











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroducción
El cultivo de cítricos es uno de los sectores agroindustriales más importantes en el mundo, siendo una rama decisiva de la economía de varios países e importante generador de empleos (Mattos, 2019; Silva et al., 2022). En Colombia, este cultivo tiene un gran potencial de crecimiento, pasó de exportar 2,6 millones de dólares en 2012 a más de 58 millones de dólares en 2021, exportaciones lideradas por los departamentos de Caldas, Antioquia, Santander, Valle del Cauca y Quindío (Procolombia, 2022).
Este cultivo se ve afectado durante su desarrollo por artrópodos plaga, entre los que se destacan: el ácaro tostador Phyllocoptruta oleivora (Acarina: Eriophyidae), el ácaro blanco Poliphagotarsonemus latus (Acarina: Tarsonemidae), la escama articulada Selenaspidus articulatus (Hemiptera: Diaspididae), el piojo blanco Unaspis citri (Hemiptera: Diaspididae), Orthezia praelonga (Hemiptera: Ortheziidae), la cochinilla harinosa Planococcus citri (Hemiptera: Pseudococcidae), las moscas blancas Aleurocanthus woglumi, Aleurothrixus floccosus y Dialeurodes citri (Hemiptera: Aleyrodidae), el minador Phyllocnistis citrella (Lepidoptera: Gracillariidae), Diaphorina citri (Hemiptera: Liviidae) y los picudos de los cítricos Compsus obliquatus Hustache, 1938 y Compsus viridivittatus (Guérin-Méneville, 1855) (Coleoptera: Curculionidae: Entiminae) (Espinal et al., 2005; León, 2005; O’Brien y Peña, 2012; León y Kondo, 2017; Carabalí y Cardona, 2019).
Los picudos de los cítricos se consideran plagas de importancia económica para la citricultura en varios países productores. En Colombia, C. viridivittatus se encuentra distribuida en los departamentos de Antioquia, Caldas, Cundinamarca, Quindío, Risaralda, Santander, Tolima y Valle del Cauca (O’Brien y Peña, 2012). Su alta capacidad de reproducción y el daño que las larvas y adultos causan a las raíces, hojas, flores y frutos, lo convierten en una seria amenaza para la citricultura colombiana (Cano et al., 2002; Peñaloza y Díaz, 2004). Estos insectos presentan una generación por año, o sea que son univoltinos. Los huevos se colocan en las hojas y tienen una duración de 10 a 20 días; las larvas pueden vivir entre 7 y 18 meses. Cuando nacen las larvas, rápidamente se dirigen o se dejan caer hacia el suelo para alimentarse de las raíces de los árboles; consumen el tejido externo de la corteza y de esta forma ocasionan el daño. Las altas infestaciones de larvas pueden producir la muerte de los árboles jóvenes cuando logran consumir totalmente la corteza de sus raíces (León y Kondo, 2017), ocasionando pérdidas hasta del 50% (Gallego et al., 2004).
En Colombia se registra a Gonalezia gloriosa (Hymenoptera: Encyrtidae) y a Fidiobia sp. (Hymenoptera: Platygastridae) parasitando el 50% de los huevos de Compsus (Jacas et al., 2007; Estrada y Soto, 2011). Trigonospila unicaldasi (Díptera: Tachinidae) se encontró en Caldas (Colombia) y descrita como una nueva especie de parasitoide de adultos de C. viridivittatus, ejerciendo control del 75% de este insecto plaga en campo (Soto, 2011; Vinasco et al., 2017).
Para su control, los agricultores realizan aplicaciones de productos químicos altamente tóxicos, ocasionando desequilibrio ambiental y afectando los enemigos naturales (Cano, 2002). Por lo tanto, se hace necesario utilizar productos alternativos a los agroquímicos convencionales. El polisulfuro de calcio es obtenido por el tratamiento térmico del azufre y la cal (Abbot, 1945). Esta mezcla se conoce, principalmente, debido a su acción fungicida (Montag et al., 2005), y también es utilizada como acaricida e insecticida (Venzon et al., 2007; Soto, 2009; Soto et al., 2011, Fajardo et al., 2013; Cabrera et al., 2018). El efecto tóxico del polisulfuro de calcio a los insectos se da por la reacción de los compuestos de este sobre la planta, con el agua y el gas carbónico, resultando en gas sulfhídrico y azufre coloidal (Abbot, 1945).
Los compuestos orgánicos volátiles inducidos por herbívoros y liberados por las plantas son importantes como mecanismo de defensa para repeler insectos, para promover la reproducción al atraer polinizadores, son mensajeros intraespecíficos e interespecíficos, actúan como disuasivos de alimentación y de oviposición en insectos plaga (Marín y Céspedes, 2007; Ignacimuthu et al., 2011; Ángeles et al., 2012; Scala et al., 2013; Dong et al., 2016; Hijaz et al., 2016), además sirven como señales importantes para parasitoides y depredadores para ubicar a su hospedante o presa (Baldwin et al., 2001; Ponzio et al., 2013; Schettino, 2017; Salamanca et al., 2018; De Lange et al., 2019). Así se abren opciones para desarrollar nuevas estrategias en programas de control biológico y etológico, con el objetivo de mejorar la eficacia de los enemigos naturales en la supresión de poblaciones de plagas en los cultivos (Peñaflor y Bento, 2013). Además, la defensa de las plantas contra herbívoros implica la participación de una amplia gama de metabolitos, algunos actúan directamente como armas defensivas contra enemigos como son los glucosinolatos y benzoxazinoides, que desencadenan la acumulación de polisacáridos protectores como una barrera contra áfidos y patógenos (Maag et al., 2015).
En este trabajo se estudió el efecto del polisulfuro de calcio en la respuesta olfativa de Trigonospila unicaldasi a olores de plantas de cítricos no atacadas y atacadas por Compsus viridivittatus.
Materiales y métodos
El estudio se realizó en el Centro de Investigación y Cría de Enemigos Naturales de la Universidad de Caldas, ubicado en el municipio de Manizales, Colombia (Coordenadas: 5° 05’N y 75° 40’ W). Se evaluó el efecto del polisulfuro de calcio sobre la capacidad de búsqueda del parasitoide T. unicaldasi por volátiles provenientes de plantas de cítricos infestadas por C. viridivittatus.
Se estimaron las concentraciones letales y subletales del polisulfuro de calcio sobre adultos de C. viridivittatus, los cuales se colectaron en cultivos de cítricos de la granja Montelindo de la Universidad de Caldas. Las concentraciones evaluadas se seleccionaron a través de bioensayos preliminares, se situaron entre el límite inferior, donde el producto no causó mortalidad, y el límite superior de respuesta, con un 100% de mortalidad.
El efecto letal y subletal del polisulfuro de calcio sobre C. viridivittatus, se ha estudiado de acuerdo con metodología adaptada de Penteado (2000), manejando tratamiento térmico del azufre y cal virgen, utilizando para cada litro de agua, 250 g de azufre y 125 g de cal virgen. La concentración obtenida del polisulfuro de calcio fue de 32°Baumé.
Para los bioensayos de concentración-respuesta se seleccionaron ramas de cítricos las cuales se infestaron con adultos de C. viridivittatus. Las ramas se depositaron en recipientes trasparentes de 25 cm de largo por 20 cm de diámetro y dentro de este se colocó una base de espuma humedecida, después se expusieron los individuos a la aplicación directa con el producto a diferentes concentraciones (0,25; 0,50; 0,75; 1,0; 1,25; 1,50; 1,75; 2,0; 2,25; 2,5; 2,75 y 3,0%), se utilizó un testigo absoluto y por cada tratamiento se realizaron cinco repeticiones. La mortalidad se evaluó cada 12 h después a la aplicación del producto y la concentración-mortalidad se estimaron por el análisis de Probit (Finney, 1971), y la concentración letal media (CL50) y concentración letal 95 (CL95) se calcularon utilizando la función descrita por Pacheco y De Freitas-Rebelo (2013) en el programa estadístico R (R Development Core Team).
La respuesta olfativa de adultos de T. unicaldasi a los volátiles de la planta hospedera infestada con C. viridivittatus y asperjados con polisulfuro de calcio fue evaluada en olfatómetro tipo “Y” (diámetro interno de 0,5 cm y el largo de los brazos de 5 cm) (Pallini et al., 1997). El aparato está constituido por un tubo de vidrio en forma de “Y”, siendo que cada brazo está conectado por una manguera a dos cajas de plástico transparentes (50 x 36 x 43 cm). Dentro de estas cajas se pusieron las fuentes de olores, que fueron plantas de cítricos no atacadas por fitófagos o plantas de cítricos atacadas con C. viridivittatus. El flujo de aire se produjo por un ventilador conectado a la base del tubo de vidrio, formando una corriente de aire uniforme en los dos brazos del aparato. La velocidad de la corriente de aire dentro del olfatómetro fue de 0,45 m/s en cada brazo, medida por fluxómetros digitales y calibrada por registro manual. Los adultos de T. unicaldasi se colocaron en la extremidad basal del olfatómetro, contra la corriente de aire formada en el interior del aparato. Se consideraron en el análisis aquellos insectos que respondieron y llegaron al punto de unión del tubo en donde realizaron escogencia por una de las fuentes, considerándose una respuesta positiva la llegada del parasitoide hasta la extremidad de uno de los brazos del olfatómetro.
Los tratamientos correspondieron a la exposición de los parasitoides T. unicaldasi a agua destilada (control) y a C. viridivittatus con residuos de polisulfuro de calcio (1,75%), correspondiente a la CL50 de la plaga (Tabla 1). Como fuentes de olores se utilizaron plantas de cítricos con 20 hojas no infestadas y plantas infestadas con 20 adultos de C. viridivittatus por planta. Los adultos de T. unicaldasi se liberaron individualmente en el olfatómetro y evaluaron uno a uno, en un total de 20 parasitoides por repetición, totalizando tres repeticiones por tratamiento. A cada cinco adultos evaluados, las posiciones de las fuentes de olores se invirtieron, y al final de cada repetición las hojas de cítricos se trocaron con el fin de evitar la pseudorepetición (Ramírez et al., 2000). Las diferencias entre los números de parasitoides que escogieron para cada una de las fuentes de olores fueron evaluadas utilizando la prueba estadística Replicated Goodness of Fit (Sokal y Rohlf, 1995) considerando una frecuencia esperada de 0,5 para cada fuente de olor (Pallini et al., 1997).
Resultados y discusión
En la toxicidad letal del polisulfuro de calcio se obtuvieron la Cl 50 de 1,75 y la Cl 95 de 2,26% de concentración del producto a 32° B (Tabla 1).
Tabla 1 Toxicidad del polisulfuro de calcio a adultos de C. viridivittatus.
| Producto | N1 | Coeficiente Angular ± EP2 | Cl50 3 (I.C. a 95%) | Cl95 4 (I.C. a 95%) | X2 | P |
|---|---|---|---|---|---|---|
| Polisulfuro de calcio | 160 | 0,39± 0,03 | 1,75 (1,42 -1,96) | 2,26 (2,11- 2,65) | 0,83 | 0,96 |
1 Número de individuos evaluados
2 Coeficiente angular y error padrón de la media
3 Concentración letal media e intervalo de confianza a 95%
4 Concentración letal que causa 95% de mortalidad e intervalo de confianza a 95%.
En los análisis de olfatómetro, el parasitoide T. unicaldasi pulverizado con agua prefirió los olores de plantas de cítricos infestadas con C. viridivittatus a olores de plantas no infestadas (Gp= 42,35; g.l.= 1; p< 0,0001) (Figura 1). De estos parasitoides, 86% prefirieron plantas de cítricos infestadas con C. viridivittatus a plantas no infestadas. Estos resultados indican qué volátiles de la planta hospedera atacada por C. viridivittatus tienen un papel relevante en la localización de la presa por parte de T. unicaldasi.
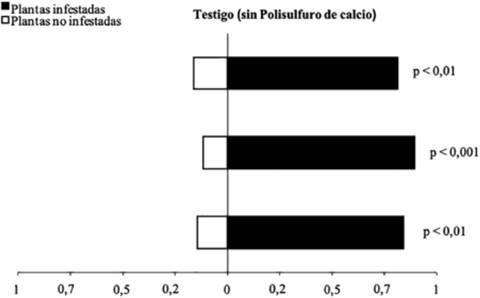
Figura 1 Respuesta del parasitoide T. unicaldasi a los olores de plantas no infestadas y plantas de cítricos infestadas por C. viridivittatus en olfatómetro tipo “Y”. Cada barra corresponde a una repetición (n= 20 parasitoides). (Gp = 42,35; g.l.= 1; p< 0,0001).
Cuando C. viridivittatus se asperjó con la dosis subletal de polisulfuro de calcio, el parasitoide T. unicaldasi expuesto a residuos del producto continuó prefiriendo los olores de plantas de cítricos infestadas con C. viridivittatus a los de plantas no infestadas (Gp= 52,21; g.l.= 1; p< 0,0001) (Figura 2). De estos parasitoides, 81% prefirieron plantas de cítricos infestadas con C. viridivittatus asperjadas con Polisulfuro de calcio, lo que indica que la dosis subletal aplicada de este producto no afecta la capacidad de búsqueda de T. unicaldasi.
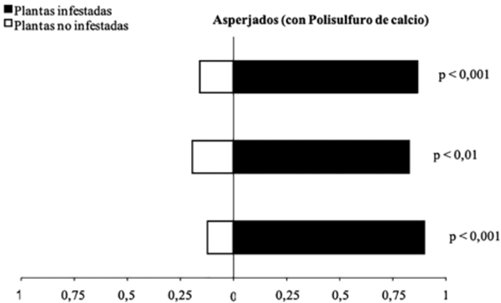
Figura 2 Respuesta del parasitoide T. unicaldasi a los olores de plantas no infestadas y plantas de cítricos infestadas por C. viridivittatus con aplicación de polisulfuro de calcio en olfatómetro tipo “Y”. Cada barra corresponde a una repetición (n=20 parasitoides). (Gp = 52,21; g.l.= 1; p< 0,0001).
De acuerdo con los resultados obtenidos, T. unicaldasi puede diferenciar entre olores de plantas de cítricos no atacadas y plantas de cítricos atacadas por C. viridivittatus. El polisulfuro de calcio en la concentración de 1,75% no interfirió en la respuesta olfativa de T. unicaldasi alimentado de presas asperjadas, pudiendo discriminar entre plantas infestadas por C. viridivittatus y plantas no infestadas.
T. unicaldasi expuesta a concentraciones subletales de polisulfuro de calcio en el campo, al conseguir encontrar sus presas, no compromete el control biológico natural ejercido sobre C. viridivittatus. De esta manera T. unicaldasi puede aumentar la eficiencia en la localización de presas, pudiendo reducir el tiempo necesario para localizarlas. Con estos resultados se evidencia que el polisulfuro de calcio aplicado al 1,75% de concentración se podría utilizar en plantaciones de cítricos cuando C. viridivittatus alcance los niveles de daño económico, complementando el control con liberaciones del parasitoide T. unicaldasi.
Los volátiles de plantas juegan un papel vital en la atracción de los insectos herbívoros (Guerin y Visser, 1980), además sirven como señales para atracción de parasitoides y depredadores (Turlings y Wackers, 2004). Ton et al. (2007), observaron en maíz el aumento de compuestos volátiles ocasionado por el daño de Spodoptera exigua (Lepidoptera: Noctuidae) a la planta, lo que generó atracción del parasitoide Cotesia marginiventris (Hymenoptera: Braconidae), por lo que deducen que los volátiles juegan un papel clave en las decisiones de localización del hospedante. Norin (2001) y Metcalf y Luckmann (1992) expresaron que los áfidos y otros insectos son repelidos por compuestos volátiles terpenoides, como el salicilato de metilo, inhibiendo su atracción hacia la planta hospedante.
Murillo et al. (2020) encontraron que cuando Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) fue asperjado con la dosis subletal de azadirachtina, el parasitoide Encarsia formosa (Hymenoptera: Aphelinidae) expuesto a residuos de azadirachtina continuó prefiriendo los olores de plantas de fríjol infestadas con T. vaporariorum a los de plantas no infestadas.
Venzon et al. (1999) observó en experimentos de invernadero que el predador Orius laevigatus (Hemiptera: Anthocoridae) prefirió plantas de pepino infestadas por el ácaro Tetranychus urticae (Acari: Tetranychidae) que plantas no atacadas por estos herbívoros. Teodoro (2003) estudió la interferencia de los acaricidas fenibutatina y azufre sobre la capacidad de búsqueda del ácaro predador Iphiseiodes zuluagai (Acari: Phytoseiidae) a olores de plantas no atacadas y atacadas por los ácaros fitófagos Oligonychus ilicis y Brevipalpus phoenicis (Acari: Tetranychidae, Tenuipalpidae), encontrando que los acaricidas en concentraciones subletales interfirieron en la capacidad de búsqueda del ácaro predador.
Con el entendimiento del papel que juegan los semioquímicos en la ecología y en los mecanismos de emisión de los compuestos orgánicos volátiles de las plantas inducidos por herbívoros, y que estos compuestos pueden incrementar atracción de enemigos naturales; se presentan opciones para desarrollar nuevas estrategias en programas de control biológico y etológico a partir de estos compuestos, con el objetivo de mejorar la eficacia de los enemigos naturales en la supresión de poblaciones de plagas en los cultivos (Reddy y Guerrero, 2004; Peñaflor y Bento, 2013).

