










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.10 n.2 Bogotá jul./dez. 2008
ARTÍCULO DE INVESTIGACIÓN
Toxicidad de δ-endotoxinas recombinantes de Bacillus thuringiensis sobre larvas de la polilla guatemalteca (Tecia solanivora) (Lepidóptera: Gelechiidae)
Toxicity of Bacillus thuringiensis recombinant δ-endotoxins on Guatemalan potato moth (Tecia solanivora) (Lepidoptera: Gelechiidae) larvae
Leanis Pitre Ruiz1 , Javier Hernández-Fernández2 , Jaime Bernal Villegas3
1Universidad de La Guajira.
2Docente-investigador, Universidad Jorge Tadeo Lozano. javier.hernandez@utadeo.edu.co
3Director, Instituto de Genética, Pontificia Universidad Javeriana.
Resumen
Con el objetivo de determinar la actividad tóxica específica de las proteínas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C de Bacillus thuringiensis (Bt), sobre larvas de primer instar de Tecia solanivora se estableció la CL50 para las toxinas. Para este fin se implementó la cría masiva de este insecto bajo condiciones de laboratorio, 58±5% de humedad relativa, 18±5ºC de temperatura y un fotoperiodo de 23 h oscuridad y 1 h luz. Se utilizó una dieta seminatural consistente en láminas de papa variedad parda pastusa autoclavada con solución preservante (ácido ascórbico 7 g/L y metilparabeno 5 g/L), ya que fue estable en el tiempo, garantizó la reproducibilidad de los resultados y fue de fácil evaluación. Las proteínas recombinantes se evaluaron a una concentración de 0,1 µg/cm2. Los resultados obtenidos de porcentaje de mortalidad indicaron que no había diferencias entre las delta-endotoxinas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C de Bt (P<0,01 y P<0,05). Se calculó la CL50 de las cuatro toxinas, evaluándolas a concentraciones de 0,05; 0,1; 0,15; 0,2; 0,25 y 0,3 µg/cm2 de proteína, obteniendo las CL50: Cry1Aa 0,103, Cry1Ac 0,107, Cry1B 0,085 y Cry1C 0,112 µg de proteína/cm2. Se comprobó que las δ-endotoxinas evaluadas poseen una alta toxicidad sobre larvas de T. solanivora, siendo levemente mayor la proteína Cry1B. Este resultado es promisorio para posteriores investigaciones en ingeniería genética para la posibilidad de obtener cultivos de papas transgénicas resistente a T. solanivora
Palabras clave: Bacillus thuringiensis, proteínas recombinantes, Tecia solanivora, papa parda pastusa.
Abstract
The specific toxicity of Cry1Aa, Cry1Ac, Cry1B and Cry1C recombinant proteins from Bacillus thuringiensis (Bt) was determined on Tecia solanivora first instar larvae. Mass breeding of this insect was established in laboratory conditions: 58 ± 5% relative humidity, 18 ± 5ºC temperature and 23 h dark and 1 h light photoperiod. A semi-natural diet was supplied consisting of brown pastusa variety potato slices sterilised with preserver solution (7 g/L ascorbic acid and 5 g/L methylparabene) as it was steady over time, thereby guaranteeing the reproducibility of results and was easy to assess. The recombinant proteins were also evaluated at 0.1 µg/cm2 concentration to find significant differences between them. Mortality percentage results indicated that there were no differences between the B thuringiensis Cry1Aa, Cry1Ac, Cry1B and Cry1C delta-recombinant endotoxins (p<0.01 and p<0.05); however, difference s were seen when comparing them with the reference B. thuringiensis var. kurstaki HD1 strain. CL50 was calculated for the four toxins, testing at 0.05, 0.1, 0.15, 0.2, 0.25 and 0.3 µg/cm2 protein concentration. Probit Polo PC software was used for determining that the CL50 obtained inside this rank was: Cry1Aa 0.103, Cry1Ac 0.107, Cry1B 0.085 and Cry1C 0.112 µg protein/cm2. It was clear that the evaluated d-endotoxins had high toxicity on T. solanivora larvae, the Cyr1B protein being slightly stronger. This result is promissory for further research on genetic engineering aimed at obtaining transgenic T. solanivora-resistant potato crops.
Key words: Bacillus thuringiensis, bioassay, recombinant endotoxin, Tecia solanivora, pastusa potato variety.
Introducción
La papa (Solanum tuberosum) es uno de los cultivos más importantes en Colombia, en el año 2004 ocupó el quinto lugar en la producción agropecuaria nacional con 2,8 millones de Tm. (Cevipapa, 2004), fue el noveno cultivo en extensión con 161.873 ha, y el sexto en valor de la producción (Espinal et ál., 2006). Alrededor de 90.000 familias se encuentran vinculadas con la explotación directa de este cultivo, y se generan cerca de 20 millones de jornales al año (Espinal et ál., 2006). El cultivo de la papa es atacado por una gran variedad de insectos plagas, en donde se destaca Tecia Solanivora (povolni). o "polilla guatemalteca" (Lepidóptera: Gelechiidae), esta polilla se le reconoce actualmente por ser un insecto plaga de la papa en Sur y Centroamérica. En Guatemala fue detectado por primera vez en 1956 y se expandió a Suramérica por medio de la importación de semillas contaminadas realizadas por Venezuela en 1983. En Colombia se identificó en 1985 (Pollet, 2001; Pollet et ál., 2003). Hace algunos años se detectó en Islas Canarias (Tenerife), España (EPPO, 2001), produciendo pérdidas cuantiosas (MacLeod, 2005). Desde entonces, la polilla guatemalteca se introdujo en la lista roja de la Organización para la Protección de Plantas de Europa y el Mediterráneo (EPPO, 2001). Desde su introducción se ha producido un fuerte impacto económico, ya que esta plaga en su estado larval causa daños directos a los tubérculos tanto en campo como en almacén, afectando la calidad de los mismos y ocasionando grandes pérdidas económicas, entre el 30-50% de los costos totales de producción (Sánchez, 2000). Para el control de T. solanivora los agricultores utilizan principalmente plaguicidas químicos, los cuales no realizan un control eficiente de esta plaga (Rodríguez, 1996). Sin embargo, sí generan una serie de problemas secundarios como resistencia de la plaga a los insecticidas, destrucción de los enemigos naturales y presencia de residuos químicos en los tubérculos.
Esta situación ha estimulado investigaciones que pretenden explorar y encontrar otras alternativas de control de plagas, entre éstas, el control biológico mediante el uso de microorganismos entomopatógenos, alternativa de menor impacto ecológico. Entre los microorganismos más utilizados para el control biológico de insectos plaga se destaca Bacillus thuringiensis (Bt), una bacteria Gram positiva, esporulada, nativa del suelo, caracterizada por la producción de cristales paraesporales compuestos por proteínas insecticidas (ICP) (Schnepf et ál., 1998). La mezcla de esporas e ICP de varias cepas de Bt se ha utilizado como principio activo de formulaciones comerciales para el control de insectos lepidópteros, coleópteros y dípteros plaga, en una amplia gama de cultivos agrícolas desde hace casi medio siglo (Schnepf et ál., 1998).
Identificar las ICP tóxicas sobre T. solanivora es de gran importancia para un programa de mejoramiento genético que tenga como objetivo la producción de plantas resistentes al ataque de esta plaga (Estrada, 1997; Shelton et ál., 2002). Las proteínas Cry recombinantes se utilizan para determinar la toxicidad individual en bioensayos corrientes, ya que la mayoría de cepas de Bt poseen más de una toxina y al realizar bioensayos con éstas no se puede saber cuál es la proteína responsable de la actividad tóxica, lo que sí se puede revelar utilizando proteínas Cry recombinantes. Es muy posible la aparición de resistencia a una determinada toxina por parte de un insecto plaga, considerando que muchos de los casos reportados de alta resistencia a proteínas Cry están relacionados con alteraciones de los receptores de membrana en el intestino medio (Ferre y Van Rie, 2002), el estudio desarrollado provee la posibilidad de utilizar diferentes proteínas Cry para el control de Tecia solanivora y de esta manera controlar o retrasar la aparición de resistencia.
Dado que diferentes proteínas insecticidas de B. thuringiensis difieren en su potencia y especificidad al huésped, se justifica la evaluación de la actividad tóxica de esta sobre larvas de Tecia solanivora para su control y, por consiguiente, para utilizarse como principio activo en la producción de bioinsecticidas de mayor efectividad y espectro de acción, y la producción de plantas transgénicas autorresistentes al ataque de esta plaga. Por tal motivo, en el presente trabajo se determinó la actividad tóxica de las proteínas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C de Bacillus thuringiensis sobre larvas de T. solanivora como una alternativa de control de este insecto plaga.
Materiales y métodos
Proteínas recombinantes
Se utilizaron las proteínas recombinantes Cry1Aa, Cr1Ac, Cry1B y Cry1C de Bt cedidas gentilmente por el doctor Ruud A. de Maagd (Plant Research International P.O. Box 16 6700 AA Wageningen, Holanda). Todas las protoxinas de aproximadamente 120-130 kDa se activaron con un tratamiento con tripsina y se purificaron por cromatografía de columna de intercambio iónico MonoQ HR 5/5 (FPLC system), y se resuspendieron en un buffer que contenía 20 mM Tris-HCL pH 9, 250 mM y NaCl pH 9.
Electroforesis de proteínas
Se realizó un gel de poliacrilamida (SDS-PAGE) al 12%, para visualizar y determinar la integridad de las δ-endotoxinas. El corrido electroforético se llevó a cabo en una cámara de electroforesis vertical Mini-PROTEAN II (BIO RAD). Las muestras se prepararon siguiendo la metodología propuesta por Laemmli (1970). El corrido electroforético se llevó a cabo a 150 voltios durante 60 minutos y el gel fue teñido con Azul de Coomassie R-250 al 0,1%durante toda la noche. Posteriormente, el gel se decoloró en una solución de metanol-ácido acético (40-10%) durante 4 horas.
Cepa bacteriana, cultivo y cuantificación de proteínas totales
Se utilizó la cepa de referencia Bacillus thuringiensis var. kurstaki HD-1, proveniente del Institute Entomopathogenic Bacillus Center (Instituto Pasteur, París). La cepa se sembró en medio Luria Bertani líquido (LB) modificado (10 g/L NaCl, 10 g/L triptona, 7,5 g/L extracto de levadura, 4,0 g/L de sulfato de amonio), se incubó a 30 oC por 7 días hasta alcanzar su esporulación. La biomasa se recuperó por centrifugación a 4000 rpm por 20 min, y se obtuvo un extracto crudo de esporas y cristales después de sucesivos lavados con agua destilada desionizada estéril. Para la cuantificación de proteína total de la mezcla espora-cristal se utilizó el método de Bradford (1976), utilizando como patrón estándar una curva de calibración con albúmina sérica bovina (BSA).
Material entomológico y cría del insecto
La población inicial del insecto utilizada para los bioensayos provino de la sección de Entomología del Programa Nacional de Manejo Integrado de Plagas Corpoica-Tibaitatá (Cundinamarca). Se utilizaron larvas de primer instar de T. solanivora obtenidas a partir de una cría masiva establecida y mantenida a 58±5% de humedad relativa, 18±5ºC de temperatura y un fotoperiodo de 23 h oscuridad y 1 h luz, en dieta natural consistente en tubérculos de papa parda pastusa. Las ovoposiciones se recolectaron semanalmente y se colocaron en cajas de Petri estéril para esperar la eclosión de los huevos y la emergencia de las larvas. Las larvas de primer instar se utilizaron en los bioensayos.
Como cámara de cría se usaron recipientes plásticos translúcidos de 33 x 10 x 22 cm. En cada uno se colocaron 500 g de arena en el fondo, previamente lavada y esterilizada. Dentro de cada cámara se colocó una rejilla que poseía espacios cuadrados de 3 x 3 cm a una distancia de 3 cm por encima de la arena, que sirvió como soporte para colocar 12 papas variedad parda pastusa, las cuales fueron infestadas aproximadamente con 30 huevos del insecto (figura 1). Cada cámara fue cubierta con tapas de plástico cortadas en el centro cubiertas con tela organdí que sirvió para mantener la aireación y previno la salida de los insectos adultos, estas cámaras fueron mantenidas bajo iguales condiciones que la cría.

Los insectos desarrollaron su ciclo larval en los tubérculos, al fi nalizar esta etapa de su desarrollo las larvas empupaban en la arena y después de 20 días los adultos de Tecia solanivora que emergían eran retirados con la ayuda de un tubo de ensayo. Un promedio de 30 adultos eran colocados en recipientes de vidrio de 100 ml, los cuales se taparon con tela organdí. A cada frasco le fue acondicionado un copo de algodón embebido en una solución de sacarosa al 10%, que fue cambiado cada tres días para mantener una dieta constante y evitar la contaminación por hongos. Los frascos con adultos fueron ubicados boca abajo sobre círculos de papel fi ltro para que las hembras adultas ovipositaran. Posteriormente, los huevos se colectaron y se utilizaron para el mantenimiento de la cría o la realización de bioensayos.
Montaje del bioensayo
El bioensayo se realizó utilizando láminas de papa de 1,0 mm de grosor y 5 cm de diámetro. Estos discos de papa se colocaron en cajas de Petri de 60 x 15 mm estériles y se autoclavaron a 15 libras de presión por 1 minuto. Inmediatamente después, la superficie de las papas se impregnó con solución preservante compuesta de ácido ascórbico 7 g/l y metil parabeno 5 g/l, se dejó secar a 35 ºC por 7 horas.
La proteína total de la cepa de Bt kurstaki HD1 o las toxinas recombinantes se colocaron sobre la superfi cie de la papa y se dejaron secar en horno a 35 °C. Luego, nuevamente en la superfi cie de la papa, y en condiciones de esterilidad, se colocaron las larvas de T. solanivora con un pincel de pelo de marta de 0,5 mm. Este montaje se mantuvo bajo condiciones controladas a 18±5 ºC, humedad relativa de 58±5%, por 23 horas de oscuridad y 1 hora de luz.
Evaluación biológica de la cepa de referencia y de las proteínas recombinantes de Bacillus thuringiensis sobre larvas de primer instar de Tecia solanivora
Se realizó un bioensayo general para determinar la concentración de proteína total previa a la realización de la CL50 con y sin extracto crudo de esporas y cristales de Bt. var. kurstaki HD1, se utilizaron dosis de 0, 1, 5 y 10 µg/cm2 de proteína por superficie de dieta, para aproximar la concentración óptima de la toxina por utilizar en los bioensayos con las proteínas recombinantes. Aunque no es fácil considerar esta comparación, se realizó teniendo en cuenta que se utilizó un extracto crudo de la cepa de referencia con una cuantificación indirecta de proteínas totales, y en vez de obtener la dosis letal media (DL50) se trabajó con la concentración letal media (CL50) (microogramos de toxina por gramo de larva), igual al trabajo realizado por Lecadet et ál. (1988). En un segundo bioensayos, utilizando proteínas totales de la cepa de referencia Bt kusrtaki HD1, se utilizaron concentraciones de 0,05; 1; 1,5; 2; 2,5 y 3,0 µg/cm2 y con esta se determinó la CL50.
Las proteínas recombinantes se evaluaron de acuerdo con su toxicidad sobre larvas de 1er instar de T. solanivora, igualmente que el extracto crudo de la cepa de referencia. Primero se realizó un bioensayo con una sola dosis de toxina a 0,8 µg/cm2 y después se realizó una CL50 a concentraciones de 0,05; 1; 1,5; 2; 2,5 y 3,0 µg/cm2
Diseño experimental y análisis estadístico de resultados
Se empleó un diseño completamente al azar con 3 repeticiones, el cual incluyó un testigo absoluto (dieta, tampón y larvas), un testigo tratado (dieta, toxina y larvas), y un control positivo con la cepa de referencia. Para cada una de las repeticiones se utilizaron 30 larvas de primer instar de T. solanivora, para un total de 90 larvas por tratamiento. Además de las 3 repeticiones realizadas por tratamiento, el experimento fue repetido 3 veces en días diferentes. Se realizaron los conteos de mortalidad (inmóviles o cambio de color), a los 7 días. Se realizó el conteo del número de larvas vivas en cada bioensayo y se corrigió la mortalidad del control mediante la fórmula de corrección de mortalidad de Abbott.
Para analizar las interacciones de los factores que presentaron significancia se realizó un análisis de varianza y contrastes ortogonales, este último para hacer una comparación entre las medias de los promedios del porcentaje de mortalidad y las Cl50 de las proteínas recombinantes, y la cepa de referencia en un nivel de significancia del 0,01 y el 0,05%. Para el análisis estadístico de los resultados se utilizó el programa SAS en versión 6.0 Microsas, y para la determinación de los valores de la concentración letal (CL50) se utilizó el análisis de regresión logarítmico Probit de POLO-PC (LeOra Software, 1987) (Finney, 1971).
Resultados y discusión
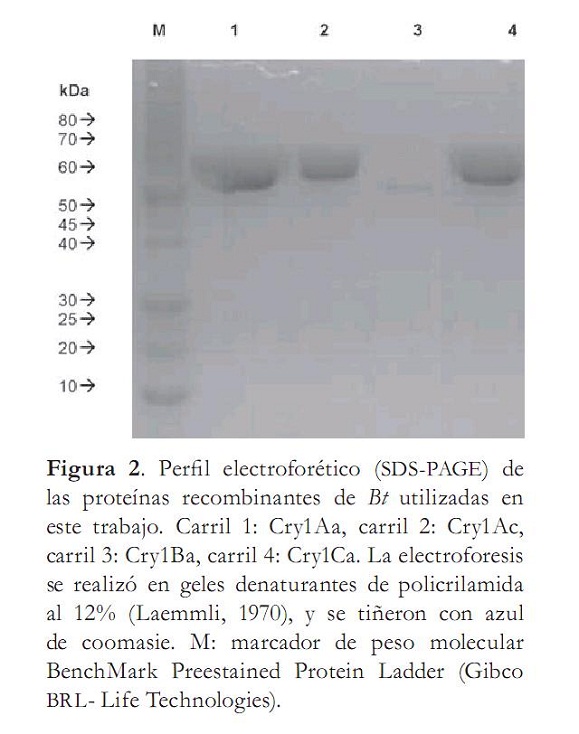
Electroforesis de las proteínas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C, por SDS-PAGE
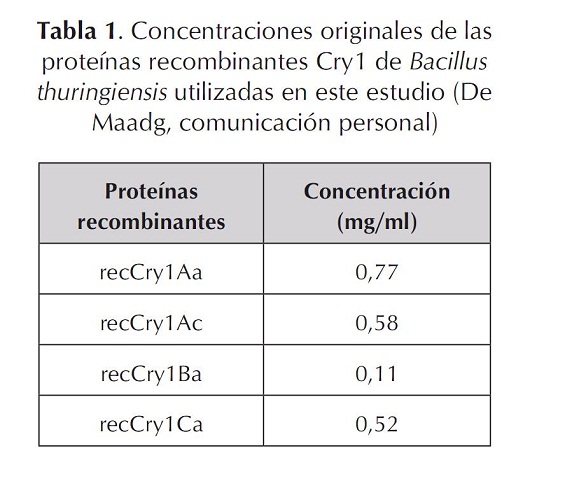
Las proteínas recombinantes mostraron bandas de proteínas con niveles de expresión diferenciales de peso molecular promedio entre 55 y 60 kDa (figura 2, tabla 1). La variación en el peso de las toxinas es bastante sutil; las proteínas Cry1Ac y Cry1Ca presentaron un peso mayor de aproximadamente 60 kDa, con respecto a las Cry1Aa y Cry1B, que muestran un peso aproximado de 50 kDA (figura 1). Estas bandas electroforéticas corresponden a los pesos esperados para estas proteínas Cry en estado clivado o activado de la toxina. Las diferencias en las concentraciones están de acuerdo con la concentración establecida para cada una de las proteínas (De Maagd, Comunicación personal). Las proteínas Cry1Aa, Cry1Ac y Cry1Ca presentan en el gel bandas de concentración parecida que contienen 0,77; 0,58 y 0,52 mg/ml respectivamente, mientras que la proteína Cry1Ba mostró una banda tenue y delgada comparada con las otras proteínas Cry, ya que la concentración de esta fue por lo menos 5 a 7 veces menor (tabla 1). La visualización de las proteínas permitió evidenciar su pureza y su estabilidad en la solución preservante. Adicionalmente, con la concentración determinada se hicieron las diluciones para impregnar superficialmente los discos de papa autoclavados utilizados en los bioensayos.
Mantenimiento de la cría de Tecia solanivora
La polilla tuvo una metamorfosis completa, lo que significa que pasó por todos los estadios de huevo, larva, pupa y adulto. La duración del ciclo fue de tres meses, a una temperatura de 25 ºC y una humedad relativa entre 75-95% (Torres et ál., 1997). Las posturas de huevos realizadas por los insectos adultos fueron de entre 100 a 200 huevos, de los que eclosionaron en promedio cerca del 95%, obteniéndose una buena cantidad de instares larvales para la realización de bioensayos. Se utilizaron larvas de primer instar o neonatas, ya que es el estado más susceptible del insecto, da una respuesta más exacta del nivel de toxicidad de la proteína evaluada, generalmente es el instar para controlar en campo, está disponible en un número más grande que otros instares, el periodo de bioensayo es generalmente corto, y la precisión es más alta ya que la mortalidad es uniforme y los intervalos de confidencia son más pequeños (Navon, 2000).
Evaluación de la concentración letal media de la cepa de referencia y proteínas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C de Bacillus thuringiensis
Se observó que la cepa de Bt kurstaki HD1 en concentraciones de 5,0 y 10 µg/cm2 presentó una mortalidad en las larvas del 100%, y del 55,5% con la concentración de 1,0 µg/cm2 después de 7 días de exposición. Estos resultados preliminares permiten establecer que entre las toxinas producidas por Bt var. kurstaki HD-1 ejercen una actividad tóxica sobre larvas de primer instar de Tecia solanivora (Pérez et ál., 1997; Prias, 1998; Martínez et ál., 2003).
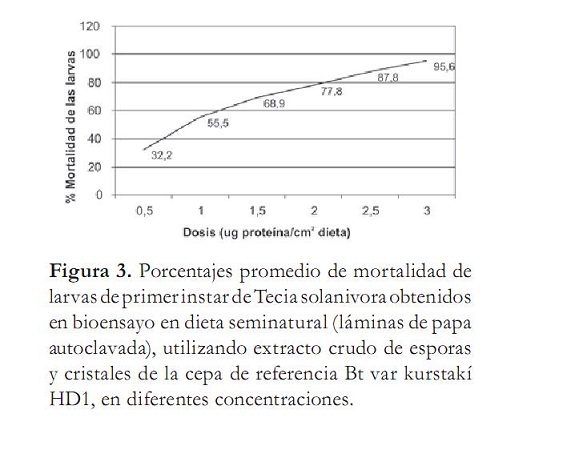
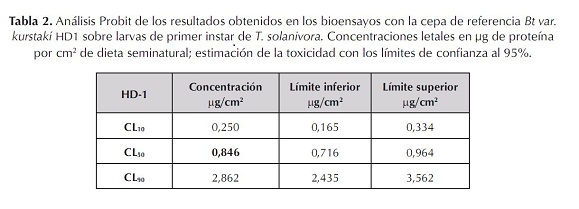
Para calcular la CL50 de la cepa Bt kurstaki HD1 se tomó como referencia el valor obtenido en el ensayo anterior y se realizaron bioensayos con dosis de 0 a 3,0 µg/cm2, en intervalos de 0,5 (figura 3). Los datos obtenidos en los bioensayos se procesaron mediante el paquete estadístico Probit Polo PC (Finney, 1971), encontrándose como CL50 un valor de 0,846 µg de proteína/cm2 de dieta (tabla 2). Este valor de CL50 está muy por debajo del obtenido por Martínez et ál. (2003) y Pérez et ál. (1997), quienes determinaron la CL50 de esta misma cepa sobre larvas de primer instar de T. solanivora en 4,37 y 2,84 µg/cm2 respectivamente. Esta diferencia puede ser atribuible a los distintos métodos de bioensayo empleados, ya que probablemente la dieta utilizada no proporcionó las condiciones adecuadas para obtener un porcentaje de supervivencia de las larvas del 100% en los trabajos previos. Es probable que también infl uyera en la producción de factores tóxicos de la espora que coadyuvan en la actividad insecticida de Bt; además, hay que tener en cuenta que factores como el tipo de alimento utilizado y diferentes métodos de suministro de las proteínas en la dieta pueden originar respuestas diferentes en el insecto evaluado (Martínez et ál., 2003).
Con la DL50 obtenida para la cepa de referencia se evaluaron las proteínas recombinantes Cry1Aa, Cr1Ac, Cry1B y Cry1C produciendo un 100% de mortalidad en las larvas de primer instar de T. solanivora Se decidió entonces disminuir la dosis a 0,1 µg de proteína/cm2, con el fi n de determinar si existían diferencias en las mortalidades causadas por estas proteínas. A esta concentración las proteínas produjeron mortalidades de 52, 43%, 53 y 45,5% respectivamente. El análisis de varianza de los datos obtenidos indicó que no había diferencias significativas entre los tratamientos (P> 0,01 y P> 0,05); donde las mortalidades promedios de las proteínas fueron similares entre sí (P> 0,01, P>0,05) (figura 4).
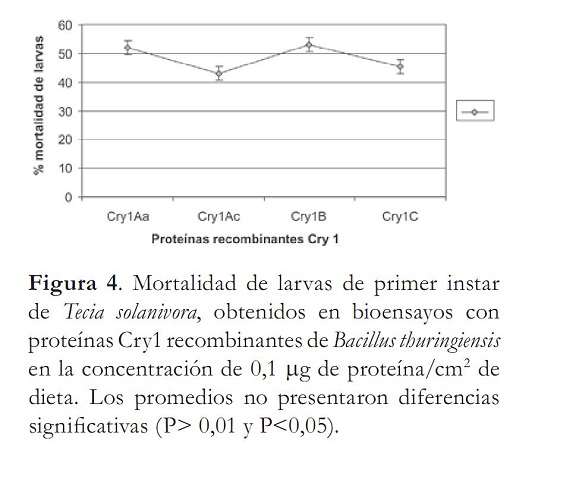
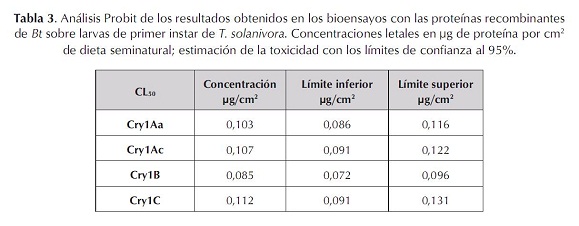
Al no existir diferencias signifi cativas entre las proteínas se calculó la concentración letal media de las cuatro toxinas, evaluando seis diluciones sucesivas de las proteínas que corresponden a: 0,05; 1; 1,5; 2; 2,5 y 3,0 µg/ cm2 de proteína. Se encontró que en este rango las CL50 obtenidas con el programa estadístico Probit Polo PC (fi nney, 1971), fueron: Cry1Aa, 0,103; Cry1Ac, 0,107; Cry1B, 0,085 y Cry1C, 0,112 µg/cm2 de proteína (tabla 3). Al someter estos resultados a un análisis de varianza y contraste ortogonal, comparando las Cl50 con el mismo valor obtenido de la cepa referencia, se determinó que no hay diferencias signifi cativas entre las proteínas recombinantes (P> 0,01), ya que las mortalidades promedio de estas toxinas Cry1Aa, Cry1Ac, Cry1B y Cry1C fueron similares (α = 0,05) (tablas 4 y 5).


En todas las repeticiones de los bioensayos la mortalidad de las larvas utilizadas como testigos fue nula, lo cual cumplió con los requerimientos necesarios para que el bioensayo sea considerado como válido según Dulmage et ál. (1971); por otro lado, los efectos producidos sobre las larvas en lo que se refi ere a disminución de tamaño, movilidad y vigor, se presentaron en forma similar con todas las proteínas evaluadas. Las concentraciones que produjeron los menores porcentajes de mortalidad permitieron que las larvas sobrevivientes se desarrollaran casi normalmente, en tanto que el crecimiento de las larvas sobrevivientes a los tratamientos con las concentraciones más tóxicas estuvo restringido.
Estos resultados demostraron que T. solanivora es altamente susceptible a las toxinas codifi cadas por los genes Cry1Aa, Cry1Ac, Cry1B y Cry1C (figura 2) Pérez et ál. (1997) propusieron que las proteínas Cry1Aa, Cry1Ab y Cry1Ac de la cepa Bt kurstakí HD1, y las proteínas Cry1Ab, Cry1B y Cry1D de una cepa nativa del banco de cepas de Corpoica (Tibaitatá, Mosquera, Cundinamarca) eran las responsables de la actividad en dicha cepas; sin embargo, estas cepas poseían múltiples genes y no pudieron determinar exactamente cuál o cuáles de estas proteínas eran las responsables de la actividad tóxica en larvas de primer instar de T. solanivora
Escriche et al. (1994) encontraron que el gen cry más específi co para Phthorimea opercullella plaga, similar por sus características a T. solanivora fue el cry1B, seguido por el cry1Aa y por el cry1C. T. solanivora parece ser susceptible a las toxinas codifi cadas por estos tres genes y además a la toxina codifi cada por el gen cry1Ac.
Igualmente, en un trabajo reciente con cepas recombinantes de Escherichia coli, Martínez et ál. (2003) analizaron la actividad tóxica de las proteínas Cry1Aa, Cry1Ab, Cry1Ac, Cry1B, Cry1C, Cry1D y Cry1E de Bt en larvas de primer instar de T. solanivora, los resultados obtenidos indicaron una toxicidad mayor de la proteína Cry1Ac respecto a las demás proteínas evaluadas. Se obtuvo un porcentaje de mortalidad promedio cercano al 20% con esta proteína a una concentración de 4,37 µg/cm2. Comparando este resultado con el obtenido en el presente estudio (CL50 = 0,107 µg/cm2) se puede suponer que la diferencia radica en que este trabajo se realizó con proteínas purificadas y clivadas. Es posible que este estado le confiera a las proteínas recombinantes una potencia tóxica mayor, ya que el proceso de proteólisis de la protoxina ya ha sido realizado favoreciendo la unión específica con el receptor (De Maadg et ál., 1999). Este efecto de las toxinas activadas tiene una influencia en la toxicidad final sobre las larvas, ya que el efecto tóxico es más rápido, produciendo el daño histopatológico característico en las células columnares del epitelio intestinal del insecto (Herrero et ál., 2001; Liu et ál., 2000).
Estos resultados son alentadores, ya que aunque se han iniciado programas de ingeniería genética en Colombia para el control de Tecia solanivora, los primeros bioensayos mostraron una mortalidad entre el 83 y 100%, y una expresión del trasgen Cry1Ac de 0,02 a 17 µg/g en tejido de tubérculo fresco (Valderrama et ál., 2007). En este estudio se evidencia la posibilidad de obtener plantas transgénicas de papa que expresen entre dos o tres genes de B. thuringiensis, y de esta forma reducir o prolongar la aparición de resistencia a este insecto plaga.
La eficiencia de la solubilización es importante para la potencia de las delta-endotoxinas de Bt, algunas inclusiones son más tóxicas para el insecto cuando se solubiliza in vitro antes de la aplicación en el ensayo, presumiblemente porque el cristal puede no disolverse realmente en el intestino medio del insecto (Zhong et ál., 2000).
El mecanismo de acción de las delta-endotoxinas establece que después del procesamiento proteolítico, la toxina se une a receptores encontrados en las microvellosidades de las células del epitelio intestinal. La interacción de las toxinas Cry con sus receptores es sin duda el paso más importante en la determinación de la especificidad de las toxinas ya que hay una correlación estrecha entre unión a receptores y toxicidad. Karim y Dean (2000), examinaron las propiedades de unión de Cry1Aa, Cry1Ab, Cry1Ac y Cry1B en varias especies de lepidópteros plaga, encontrando que en todos los insectos evaluados estas toxinas mostraron el mismo sitio de unión al receptor de membrana, lo que indica que una concentración alta de toxina puede saturar más rápidamente el sitio de unión, indicando que estas toxinas no pueden ser usadas en combinación, o que en combinación no se presenta sinergismo. Lo anterior puede explicar el aumento del valor de la CL50 en la cepa de referencia Bt HD1 al obtenido con las delta-endotoxinas recombinantes individuales, ya que esta cepa produce 5 toxinas Cry1.
El estudio realizado demostró que las larvas de primer instar de T. solanivora son sus¬ceptibles a las toxinas producidas por la cepa de referencia Bt var kurstakí HD1, y las proteí¬nas recombinantes Cry1Aa, Cry1Ac, Cry1B y Cry1C de Bacillus thuringiensis, son promisorias por su elevada actividad insecticida para el control biológico de este insecto. La actividad insecticida de las proteínas recombinantes eva¬luadas no arrojó diferencias estadísticas entre ellas, indicando que cualquiera de estas toxinas puede ser utilizada en la producción de bio¬plaguicidas más efectivos, o para la ingeniería genética de la resistencia a esta plaga en papas transgénicas.
Agradecimientos
Los autores agradecen al doctor Ruud de Maagd, Plant Research International P.O. Box 16 6700 AA Wageningen, Holanda, por proveer las toxinas recombinantes.
Referencias bibliográficas
1 Valderrama, A. M.; Velásquez, N.; Rodríguez, E.; Zapata, A.; Zaidi, M. A.; Altosaar, I.; Arango, R. 2007. Resistance to Tecia solanivora Lepidoptera: Gelechiidae) in Three Transgenic Andean Varieties of Potato Expressing Bacillus thuringiensis Cry1Ac Protein. J Econ Entomol 100 (1), 172 - 179. [ Links ]
2 Bradford, MM. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254. [ Links ]
3 Centro virtual de investigación de la cadena agroalimentaria de la papa (Cevipapa) (2004). Disponible en http:// www.cevipapa.org.co [ Links ]
4 Ciba-Geigy. 1978. Manual de ensayos de campo. Boletín cómo realizar un bioensayo. Bogotá. [ Links ]
5 De Maagd, R. A.; Kwa, M. S.-G.; Van der Klei, H.; Yamamoto, T.; Schipper, B.; Lak, J. M.; Stiekema, W. J.; Bosch, D. 1999. Domain III substitution in Bacillus thuringiensis delta-endotoxin CryIA(b) results in superior toxicity for Spodoptera exigua and altered membrane protein recognition. Applied and Environmental Microbiology 62, 1537-1543. [ Links ]
6 Dulmage, H. T.; Boening, O. P.; Rehnborg, C. S.; Hansen, G. D. 1971. Proposed standardized bioassay for formulations of Bacillus thuringiensis based on the International Unit. J Invert Pathol 18, 240-245 C. Reviews. Biotechenology 6, 87-127. [ Links ]
7 Estrada, N. 1997. Resistencia genética como una alternativa de manejo de la polilla de la papa. Resúmenes. Seminario experiencias y avances en el manejo de la polilla guatelmateca de la papa. Bogotá. [ Links ]
8 Escriche, B. 1994. Ocurrence of three different binding sites for Bacillus thuringiensis d-endotoxin in the midgut brush border membrane of the potato tuber moth, Phthorimaea operculella (Zeller). Archives of Insect biochemistry and physiology 23, 315-327. [ Links ]
9 EPPO. 2001. European and Mediterranean Plant Protection Organization, Tecia solanivora. Disponible en http://www.eppo.org/QUARANTINE/Alert List/Inoects/scrsso.html [ Links ]
10 Espinal, F.F.; Martínez, H.; Pinzón, N.; Barrios, C. 2006. La cadena de la papa en Colombia una mirada global de su estructura y dinámica 1991-2005. Ministerio de Agricultura y Desarrollo Rural. Observatorio Agrocadenas Colombia. Documento de trabajo No. 100. Disponible en http://www.agrocadenas.gov.co [ Links ]
11 Ferré, J.; Van Rie, J. 2002. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu Rev Entomol 47, 501-33 [ Links ]
12 Lopez-Ávila, A. 1996. Insectos plaga del cultivo de la papa en Colombia y su manejo. En: Papas colombianas con el mejor en-torno ambiental. Fedepapa. pp. 146-148. [ Links ]
13 Navas, L. 1985. Ciclo biológico del Chelonus phthorimaea parásito de la polilla de la papa (Scrobipalpopsis solanivora) Povolny y (Phthorimaea operculell Zeller). Memorias Congreso de Manejo Integrado de Plagas 4. Guatemala. pp. 466-474. [ Links ]
14 Finney, D. J. 1971. Probit analysis. 3 edición. Cambridge: Cambridge University Press. pp. 20-63. [ Links ]
15 Herrero, S.; Oppert, B.; Ferré, J. 2001. Diferent mechanism of resistence to Bacillus thuringiensis toxins in the indianmeal moth. Applied and Environmental Microbiology 67, 1085-1089. [ Links ]
16 Karim, S.; Dean, D. H. 2000. Toxicity and receptor binding propierties of Bacillus thuringiensis d-endotoxins to the midgut brush border membrane vesicles of the rice leaf folders, Cnaphalocrocis medinalis and Marasmia patnali. Current Microbiology 41, 276-283. [ Links ]
17 Içgen, Y. B.; Içgen, G.; Ozcengiz. 2000. Regulation of cristal protein biosintesis by Bacillus thuringiensis: II. Effects of carbon and nitrogen sources. Resource Microbiology 153, 605-609. [ Links ]
18 Lecadet, M.-M.; Sanchis, V.; Menou, G.; Rabot," P.; Lereclus, D.; Chaufaux, J.; Martouret, D. 1988. Identification of a 8-Endotoxin Gene Product Specifically Active against Spodoptera littoralis Bdv. among Proteolysed Fractions of the Insecticidal Crystals of Bacillus thuringiensis subsp. aizawai 7.29. Appl Environ Microbiol 54 (11), 2689-2698. [ Links ]
19 MacLeod, A. 2005. Data sheets on quarantine pests: Tecia solanivora. EPPO Bull 35, 399 - 401. [ Links ]
20 Martínez, O. W.; Uribe, V. D.; Ceron, S. J. 2003. Efecto tóxico de proteínas Cry de Bacillus thuringiensis sobre larvas de Tecia solanivora. Revista Colombiana de Entomología 29, 89-93. [ Links ]
21 Niño, L. 2004. Revisión sobre la polilla de la papa Tecia solanivora en Centro y Suramérica. Suplemento Revista Latinoamericana de la Papa. [ Links ]
22 Navon, A. 2000. Bioassays of Bacillus thuringiensis products used against agricultural pests bioassays of entomopathogenic Microbes and Nematodes. Wallingford, UK: CABI Publishing. pp. 1-24. [ Links ]
23 Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685. Reviews 62, 775-806. [ Links ]
24 López-Ávila, A. 2000. Insectos plagas del cultivo de la papa en Colombia y su manejo. Papas Colombianas. pp. 146-223. [ Links ]
25 Liu, Y. B.; Tabashnik, B. E.; Moar, W. J.; Smith, R. A. 2000. Synergism between Bacillus thuringiensis spores and toxins against resistant and susceptible diamondback moths (Plutella xylostella). Applied and Environmental Microbiology 64, 1385-1389. [ Links ]
26 Pérez, O.; Rodríguez, A.; Cotes, A. 1997. Método rápido y confiable para evaluar la actividad insecticida de cepas nativas de Bacillus thuringiensis contra Tecia solanivora Povolny (Lepidoptera: Gelechiidae). Revista Colombiana de Entomología 23, 113-118. [ Links ]
27 Pollet, A. 2001. Guatemalan moth Tecia solanivora devastating potato crops in Ecuador. International Pest Control 43, 75-76 [ Links ]
28 Pollet, A.; Barragán, A.; Lagnaoui, A.; Prado, M.; Onore, G.; Aveiga, I.; Lery, X.; Zeddam, J. L. 2003. Predicción de daños de la polilla guatemalteca Tecia solanivora (Povolny) 1973 (Lepidoptera: Gelechiidae) en el Ecuador. [ Links ]
29 Samir, N.; Weemen-Hendriks, M.; Dukiandjiev, S.; De Maagd, R. A. 2001. Bacillus thuringiensis delta -endotoxin Cry1 hybrid proteins with increased activity against the Colorado Potato Beetle. Applied and Environmental Microbiology 67 (11), 5328-5330. [ Links ]
30 Sánchez, G. 2000. La polilla guatelmateca de la papa en Colombia. Boletín de la papa. Corpoica 11 (4), 239-243. [ Links ]
31 Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D. R.; Dean, D. H. 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology Molecular Biology Review 62, 775-806. [ Links ]
32 Shelton A. M.; Zhao, J. Z.; Roush, R. T. 2002. Economic, ecological, food safety, and social consequences of the development of Bt transgenic plants. Annual Review Entomology 47, 845-881. [ Links ]
33 Torres, F.F.; Notz, A.; Valencia, L. 1997. Ciclo de vida y otros aspectos generales de la biología de la polilla de la papa Tecia solanivora en el estado de Táchira, Venezuela. Biology Entomology Venezuela N.S. 12 (1), 81-94. [ Links ]
34 Povolny, D. 1973. Scrobipalpopsis solanivora. A new pest of potato (Solanum tuberosum) from Central América. Acta Universitatis Agriculture, Facultas Agronómica 21, 133146. [ Links ]
35 Prias, J. 1998. Caracterización de aislamientos nativos de Bacillus thuringiensis para el control biológico de Tecia solanivora (Povolny) insecto plaga de la papa. Tesis de grado Universidad Pedagógica Nacional, Bogotá [ Links ]
36 Rodríguez, A. 1996. Consideraciones al manejo de plagas y enfermedades de la papa en Colombia. En: Papas colombianas con el mejor entorno ambiental. Bogotá D. C.: Fedepapa. pp. 122-126 [ Links ]
37 Zhong, C.; Ellar, D.; Bishop, A.; Jhonson, C.; Lin, S.; Hart, E. 2000. Characterization of Bacillus thuringiensis delta-endotoxin which is toxic to insect in three orders. Journal of Inverthebrate Pathology 76, 131-139. [ Links ]

