler, 2001; Henriques-Oliveira et al., 2003; Trivinho-Strixino, 2011).
Uno de los factores más influyentes en los ecosistemas de agua dulce es la altitud, hecho ya señalado en otros estudios, que indican que hay cambios en las comunidades bentónicas a lo largo del gradiente altitudinal (Jacobsen, 2003, 2008; Ramos-Pastrana et al., 2021; Vannote et al., 1980). La importancia de la altitud se debe a la variabilidad que ocasiona en algunos factores como la velocidad de la corriente, el oxígeno disuelto y la temperatura del agua a lo largo de este gradiente. A su vez, dichas variables controlan en gran medida la distribución, la estructura y la composición de las comunidades bentónicas (González-Trujillo et al., 2020). En este estudio en particular, se observó que la abundancia relativa de las subfamilias responde al gradiente altitudinal, con las subfamilias euritérmicas como Chironominae como las predominantes en zonas bajas, mientras que otros taxones reófilos estuvieron restringidos a zonas altas.
Las especies restringidas a las partes medias y altas de la cuenca parecen tener una estrecha relación con aguas más oxigenadas y, por tanto, con zonas de rápidos. Djalmabatista, por ejemplo, se registró solo en la estación La Victoria; es un género común en corrientes rápidas, en lagos o charcas estacionales y se adhieren a una amplia variedad de sustratos, como hojarasca, gravas y sedimentos (Epler, 2001). Por otra parte, las larvas de Parametriocnemus y Riethia se asociaron a la parte más alta del río (La Cascada), donde las condiciones como la vegetación ribereña y la madera en descomposición son apropiadas para el desarrollo de sus poblaciones (Laurindo da Silva et al., 2018; Trivinho-Strixino, 2011).
Temperaturas más altas en las tierras bajas podrían contribuir a explicar las diferencias entre los puntos de muestreo en términos de la composición de géneros, y en particular, el incremento de la riqueza genérica. Sin embargo, como lo muestran nuestros resultados, el patrón altitudinal no es lineal, y se evidencia una reducción considerable de la riqueza genérica en la estación de menor altitud. Este resultado da indicios de que otros factores más allá de la temperatura podrían afectar los patrones altitudinales. Por ejemplo, los resultados obtenidos pueden estar influidos también por el grado de impacto antropogénico, toda vez que cerca del sitio de muestreo, se localiza un balneario, se localiza una planta captación de agua del río y se realizan actividades agropecuarias de baja intensidad. La presencia del género Chironomus, por ejemplo, suele ser común en ríos contaminados, ya que estas larvas están asociadas a flujos de corriente lenta, con sedimentos ricos en materia orgánica y ambientes pobres en oxígeno disuelto; suelen encontrarse en todo tipo de aguas, tanto en sistemas lénticos como en lóticos (Trivinho-Strixino, 2011). El presente estudio demuestra la necesidad de profundizar en la identificación de las especies de quironómidos, pues éstas se distribuyen lo largo del gradiente altitudinal de acuerdo con requerimientos ambientales particulares, como lo demostraron Puntí et al. (2009). Es importante, pues, un estudio integrativo que sume herramientas taxonómicas y genéticas en la identificación de las especies propias de distintas altitudes en la Sierra Nevada de Santa Marta. Para este fin, la asociación entre larvas, pupas y adultos, junto a los análisis de códigos de barras de ADN, podrían contribuir de manera significativa a avanzar en la caracterización de la diversidad de este grupo de insectos (Prat et al., 2018).
Conclusiones
Los resultados de esta investigación demuestran que hay una alta riqueza de géneros de Chironomidae en la cuenca del río Gaira, la cual está relacionada con los cambios ambientales asociados a un gradiente altitudinal. Esto pone en evidencia que la Sierra Nevada de Santa Marta es un importante reservorio de biodiversidad, donde los factores y procesos que subyacen a esta alta biodiversidad deben ser explorados en futuros estudios.

