Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.10 n.2 Bogotá jul./dez. 2008
ARTÍCULO DE INVESTIGACIÓN
Antagonismo in vitro de Trichoderma harzianum Rifai sobre Fusarium solani (Mart.) Sacc., asociado a la marchitez en maracuyá
Trichoderma harzianum Rifai in vitro antagonism against Fusarium solani (Mart.) Sacc., associated to withering in passion fruit
Carol Libeth Suárez Meza1 , Reinel José Fernández Barbosa2 , Nelson Osvaldo Valero3 , Rocío Margarita Gámez Carrillo4 , Alberto Rafael Páez Redondo5
1Microbióloga, Universidad Popular del Cesar (UPC). suarezaser@gmail.com
2Microbiólogo, Universidad Popular del Cesar (UPC). reifer6@gmail.com
3MSc. Microbiología, Universidad Popular del Cesar (UPC). valeronelson@gmail.com
5MSc. Fitopatología, Instituto Colombiano Agropecuario (ICA). alberto.paez@ica.gov.co
Recibido: abril 11 de 2008 Aprobado: octubre 23 de 2008
Resumen
Cultivos de maracuyá de la región Caribe colombiana presentan problemas de marchitez y el hongo Fusarium solani (Mart.) Sacc., está asociado a esta patología. Plantas de maracuyá de la Estación Experimental (E.E Caribia de Corpoica) poseen este problema y por tal razón se determinó el antagonismo in vitro de aislamientos de Trichoderma harzianum Rifai, frente a F. solani, aislado de plantas enfermas de maracuyá (Passiflora edulis). Utilizando la técnica de cultivo dual en platos Petri con Agar Sabouraud, se evaluaron competencia por nutrientes y espacio, micoparasitismo y porcentaje de inhibición del crecimiento radial (PICR), empleando un diseño estadístico factorial 2x6x1 con arreglo completamente aleatorio. Se obtuvieron tres aislamientos nativos de T. harzianum (TCN-009, TCN-010, TCN-014) de suelo de Palma de Aceite (Elaeis guineensis) de la E.E Caribia, y se compararon con tres aislamientos comerciales (TCC-001, TCC-005, TCC-006). TCC-001 y TCN-014, reportaron ser más competentes por nutrientes y espacio, con el mayor radio de crecimiento de 7,50 y 7,32 cm el día 10, comparado a FSM-011 en el cual solo fue de 2,30 cm. Aunque, TCN-014 mostró micoparasitismo grado 4 con ambos aislamientos de F. solani y TCC-005 únicamente con FSM-012, el cual fue más susceptible a ser micoparasitado. En cuanto al PICR, los tratamientos con mejores porcentajes de inhibición fueron TA-9, TA-12 y TA-6 con valores de 70,56, 68,52 y 65,32% respectivamente. El aislamiento del patógeno mayormente inhibido fue FSM-011. Todos estos resultados demuestran que hubo antagonismo in vitro al utilizar los aislamientos nativos y comerciales de T. harzianum sobre F. solani.
Palabras clave: Passiflora, competencia, micoparasitismo, inhibición, antagonismo.
Abstract
Passion fruit cultivation of the Colombian Caribbean region presents problems withering and the fungus Fusarium solani (Mart.) Sacc., this associated with this pathology. Plant of passion fruit of the experimental station of Corpoica Caribia EE. presented this problem and for that reason it was determined the antagonism in Vitro of isolations of Trichoderma harzianum Rifai, compared with F. solani, isolated from diseased plant of passion fruit (Passiflora edulis). Using the technique of cultivation dual petri dishes with Sabouraud agar was evaluated competition for space and nutrients, mycoparasitism and percentage of inhibition of radial growth (PICR), using a factorial design 2x6x1 under completely random. Three native isolations of T. harzianum were obtained (TCN-009, TCN-010, TCN-014) in soil of oil palm (Elaeis guineensis) in Caribia EE and were compared with three commercial isolations (TCC-001, TCC-005, TCC-006). TCC- 001 and TCN-014 resulting be more competent for nutrients and space, the native isolations with the highest growth of 7.50 and 7.32 radius of centimeters the day 10, compared to FSM-011 in which was only 2.30cm. Although, TCN-014 showed mycoparasitism degree 4 with both isolations of F. solani and TCC-005 only with FSM-012 which was more susceptible to micoparasitism. As for the PICR, treatments with better percentages of inhibition were TA-9, TA-12 and TA-6 with values of 70.56%, 68.52% and 65.32% respectively. The pathogenic isolation largely inhibited was FSM-011. All these results demonstrate that there was antagonism in Vitro to use the native and commercial isolations of T. harzianum on F. solani.
Key words: Passiflora, competition, mycoparasitism, inhibition, antagonism.
Introducción
El maracuyá es uno de los cultivos tropicales de mayor importancia en Colombia. La pulpa de esta fruta es utilizada para jugos y es exportada. A junio de 2006 las exportaciones colombianas de jugo de maracuyá ascendieron a 1,7 millones de dólares correspondientes a 958 toneladas (Corporación Colombia Internacional, 2006).
La enfermedad que más ataca al maracuyá es conocida como marchitez o fusariosis, esta es ocasionada por el hongo patógeno Fusarium sp., el cual provoca una muerte prematura de la planta por marchitamiento, al que también se asocia la obstrucción de haces vasculares (González et ál., 2002). La enfermedad se encuentra presente en todo el mundo y en los últimos años varias publicaciones señalan a Phytophthora sp. y Rhizoctonia spp. como patógenos asociados con el síndrome, ya que ambos están asociados con daños a la raíz en diferentes cultivos (Durán et ál., 2001; Espinoza y Mendoza 2001; Rico et ál., 2001; Guerrero et ál., 2001; López et ál., 2002). En Colombia se están presentando daños por este hongo en el Valle del Cauca y Meta (Mendivelso, 2003; Instituto Colombiano Agropecuario, 2000; Cifuente, 2000), reporta a F. solani como agente causal de la fusariosis en tomate (Lycopersicon esculentum, Mill).
El género Trichoderma posee buenas cualidades para el control de enfermedades en plantas causadas por patógenos fúngico Fusarium y otros (González et ál., 2002). Este hongo actúa por medio de una combinación de competencia por nutrientes, producción de metabolitos antifúngicos, enzimas hidrolíticas y micoparasitismo, además de producir sustancias promotoras del crecimiento vegetal (Stefanova 1996). El presente trabajo de investigación tuvo como objetivo determinar la efectividad antagónica in vitro de aislamientos nativos y comerciales de T. harzianum contra F. solani causante de la marchitez de las plántulas del maracuyá en la zona bananera colombiana.
Materiales y métodos
El estudio se realizó en el Laboratorio de Microbiología y Fitopatología de la Estación Experimental (E.E) Caribia de Corpoica. Se utilizaron aislamientos comerciales de T. harzianum provenientes del Centro de Investigación El Roble S.A. (TCC-001, TCC-005 y TCC-006), los aislamientos FSM-011 y FSM-012, pertenecientes a Fusarium solani, fueron aislados de plántulas enfermas del maracuyá de la E.E. Caribia por el protocolo del Fusarium Research Center de la Universidad de Pennsylvania, utilizado por Burges et ál. (1994), e identificados por el método usado por Gilchrist-Saavedra et ál. (2005) y las claves taxonómicas de Booth (1971), Seifert (2001), Arikan et ál. (2001), y Seifert (1996).
Aislamiento e identificación del antagonista nativo
Se muestrearon 39 ha de cultivo de palma de aceite (Elaeis guineensis) de esta misma estación, de forma aleatoria, tomándose dos submuestras por ha (76 en total), y luego fueron mezcladas para obtener una muestra compuesta por cada ha (39 muestras). Estas se seleccionaron utilizando un patrón de recorrido en zig-zag (Carrillo et ál., 1995), de las cuales se hicieron diluciones seriadas de 10-1-10-3, tal como recomienda Monzón (2001). Se sembraron por triplicado alícuotas de 0,1 ml de cada dilución en la superficies de cajas Petri de 9 cm de diámetro con Agar Papa Dextrosa (PDA), y se incubaron a 25±1 ºC, 12 h continuas en luz, seguido de 12 h de oscuridad continua por 5 días, con monitoreo constante. Los aislamientos de T. harzianum fueron contabilizados y purificados, para lo cual se trasladó un disco de agar de 1 cm de diámetro con micelio del hongo, a cajas Petri con agar PDA. Fueron incubados a 25±1 ºC por 5-6 días. La identificación de los aislamientos se realizó utilizando como ayuda el método descrito por Gilchrist et ál. (2005), y claves propuestas por Samuels et ál. (2007), Sutton et ál. (1998) y Ellis (2006).
Confrontación in vitro de cepas de Trichoderma harzianum vs. Fusarium solani
Las pruebas de enfrentamiento se realizaron en Agar Sabouraud Dextrosa (ASD) de OXOID® pH 5,5, colocándose en un extremo de la caja de Petri un disco de Agar de 4 mm de diámetro con micelio de F. solani, y en el extremo opuesto otro disco de 4 mm con micelio de T. harzianum (nativos o comerciales), a una distancia de 5 cm aproximadamente entre ellos (Howell, 2003); posteriormente se incubaron bajo las mismas condiciones del antagonista durante 10 días, haciéndose mediciones cada 24 h del crecimiento radial del micelio de la colonia de los hongos. Sin embargo, se tuvieron en cuenta solo los valores del día 10 ya que las anteriores no muestran diferencias significativas entre los tratamientos. Se sembró en cajas separadas un inóculo de cada antagonista y de cada patógeno, los cuales correspondieron a los controles y fueron incubados bajo las condiciones anteriormente mencionadas. Los tratamientos evaluados pertenecieron a los cultivos duales, controles de antagonistas y controles de patógenos (20 en total).
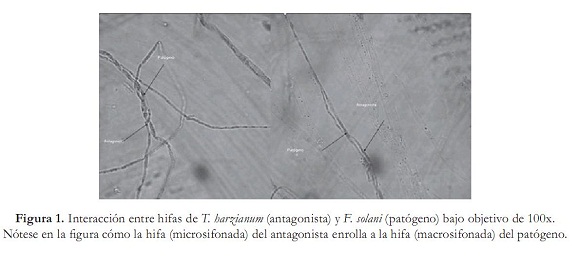
El antagonismo o capacidad antagónica de aislamientos nativos o comerciales de T. harzianum se comprobó de distintas formas: primero midiendo el grado de micoparasitismo de cada antagonista, el cual se determinó por la escala que proponen Ezziyyani et ál. (2004) (tabla 1). Además, se evaluó la interacción de las hifas por observación en un microscopio binocular ZEIZZ con aumento de 100x.
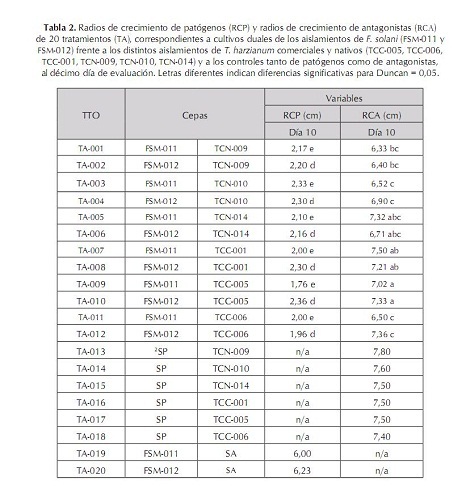
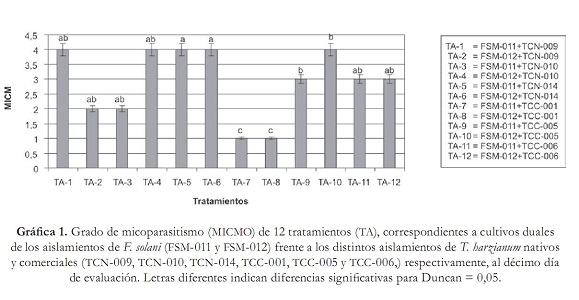
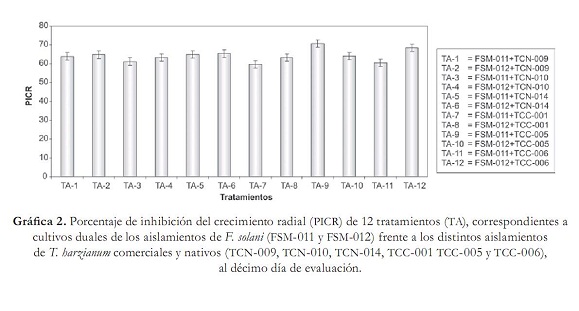
Segundo, por la evaluación de competencia por nutrientes y espacio, la cual se obtuvo con los radios de crecimiento de cada patógeno y antagonistas en cultivo dual, junto con sus respectivos testigos, utilizando un calibrador "Pie de rey", y tercero, por el porcentaje de inhibición de crecimiento radial (PICR), con la fórmula utilizada por Ezziyyani et ál. (2004), PICR = (R1 – R2)/R1 x 100, donde R1 es el radio mayor (radio patógeno testigo) y R2 es el radio menor (radio del patógeno en enfrentamiento con el antagonista). El análisis estadístico se realizó haciendo un análisis de varianza (Anova) y prueba de comparación de promedios Duncan con el programa estadístico Mstat SAS versión 6.12, bajo un diseño experimental completamente al azar con arreglo factorial 2x6x1, una caja Petri como unidad experimental, y tres repeticiones. Las variables que se midieron fueron: radio de crecimiento del antagonista (RCA), radio de crecimiento del patógeno (RCP), micoparasitismo (MICMO) y PICR. Además, los factores estudiados fueron: factor patógeno FUS (FSM-011, FSM-012), factor antagonistas TRICH (TCN-009, TCN-010, TCN-014, TCC-001, TCC-005, TCC-006), y el factor DIA (10). Resultados y discusión Aislamiento e identificación del antagonismo nativo Se obtuvieron los aislamientos TCN-009, TCN-010, TCN-014 pertenecientes a T. harzianum, los cuales crecieron rápidamente y con aspecto de polvillo; las colonias eran de color blanco con verde olivo; se observaron al microscopio fialides cortos en forma de botella, con un conidio sucesivo en la extremidad, y conidias globosas de color verde. Estas características concuerdan con las descritas por Samuels et ál. (2007) para Trichoderma harzianum. Confrontación in vitro de cepas de Trichoderma harzianum vs. Fusarium solani Todos los tratamientos evaluados de T. harzianum presentaron radios de crecimiento (RCA) superiores al de los aislamientos patógenos de F. solani (RCP). Sin embargo, los tratamientos TA-7, TA-12, TA-10 y TA-5 fueron aquellos donde los antagonistas desarrollaron el mayor crecimiento con valores de 7,5; 7,36; 7,33 y 7,32 cm respectivamente, valores que fueron muy próximos a los reportados por sus controles en TA-16, TA-18, TA-17 y TA-14 (tabla 2). Además, se observó claramente que los RCP fueron muy inferiores con respecto a los radios de crecimiento que presentaron los antagonistas y sus respectivos controles. Como se observa en TA-12 donde FSM-012 creció tan solo 1,96 cm, seguido de TA-7, donde se presentó un radio de 2 cm para FSM-011 y en TA-5, en el cual el RCP de FSM-011 midió 2,1 cm por debajo del diámetro reportado por FSM-012 de TA-10 con 2,3 cm (tabla 2). Esto indica que probablemente, T. harzianum pudo tener una tasa de incorporación de nutrientes, tasa de metabolismo y un crecimiento superior a F. solani, utilizando distintos mecanismos como secreción de enzimas hidrolíticas, entre ellas celulasas, quitinasas, glucanas, xylasas y muchas veces las proteasas, las cuales pueden estar implicadas en los mecanismos de biocontrol, permitiéndole al antagonista aprovechar mejor los nutrientes del medio y privar al patógeno de utilizar los recursos (Michel, 2001). Aunque se haya presentado mayor eficiencia a nivel experimental en la competencia por nutrientes y espacio por parte de los aislamientos comerciales debido a que fueron los aislamientos que mostraron mayor RCA frente a los aislamientos patógenos, no existieron diferencias significativas entre el aislamiento nativo TCN-014 (TA-05) y los aislamientos comerciales TCC-001 (TA-07), TCC-005 (TA-010) y TCC-006 (TA-012) (tabla 1). En cuanto al micoparasitismo, solo los tratamientos TA-1, TA-4, TA-5, TA-6 y TA-10 presentaron micoparasitismo completo en grado 4 (gráfica 1). Cabe destacar que únicamente se observó micoparasitismo en grado 4 para ambos aislamientos de F. solani en los tratamientos TA-5 y TA-6, pertenecientes a TCN-014, mientras que en TA-1, TCN-009 mostró micoparasitismo completo solamente con FSM-011, como también sucedió en TA-4 (TCN-010) y TA-10 (TCC-005), frente a FSM- 012 de F. solani (Grafica 1). Esto se debió probablemente a que, como lo propone Tronsmo (1996), Trichoderma sp. tiene la capacidad de conducir sus hifas hacia las de otros hongos, enrollándose en ellas, coincidiendo con algunos montajes microscópicos realizados donde se observaron entrelazamientos de hifas (figura 1). Lo que ocurre en reacción mediada por lecitinas que van degradando la pared celular del hospedero, y por la secreción de exoenzimas hidrolíticas. Dicho proceso limita el crecimiento y la actividad del hongo patógeno y algunas especies de Trichoderma sp. son conocidas por producir elevadas concentraciones de enzimas degradadoras de la pared celular, tales como a-1,3-glucanasa y diferentes enzimas quitinolíticas que actúan en conjunto con proteasas y celulasas (Schickler y Chet 1997; Michel, 2001). Por esta razón, es necesario continuar confirmando el micoparasitismo de los aislamientos de T. harzianum para especificar exactamente cuáles enzimas intervienen en el proceso. Existieron diferencias significativas (P=0,05) entre los aislamientos TCN-014, TCN-009 y TCC-005. Los mejores resultados de micoparasitismo in vitro se dieron cuando se empleó TCN-014, por ser de los tres aislamientos el que lo hizo en ambos aislamientos del patógeno (gráfica 1). En el porcentaje de inhibición del crecimiento radial (PICR), es importante resaltar que todos los tratamientos presentaron una inhibición del crecimiento radial del patógeno superior al 50% (gráfica 2). Resultados similares reporta Michel A. (2001), quien halló en su trabajo un nivel de PICR de 69,5%. El PICR en esta investigación varió para los aislamientos comerciales y nativos. Los mayores pertenecieron a los tratamientos TA-9 (TCC-005) con un valor de 70,6% correspondiente al aislamiento comercial del antagonista frente a FSM-012 y TA-11 (TCN-014) con 65,32% perteneciente al aislamiento nativo del antagonista frente a FSM-011 (gráfica 2). Trichoderma harzianum es capaz de inhibir del crecimiento de fitopatógenos porque produce numerosos antibióticos como la trichodermina, la suzukacilina, la alameticina, la dermadina, la penicilina, los trichotecenos, las trichorzianinas, entre otros (Durán et ál., 2003), probablemente estos aislamientos antagónicos utilizaron este método de supresión contra el patógeno. Conclusión El aislamiento nativo TCN-014, seguido del aislamiento comercial TCC-005 de Trichoderma harzianum, resultaron ser los antagonistas con mayores habilidades de micoparasitismo, competencia por espacio e inhibición de crecimiento para el control de los aislamientos patógenos FSM-011 y FSM-012 de Fusarium solani de acuerdo con las pruebas in vitro. Recomendaciones Agradecimientos A la Corporación Colombiana de Investigación Agropecuaria (Corpoica) Estación Experimental Caribia, Compañía Envasadora del Atlántico, Centro de Investigación El Roble S.A. y Universidad Popular del Cesar. Referencias bibliográficas 1 Arikan, S.; Lozano, M.; Paetznickv, V. y Rex, J. 2001. In Vitro susceptibility testing methods for caspofungin against Aspergillus and Fusarium isolates. Antimicrob Agent Chemother 45: 327-30. [ Links ] 2 Booth, C. 1971 The Genus Fusarium. CMI. Kew, Surrey 19-31. [ Links ] 3 Burges, B.; Ummerell, S.; Bullock, K.; Gott y Backhouse, P. 1994. Laboratory Manual for Fusarium Research. University of Sydney and Botanic Gardens, Department of Crop Science, Fusarium Research Laboratory, Sydney, Australia 191. [ Links ] 4 Carrillo, F. I.; Suárez, J. y Sanz, R. 1995. Cómo obtener una buena muestra para el análisis de suelos. Cenicafé. Avances Técnicos 214: 1-4 . [ Links ] 5 Cifuente, J. 2000. Evaluación de la capacidad biocontroladora del hongo Trichoderma hazianum cepa nativa Queule sobre Fusarium solani en tomate (Lycopersicon esculentum Mill). Universidad de Talca. [ Links ] 6 Corporación Colombia Internacional (CCI). 2006. Información de monitoreo internacional. Maracuyá. SIM. Inteligencia de mercados. [ Links ] 7 Durán, E.; Robles, F.; Martínez, J. y Brito, M. 2003. Trichoderma Un hongo combatiente de patógenos. Revista Técnico Ambiental Teorema Ambiental 42: 23 -26. [ Links ] 8 Durán Ortiz, L. J.; Pérez, M. L. y Sánchez Pale, J. R. 2001. Identificación de los hongos que ocasionan la marchitez del chile en la región del bajío. Querétaro: XXVIII Congreso Nacional de Fitopatología. [ Links ] 9 Ellis D. 2006. The University of Adelaide Australia. Mycology [Online]. Disponible en http://www.mycology.adelaide.edu.au/Fungal_Descriptions/Hyphomycetes_(hyaline)/ Trichoderma/ [ Links ] 10 Espinoza López, L. y Mendoza Zamora, C. 2001. Etiología de la pudrición de raíz y cuello del chile (Capsicum annuum L.) ocasionado por el hongo Phytophthora capsici en la región de Valseguillo, Puebla, México. Fitopatología 30: 47-55. [ Links ] 11 Ezziyyani, M.; Pérez, S. C.; Requena, M. E.; Rubio, L. y Candela, M. E. 2004. Biocontrol por Streptomyces rochei -Ziyani-, de la podredumbre del pimiento (Capsicum annuum L.) causadapor Phytophthora capsici. Anales de Biología 26: 69-78. [ Links ] 12 Gilchrist Saavedra, L. G.; Fuentes Dávila, C.; Martínez Cano, R. M.; López Atilano, E.; Duveiller, R. P.; Singh, M.; Henry, I. y García, A. 2005. Guía práctica para la identificación de algunas enfermedades de trigo y cebada. 2 edición. México, D. F.: CIMMYT. [ Links ] 13 González Chavira, M. M.; Torres Pacheco, I. y Guzmán Maldonado, H. 2002. Búsqueda de resistencia natural contra patógenos de raíz Phytophthora capsici, Fusarium solani y Fusarium oxysporum en colectas de chile. Tampico: Proceedings of the 16th Internacional Pepper. [ Links ] 14 Guerrero Aguilar, Z.; Sánchez Delgadillo, F.; Guevara Olvera, L.; Guevara González, R.G.; Torres Pacheco, I. & González Chavira, M. M. 2001. Caracterización de aislados mexicanos de Rhizoctonia solani (Kuhn). Querétano: XXVIII Congreso Nacional de Fitopatología. [ Links ] 15 Howell, C. R. 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Disease 87: 4-10. [ Links ] 16 Instituto Colombiano Agropecuario (ICA). 2000. Red de vigilancia epidemiológica en cultivos como café, ornamentales, cítricos y otras frutas, plátano y banano, cacao y algodón. Boletín Epidemiológico Agrícola. [ Links ] 17 López Vázquez, M. A.; Martín González, C. M.; Torres Pacheco, I.; Delgadillo Sánchez, F. y Guevara González, R. G. 2002. Patógenos involucrados en la pudrición de raíz del chile. Monterrey: XXIX Congreso Internacional de Fitopatología. [ Links ] 18 Mendivelso, B. N. 2003. A la sombra del maracuyá. UN Periódico, Disponible en http://www.biodiversityreporting.org/article.sub? docId= 728 &c=Col %C3%83%C6%92%C3%82%C2%B4mbia& cRef=Colombia &year =2003& date =maio%20 [ Links ] 19 Michel, A. 2001. Cepas nativas de Trichoderma spp. (Euascomycetes:Hypocreales), su antibiosis y micoparasitismo sobre Fusarium subglutinans y F. oxysporum (Hyphomycetes:Hyphales). México: Universidad de Colima. [ Links ] 20 Monzón, A. 2001. Producción, uso y control de calidad de hongos entomopatógenos en Nicaragua. Avances en el fomento de productos fitosanitarios no- sintéticos. Manejo Integrado de Plagas. Costa Rica 63: 95-103. [ Links ] 21 Rico Guerrero, L.; Guerrero Aguilar, B. Z.; López Vázquez, A.; Muñoz Sánchez, C. I.; Guevara Olvera, L.; Guevara González, R. G.; Torres Pacheco, I. y González Chavira, M. M. 2001. Búsqueda de resistencia natural en plantas de chile (Capsicum spp.) contra aislados del complejo fúngico que causa pudrición de raíz. Querétaro: XXVIII Congreso Nacional de Fitopatología. [ Links ] 22 Samuels, G. J.; Chaverri, P.; Farr, D. F. y McCray, E. B. 2007. Trichoderma Online, Systematic Botany & Mycology Laboratory. Disponible en http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm [ Links ] 23 Schickler, H. y Chet, I. (1997). Herologous chitinase gene expresión to improve plant defense against phytopathogenia fungi. Journal of Industrial Microbiology & Biotecnology 19: 196-201. [ Links ] 24 Seifert, K. 1996. Fuskey. Fusarium interactive key. Agriculture and agi-food Canada. Disponible en http://www.ctu.edu.vn/colleges/agri/gtrinh/fuskey.pdf [ Links ] 25 Stefanova Nalimova, M. 1996. Producción y aplicación de Trichoderma spp. como antagonista de hongos fitopatógenos. La Habana: Instituto de Sanidad Vegetal. [ Links ] 26 Sutton, D. A.; Fothergill, A. W. y Rinaldi, M. G. 1998. Guide to clinically significant fungi, Baltimore: Williams & Wilkins, Phytopathol. [ Links ] 27 Tronsmo, A. 1996. Trichoderma harzianum in biological control of fungal diseases. In: R. Hall (ed.). Principles and practices of managing soilborne plant pathogens. pp. 213-236. [ Links ]