Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
Los bosques andinos se consideran como uno de los principales centros de expresión de diversidad en el mundo debido a la alta presencia de endemismos y procesos de especiación originados por la confluencia de especies de origen austral-antártico y holártico (Churchill et al., 1995; van der Hammen, 2000). La dinámica tectónica, geomorfológica y climática de la región andina, que en su conjunto influyeron en la diversidad genética, ha generado condiciones particulares en la vegetación, a tal punto de ser considerada un “paraíso biogeográfico” (Sugden, 1996; van der Hammen y Cleef, 1983). Un factor relevante en este ecosistema es la presencia de niebla o el fenómeno conocido como “precipitación horizontal” cuya causa principal es la condensación de masas de aire húmedo desplazadas en el proceso de ascenso adiabático (Stadtmüller, 1987); esta característica propia de los bosques andinos les confiere una función fundamental en la regulación del ciclo hidrológico (Cavelier, 1997; Rodríguez et al., 2006).
Los bosques andinos en Colombia han sido fuertemente fragmentados. La principal causa en dicha dinámica es el cambio de uso del suelo de vocación forestal a actividades agropecuarias como la ganadería y el establecimiento de cultivos (Armenteras et al., 2003). Reportes de Cavelier y Etter (1995) indican que dichas coberturas se encuentran en un proceso de transformación superior al 70% de su estado inicial.
En esta situación, se hace necesario evaluar el estado de la diversidad, las funciones y beneficios que provee dicho ecosistema con el fin de tomar acciones en su planificación y manejo (Noss, 1990). Uno de los principales servicios ecosistémicos que prestan los bosques es el de regulación climática por medio del secuestro de carbono (de Groot et al., 2010). En consecuencia, la declaratoria de diferentes figuras de áreas protegidas ha contribuido en su permanencia y provisión (Llambí et al., 2020).
En este contexto, el siguiente estudio tiene como objetivo realizar un análisis ecológico, a nivel de comunidad, de la vegetación arbórea de bosques andinos ubicados en el Parque Natural Chicaque. Para lograr dicho fin, se caracterizó la composición y estructura de la vegetación. Adicionalmente, se interpretó la diversidad. Posteriormente, se determinaron las existencias aproximadas de la biomasa aérea.
Metodología
Área de estudio
El parque natural Chicaque se localiza en Colombia, región andina, cordillera oriental - vertiente occidental, departamento de Cundinamarca, municipio de San Antonio del Tequendama, vereda Chicaque (Figura 1). El predio del parque abarca 312 ha, de las cuales 243 se encuentran, registradas ante el sistema nacional de áreas protegidas, como reserva natural de la sociedad civil (Barbosa y Saénz, 2017). La vegetación del área es conocida como bosque de niebla, el cual es su principal objeto de conservación.
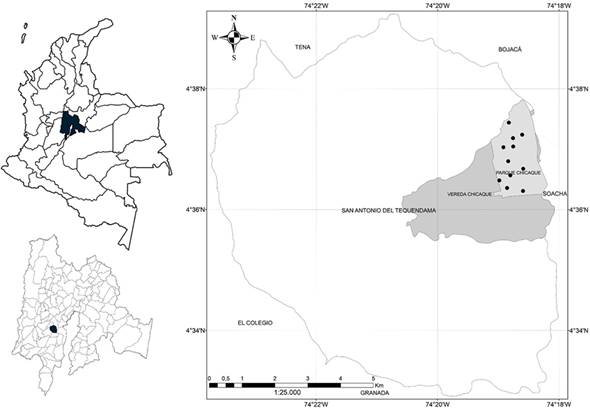
Las condiciones climáticas medias corresponden a temperaturas de 14,6 a 15,3°C; precipitación bimodal (máximos de marzo a mayo y de octubre a noviembre), promedio anual de 2000 mm; y humedad relativa de 75 a 86% (Rivera y Córdoba, 1998). El rango altitudinal de la reserva va de los 2000 a los 2700 msnm, reuniendo vegetación de selva subandina a andina (Cuatrecasas, 1958), clasificándose en la zona de vida bosque muy húmedo montano bajo -bmh-MB- (Holdridge, 1967). En el parque natural Chicaque se encuentra la formación geológica Guadalupe, compuesta de areniscas compactas y friables con intercalaciones de limolitas y arcillolitas/lutitas, proveniente del cretáceo superior y de origen marino (van der Hammen, 1995). Los suelos son de origen coluvio-aluvial y de textura franco arcillosa, en la cota superior del parque, y arcillosos, en el límite inferior (Peréz-Preciado, 2000).
Muestreo
Se realizó un muestreo simple al azar de 11 parcelas de 10 x 100 m o 0,1 ha, recolectando la información de todos los individuos que presentaran DAP ≥ 10 cm (Rangel y Velázquez, 1997). En este, se registraron las variables generales de un inventario forestal (Carrera, 1996): nombre común; nombre científico; circunferencia a la altura del pecho -CAP- (cm), calculado a DAP; altura total (m); altura fustal (m); y diámetro de la copa (m). Cabe aclarar que los datos de altura se recolectaron a partir de estimación visual, por la misma persona en todas las parcelas. Paralelamente, por cada taxón distinguido, se colectó un espécimen botánico de un individuo que presentara la mayor cantidad de rasgos morfológicos y reproductivos, de ser posible, para el proceso de determinación taxonómica (Liesner, 1997).
Análisis y procesamiento de los datos de vegetación
Composición y estructura
Se identificó y se hizó ingreso de los espécimenes botánicos en el Herbario Forestal “Gilberto Emilio Mahecha Vega” -UDBC- de la Universidad Distrital (Carvajal-Rojas, 2002). Además, se confirmaron y actualizaron las determinaciones empleando las plataformas Tropicos (Missouri Botanical Garden, 2018) y JStor Global Plants (Ryan, 2018). Así mismo, se evaluaron las singularidades bióticas por especie (Bernal et al., 2023; MADS, 2017).
Posteriormente, se halló el índice de valor de importancia (Finol, 1971) y el índice de predominio fisionómico (Matteucci y Colma, 1982). En suma, se integró el concepto de gremios ecológicos a las especies más importantes y dominantes de la vegetación muestreada (Budowski, 1965). Las condiciones ecológicas del bosque se evaluaron en función de las categorías propuestas por Guariguata y Ostertag (2001).
Adicionalmente, se incluyó un diagrama de cajas y bigotes (Williamson et al., 1989) de las alturas totales de cada parcela, analizándolas desde los estratos propuestos por Holdridge (1967), adaptados por Rangel y Lozano (1986): Rasante (ra), < 0,3 m; Herbáceo (h), 0,31-1,5 m; Arbustivo (ar), 1,51-5 m; Arbolitos (Ar), 5-12 m; Arbóreo inferior (Ai) 12-25 m; Arbóreo superior (As), > 25 m. Complementariamente, se agregó el cálculo de los coeficientes de asimetría -Ap- (Pearson, 1895) y curtosis -Cp- (Pearson, 1902) para cada unidad de muestra. La interpretación de los coeficientes se dió de la siguiente forma: Ap > 0, asimetría positiva o a la derecha; Ap < 0, asimetría negativa o a la izquierda; Ap ≈ 0, simetría / Cp > 3, leptocurtosis; Cp < 3, platicurtosis; Cp ≈ 3, mesocurtosis.
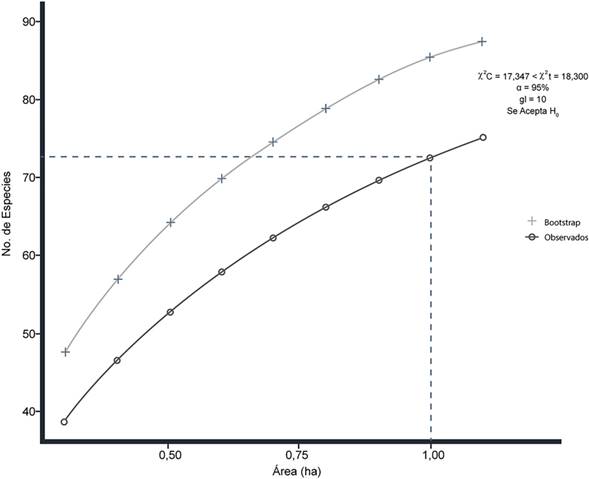
Los datos de los inventarios se procesaron en el software R (R Development Core Team, 2017). La diversidad se calculó utilizando el paquete vegan (Oksanen et al. 2018); se incluyeron índices de diversidad alfa y riqueza (Simpson, 1949; Shannon y Weaver, 1963; Hulbert, 1971). También, se generó una curva de acumulación de especies en función del área muestreada (Smith y van Belle, 1984), agregando un estimador de representatividad estadística con el método de remuestreo Bootstrap (Efron, 1979); y se verificó el nivel de significancia con la prueba de X2 (Pearson, 1992), donde la hipótesis nula (H0) se asocia a la ausencia de diferencias significativas entre los valores calculados (riqueza acumulada) y los valores estimados (bootstrap), y la hipótesis alterna (H1) a su presencia.
Biomasa aérea
Se estimó la cantidad de biomasa aérea, en función de las características meteorológicas de la zona de vida del área de estudio, utilizando el modelo tipo I - 1.3. - LMm de Álvarez et al. (2012) (Ecuación 1), en árboles, y el modelo Cagtrfn de Beets et al. (2012) (Ecuación 2), en helechos arbóreos. Para los cálculos de los modelos mencionados, se utilizaron los datos de altura total (m) y DAP (cm) del inventario; adicionalmente, se consultó la densidad básica de la madera (g cm-3), por especie, en los reportes de Zanne et al. (2009) y Kindt et al. (2017), en el caso de no encontrar el dato específico, se empleó el promedio de la categoría taxonómica disponible (media del género o de la familia), acorde al principio de conservadurismo filogenético (Eloisa-León et al., 2010). Consecuente a la determinación de los valores de biomasa aérea, se procedió a hallar las cantidades estimadas de carbono -C- (Ecuación 3) y dióxido de carbono equivalente -CO2e- (Ecuación 4) (Phillips et al., 2011).
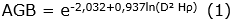
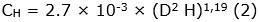
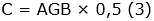
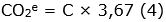
Siglas: AGB, Biomasa Aérea (kg); C, Carbono (kg); CH, Carbono de helechos arbóreos (kg); D, DAP (cm); H, Altura Total (m); ρ, Densidad Básica de la Madera (g cm-3).
Finalmente, se definió una distribución de la biomasa aérea por categorías diamétricas, cada 10 cm (Bradford y Murphy, 2019). Las gráficas se desarrollaron empleando las librerías ggplot2 (Wickham et al., 2020) y ggpmisc (Aphalo y Slowikowski, 2020); en suma, se incluyó un análisis de varianza (Marden, 2003). Con fines comparativos, se calculó una inferencia a hectárea, definiendo el error de muestreo mediante una prueba t a un alfa del 95% (Student, 1908). Adicionalmente, se incluyó un análisis en las especies dominantes que acumularan más del 50% de la biomasa aérea, agregando líneas de tendencia con el método de regresión ponderada localmente -LOESS- (Cleveland y Devlin, 1988) para identificar patrones de concentración de biomasa según los DAP.
Resultados
Composición
Se encontró un total de 75 especies arbóreas, distribuidas en 58 géneros y 36 familias; de estas, cuatro morfotipos se determinaron a nivel de género, 1 a familia y 3 están indeterminados (Anexo 1, Tabla 5). Las familias con mayor riqueza se muestran en la Tabla 1. Las especies que presentan algún grado de amenaza son: roble (Quercus humboldtii Bonpl., Fagaceae), en categoría vulnerable -VU-; y cedro (Cedrela montana Moritz ex Turcz., Meliaceae), en categoría casi amenazada -NT- (UICN, 2012). Así mismo, la palma boba (Cyathea indet.) se reporta en el Apéndice II de CITES (CITES, 2017).
Estructura
El dosel medio se encuentra entre los 12 y los 15 m, acorde a la mediana de las alturas totales (Figura 2). A nivel general, se muestra un estado homógeneo en la estructura vertical del bosque; donde los cuartiles -Q- 1 y 2, en su mayoría, se asocian al estrato de arbolitos (5-12 m) y los cuartiles 3 y 4 al arboréo inferior (12-25 m). Se hace excepción de las parcelas 11, donde una proporción considerable de indivudos se concentra en Q1 y Q2 (2-7 m), y 2, que por el contrario de la 11, reune buena parte de la densidad en Q3 y Q4 (17-22 m). Por otra parte, a pesar de ser pocos, se evidencia la presencia de individuos en el estrato arbóreo superior (>25 m).
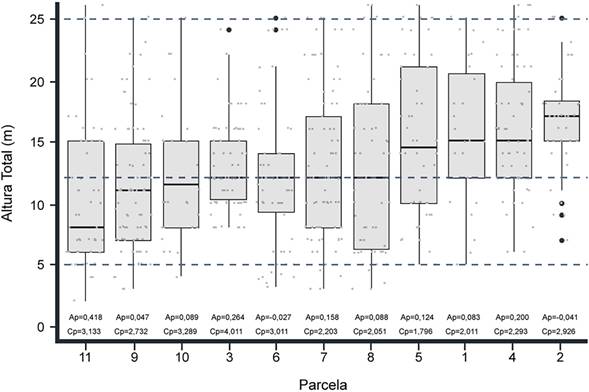
Fuente: Elaboración propia.
Figura 2 Diagrama de cajas y bigotes de alturas totales por parcela en la vegetación arbórea del bosque andino del parque natural Chicaque. Las líneas punteadas representan los límites de las categorías propuestas por Holdridge (1967), adaptadas por Rangel-Ch. y Lozano-C. (1986).
En cuanto a las medidas de forma, las parcelas 3, 10 y 11, que son leptocurticas, tienen mayor concentración de árboles en el dosel medio (12-15 m); las demás, al ser platicurticas o mesocurticas (6), presentan dispersión de los individuos en los estratos dominantes (arbolitos y arbóreo inferior). Esta misma tendencia es evidente en las unidades de muestra 1, 2, 6, 8 y 9, que se caracterizan por ser simétricas. El resto de las parcelas (4, 5 y 7) tienen una proporción considerable en los estratos arbóreo inferior y superior, lo que explica su asimetría positiva.
Por otra parte, el 50% del índice de valor de importancia se concentró en 13 especies, el 70% en 24. Así mismo, el 50% del índice de predominio fisiónomico se acumuló en 11 especies, el 70% en 19. Dos especies se caracterizan por su alta abundancia: Cyathea indet. y Montanoa quadrangularis Sch. Bip.; estas se distinguen, de las presentes en la Tabla 2, por no coincidir con las de mayor aporte de biomasa aérea (Figura 5).
Tabla 2 IVI, IPF y gremios ecológicos de especies importantes y dominantes (50% del IPF) en la vegetación arbórea del bosque andino del parque natural Chicaque.
| Especie | IVI (%) | IPF (%) | Gremio |
|---|---|---|---|
| Cecropia angustifolia | 5,98 | 6,79 | HD |
| Miconia symplocoidea | 5,03 | 6,25 | HD |
| Picrasma excelsa | 5,26 | 5,81 | HD |
| Cyathea indet. | 4,49 | 5,19 | HD |
| Styloceras laurifolium | 4,16 | 4,53 | HD |
| Montanoa quadrangularis | 3,90 | 4,84 | HD |
| Croton cf. gossypiifolius | 3,66 | 4,41 | HD |
| Aegiphila bogotensis | 2,83 | 3,97 | HD |
| Miconia floribunda | 3,19 | 3,60 | HD |
| Ficus pertusa | 3,41 | 3,53 | HD |
| Quercus humboldtii | 2,90 | 3,52 | EP |
Fuente: Elaboración propia.
Diversidad
Diversidad alfa
Los índices calculados evidencian diferentes niveles de heterogeneidad y diversidad: alta para las parcelas 4, 7 y 8 (rangos de D = 0,91-0,93, H’ = 2,67-2,88); media en 5 y 10 (D = 0,89, H’ = 2,39-2,63); baja en 1, 3, 6 y 9 (D = 0,80-0,87, H’ = 2,06-2,32); y muy baja en 2 y 11 (D = 0,64-0,71, H’ = 1,60-1,63) (Magurran, 2004). La considerable abundancia de Cyathea indet. y de Miconia symplocoidea Triana en las unidades muestreales 11 y 2, respectivamente, explica los resultados obtenidos. En cuanto a la riqueza: las parcelas 4 y 8 poseen el valor más alto; las parcelas 3, 5, 6 y 9 se encuentran cercanos a la media del muestreo; y la parcela 2 presenta la menor cantidad de especies.
La media general del muestreo se encuentra en 17 especies arbóreas y 58 individuos en 0,1 hectáreas (Tabla 3). El mayor coeficiente de variación se encontró en la densidad, donde es evidente la incidencia de la amplitud del rango, por los valores de las parcelas 1, 10 y 2; esta última, tambien influye en la dispersión de la riqueza.
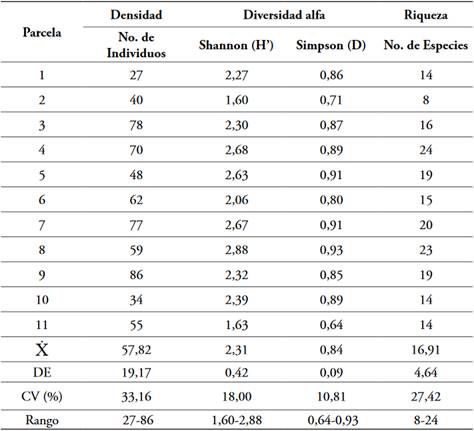
Representatividad del muestreo
La curva de acumulación de especies reune un total de 75 especies en 11 parcelas de 0,1 ha; se estima un aproximado de 72 especies arbóreas por hectárea (Figura 3). Tras el resultado de la prueba de x2 se acepta la hipótesis nula, dado que no se encontraron diferencias significativas con los valores estimados en el remuestreo Bootstrap. Por tanto, se afirma que el muestreo es representativo a nivel florístico.
Biomasa aérea, carbono y CO2 equivalente
Se estimó una cantidad media de 20,62 ± 4,94 Mg 0,1ha-1 de biomasa aérea, 10,31 ± 2,47 Mg 0,1ha-1 de carbono y 37,84 ± 9,06 Mg 0,1ha-1 de CO2e; a un error de muestreo de 23,94% (Tabla 4). Se infiere un valor de 206,22 ± 49,37 Mg ha-1. Las parcelas 1 y 8 presentaron los menores valores, debido a la baja densidad, en el primer caso, y a la ausencia de individuos en categorías diamétricas superiores a los 30 cm, en la segunda unidad de muestreo en mención.
Tabla 4 Biomasa aérea, carbono y CO2 equivalente en la vegetación arbórea del bosque de niebla del parque natural Chicaque. Siglas: EM, error de muestreo; LC, límite de confianza.
| Parcela | AGB ( Mg 0,1ha-1) | Parcela ( Mg 0,1ha-1) | AGB ( Mg 0,1ha-1) |
|---|---|---|---|
| 1 | 10,809 | 5,405 | 19,835 |
| 2 | 22,034 | 11,017 | 40,433 |
| 3 | 26,320 | 13,160 | 48,297 |
| 4 | 31,268 | 15,634 | 57,378 |
| 5 | 28,590 | 14,295 | 52,462 |
| 6 | 22,105 | 11,053 | 40,563 |
| 7 | 26,235 | 13,118 | 48,141 |
| 8 | 8,180 | 4,090 | 15,010 |
| 9 | 20,720 | 10,360 | 38,021 |
| 10 | 15,803 | 7,901 | 28,998 |
| 11 | 14,781 | 7,390 | 27,123 |
| Ẋ | 20,622 | 10,311 | 37,842 |
| DE | 7,439 | 3,720 | 13,651 |
| CV (%) | 36,075 | ||
| EM (%) | 23,940 | ||
| LC | 4,937 | 2,469 | 9,059 |
Fuente: Elaboración propia.
Por otro lado, las categorías diamétricas no presentaron diferencias significativas entre ellas, según el análisis de varianza (p-valor = 0,181); sin embargo, se evidencia una tendencia de mayor aporte medio de biomasa aérea, y menor desviación estándar, en 10-20 cm y > 70 cm de DAP (Figura 4). Este comportamiento difiere de lo esperado en bosques con crecimiento antiguo, donde la biomasa aérea debería concentrarse en el intervalo de 30 a 70 cm de DAP (Bradford y Murphy, 2019). Complementariamente, los cálculos de asimetría y curtosis indican que: la categoría 20-30 cm es simétrica, 10-20 presenta asimetría a la izquierda, las demás se caracterizan por asimetría a la derecha; por otra parte, 40-50 es leptocúrtica, las otras son platicúrticas. Cabe mencionar la reducción en la cantidad de parcelas que presentan individuos en las categorías diamétricas superiores a los 30 cm.
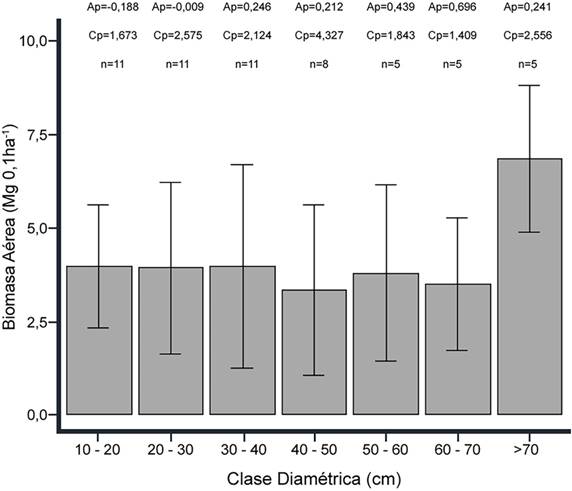
Fuente: Elaboración propia.
Figura 4 Biomasa aérea por distribución diamétrica de la vegetación arbórea del bosque andino del parque natural Chicaque. Los intervalos representan la desviación estándar de las categorías.
Las especies que secuestran la mayor cantidad de carbono en su biomasa aérea son Cecropia angustifolia Trécul, Q. humboldtii y M. symplocoidea. El 50% de esta variable se acumuló en nueve especies; de estas, la presencia de individuos dispersos o únicos en las categorías diamétricas superiores a los 50 cm se da en Q. humboldtii, Ficus pertusa Aubl., Ficus sp. y C. angustifolia (Figura 5). En contraste, Q. humboldtii no presenta individuos en las categorías 10-20 cm y 30-40 cm; así mismo, F. pertusa es ausente en 20-30 cm y de F. maxima solo se encuentran dos árboles en clases diamétricas extremas.
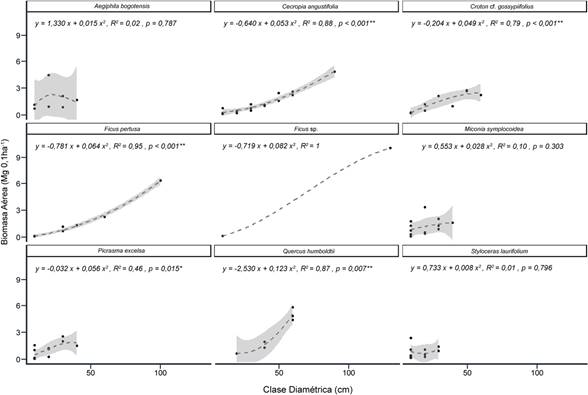
Fuente: Elaboración propia.
Figura 5 Biomasa aérea por distribución diamétrica en especies dominantes (50% de la AGB) de la vegetación arbórea del bosque andino del parque natural Chicaque. Siglas: *, α = 95%; **, α = 99%.
Las regresiones ponderadas localmente de biomasa aérea aplicadas fueron estadísticamente significativas en C. angustifolia, Croton gossypiifolius Vahl, F. pertusa, Q. humboldtii y Picrasma excelsa (Sw.) Planch. Por otra parte, las especies difieren en los alcances de la categoría diamétrica en la que pueden encontrarse individuos: Aegiphila bogotensis (Spreng.) Moldenke, M. symplocoidea, P. excelsa y C. gossypiifolius llegan hasta los 50 o 60 cm; Q. humboldtii, C. angustifolia, F. pertusa y F. sp. pueden presentar diamétros de 70 cm o mayores; y Styloceras laurifolium (Willd.) Kunth se límita en 40 cm. De estos grupos, se evidencian tendencias similares: en el caso del primero, existe relación directamente proporcional entre la biomasa aérea y el diamétro; para el segundo, con un comportamiento similar al descrito anteriormente, que difiere por la reducción en los individuos con mayor diámetro; y S. laurifolium, con patrón continuo, que sugiere estabilidad de la biomasa en las categorías intermedias.
Discusión
Composición
En los bosques altoandinos de la reserva de Carpanta, Gentry (1993) realizó muestreos de 0,1 ha y divulgó datos promedio de 30 especies para árboles con DAP > 10 cm; así mismo, los reportes exponen una cantidad promedio de 91 árboles con DAP > 10 cm. Los niveles de riqueza y de densidad encontrados en este estudio, en promedio (17 especies y 58 individuos en 0,1 ha), son inferiores a los encontrados por el autor anteriormente mencionado. En cuanto a los registros, la vegetación evidencia relación taxonómica con lo reportado por Franco et al. (1986), Rangel-Ch. y Aguirre (1986), Cantillo et al. (2004), Rangel-Ch. et al. (2008), Cantillo y Gracia (2013), Avella y Ávila (2017), Arellano y Rangel-Ch. (2008) y Rojas (2017).
Los resultados encontrados coinciden con lo encontrado por Mendoza-Cifuentes (2012) para la familia Melastomataceae, promedio de 7 especies en transectos de 80 x 5 m en gradientes de 2000 a 2500 msnm; sin embargo, son inferiores respecto a Rubiaceae, cuya media varía de 10 a 12 especies. Por otra parte, en Araliaceae y Clusiaceae la cantidad de géneros fue cercana a los tres taxones característicos, para cada familia, registrados en el norte de los Andes (Gentry, 1982). Así mismo, el número de especies se aproxima a los reportes de Galindo et al. (2003), en Asteraceae (4) y Lauraceae (6). Por último, este estudio concuerda con Hernández et al. (2011), en cuanto a riqueza en Meliaceae (5 especies, 3 géneros); pero, difiere a lo cuantificado por Reina et al. (2010) en Euphorbiaceae (6 géneros).
Estructura
Estructuralmente, la vegetación analizada en este estudio contrasta con los resultados de Cantillo et al. (2004) y Cantillo y Gracia (2013), dado que los pesos ecológicos reportados por estos autores se concentran en una alta proporción (intervalos de 10 a 30% en el IVI y de 15 a 45% en el IPF) en tres especies (Hedyosmum bonplandianum Kunth, Beilschmiedia pendula (Sw.) Hemsl.y Clusia ducu Benth.; Weinmannia rollotii Killip, Weinmannia tomentosa L.f. y Clusia multiflora Kunth, respectivamente), acumulando entre el 30 y 60% de la importancia y dominancia del bosque. Así mismo, Rangel-Ch. (2015) menciona que, en las áreas andinas y altoandinas de la vertiente occidental de la cordillera oriental, se identifican los bosques dominados por Clusia elliptica Kunth e Ilex pernervata Cuatrec. A pesar de coincidir con taxones en la composición, la dominancia difiere notablemente con lo encontrado en esta investigación.
En cuanto a las condiciones de dosel, los estratos dominantes (arbolitos y arboréo inferior) coinciden con lo propuesto por Rangel-Ch. (1991) en transectos de la región andina. Sin embargo, contrasta con otras investigaciones por la baja cantidad de árboles cuya altura supera los 25 m (Cantillo et al., 2004). Aunque, esta misma tendencia se ha evidenciado en otros levantamientos cercanos al área de estudio (Cortés-Ballén et al., 2020).
Diversidad
En cuanto a la diversidad, Restrepo (2016), en la vegetación de Guasca, reporta valores de H’=2,87 y D=0,89 en bosques de crecimiento antiguo, y para bosques con desarrollo tardío los datos corresponden a H’=2,27 y D=0,83; los resultados de este estudio muestran un promedio de 2,31 en Shannon y 0,84 en Simpson, son datos cercanos a la tendencia de bosques con desarrollo tardío. Cantillo et al. (2004), en la reserva Cárpatos, presenta resultados de H’ = 2,87-3,02 y D = 0,91 a 0,93, situación similar a la presentada en la parcela 8; sin embargo, Cantillo y Gracia (2013) registran promedios de H’=3,61 y D=0,97 en los bosques de los cerros orientales de Bogotá, valores contrastantes con los índices calculados en esta investigación, los cuales son inferiores y dan noción de un estado de diversidad media.
Biomasa aérea
De las especies con mayor aporte, en aquellas con relación directamente proporcional entre el diámetro y la biomasa aérea, es evidente la ausencia de individuos en varias categorías diamétricas, indicador de alta heterogeneidad estructural en el bosque debido a patrones de desagregación por árboles con altos valores de área basal (Lutz et al., 2013). Sin embargo, esta condición aplica para los taxones que pueden alcanzar los 70 cm o más, cuya biomasa se encuentra en función de la dominancia (C. angustifolia, F. pertusa, F. sp., Q. humboldtii); dado que los que llegan a los 50 o 60 cm tienden a manifestar reducción al llegar a la última categoría, lo que puede ser indicio de biomasa condicionada por la abundancia (C. gossypiifolius, P. excelsa, M. symplocoidea y A. bogotensis) (Bohn et al., 2014). Así mismo, es posible que S. laurifolium se encuentre en condiciones similares al último grupo descrito, pero los árboles muestreados no manifiestan tendencia al no superar la tercera clase diamétrica.
Por último, Phillips et al. (2011) señalan que el valor de biomasa aérea total en bosques montanos en Colombia corresponde a una media de 131,9 Mg ha-1 en un intervalo de 106,4 y 145,5 Mg ha-1. También, Pérez y Díaz (2010) exponen cifras de biomasa aérea en fustales que varían entre 76,2 y 151,6 Mg ha-1 para bosques en Santander, donde el principal aporte de dicha variable lo proporciona W. tomentosa; así mismo, en el caso de Cundinamarca, se registran de 62,8 a 171,9 Mg ha-1 con un rol significativo de Q. humboldtii. En suma, Rodríguez-Alarcón et al. (2018) reportaron biomasa aérea en bosques de La Calera y Guasca, hallando intervalos de 141,4 a 244,4 Mg ha-1.
Los resultados de otras investigaciones en biomasa aérea se posicionan en niveles inferiores a los encontrados en este estudio (206,22 ± 49,37 Mg ha-1). Las existencias encontradas son similares a las reportadas por Álvarez et al. (2017) en bosques andinos con Q. humboldtii en el norte de Sudamérica (229,9 ± 85,8 Mg ha-1), taxón que coincide con la dominancia, a nivel de biomasa, de la vegetación arbórea analizada.
Conclusiones y recomendaciones
Las condiciones ecológicas de la vegetación arbórea del Parque Natural Chicaque corresponden a bosque secundario de desarrollo tardío, teniendo en cuenta los resultados encontrados en composición, diversidad alfa y estructura. No obstante, los valores determinados proporcionan un marco de referencia favorable en el reconocimiento del servicio ecosistémico de regulación climática que presta la zona, a través del secuestro de carbono en la biomasa aérea.