Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.11 n.2 Bogotá jul./dez. 2009
ARTÍCULO DE INVESTIGACIÓN
Caracterización de accesiones de papaya (Carica papaya L.) a través de marcadores AFLP en Cuba
Characterising Cuban papaya accessions (Carica papaya L.) by AFLP markers
Maruchi Alonso Esquivel1 , Martín Bautista Alor2, Matilde Ortiz García2, Adriana Quiroz Moreno2, Wolfgang Rohde 3, Lorenzo Felipe Sánchez Teyer2
1Instituto de Investigaciones en Fruticultura Tropical, Ave. 7ma. No. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana, Cuba, C.P. 11300. mejoramiento@iift.cu2Centro de Investigaciones Científicas de Yucatán, Mérida, México. santey@cicy.mx
3Max-Planck Institut für Züchtungsorschung (MPIZ), Carl-von-Linné-Weg 10, 50829, Köln, Germany. rohde@mpiz-koeln.mpg.de
Recibido: junio 13 de 2009 Aprobado: noviembre 3 de 2009
Resumen
Los marcadores moleculares son herramientas valiosas en los estudios genéticos en plantas, y están siendo empleados exitosamente en programas de mejoramiento principalmente en la elección de progenitores y en la selección. El polimorfismo observado mediante la técnica molecular AFLP (Amplified Fragment Length Polymorphism) ha sido de utilidad para estudios de diversidad genética en frutales. En el presente trabajo se realizó la caracterización molecular de 12 accesiones de papaya (Carica papaya L.) del banco de germoplasma del Instituto de Investigaciones en Fruticultura Tropical (IIFT), empleando la técnica AFLP. Se evaluaron seis combinaciones de iniciadores para la amplificación selectiva, las cuales amplificaron un total de 431 bandas con 73,3% de polimorfismo. El número total de patrones de bandas identificados fue igual en todas las combinaciones utilizadas, con un porcentaje de identificación alto, lo que sugiere que dichas combinaciones pudieran ser empleadas para estudios de variabilidad genética en papaya. En general, los resultados presentados demuestran que existe diversidad genética entre las accesiones evaluadas, lo cual constituye un reflejo del origen que presentan los genotipos analizados a partir de la introducción de materiales foráneos y la polinización abierta de un grupo de materiales selectos. Por tanto, se recomienda retomar la prospección y selección de accesiones locales, así como la introducción de nuevos genotipos foráneos, como dos vías fundamentales para aumentar la diversidad genética presente en el banco de germoplasma de papaya de Cuba.
Palabras clave: marcadores moleculares, polimorfismo, diversidad genética.
Abstract
Molecular markers are valuable tools for genetic studies in plants and they are often used successfully in genetic breeding, mainly for choosing progenitors and selection. Polymorphism observed by amplified fragment length polymorphism (AFLP) has been useful for genetic diversity studies in fruit trees. Twelve papaya accessions from the Tropical Fruit Crop Research Institute germplasm bank were molecularly characterised by AFLP. 431 bands having 73.3% polymorphism were obtained using 6 primer combinations. The total number of band patterns identified was the same in all combinations assayed with a high percentage of identification, suggesting that such primer combinations could be used for genetic variability studies in papaya. The results demonstrated genetic diversity among the papaya accessions evaluated, indicating the origin of the analysed genotypes from exogenous material and open pollination of a selected group of material. It is thus recommended that local accessions and their selection be monitored as well as the introduction of new foreign genotypes as two ways of increasing the genetic diversity of the Cuban papaya germplasm bank.
Keywords: molecular marker, polymorphism, genetic diversity.
Introducción
La papaya es una planta herbácea, de crecimiento rápido y de vida corta. Es nativa de América Tropical y ampliamente cultivada en todas las regiones tropicales por sus frutos comestibles y alto contenido de vitaminas (Storey, 1969).
La especie Carica papaya L. pertenece a una pequeña familia de dicotiledóneas: Caricaceae, la cual consta de seis géneros y 35 especies. Entre ellos se encuentra el género Carica, que incluye esta sola especie y resulta la más importante desde el punto de vista económico (Badillo, 2000).
El Caribe constituye una zona de diversificación secundaria de la papaya, donde las poblaciones locales presentan adaptaciones a las condiciones regionales. La conservación y evaluación de la papaya o fruta bomba constituye un paso importante para el desarrollo sostenible del cultivo.
La introducción, caracterización y evaluación de accesiones de papaya pueden permitir la identificación de nuevos genotipos, con vista a proporcionar el material básico para programas de mejoramiento genético de este frutal (Alonso et al., 2007). Estos programas van encaminados hacia el logro de mejores variedades desde el punto de vista morfoagronómico y tolerancia a las virosis, por ser este último el problema patológico de mayor importancia en la economía del mismo (Paris, 2005).
Existe gran variedad de métodos para clasificar las accesiones, los niveles de diversidad y las relaciones entre los individuos (Marcucci et al., 2001). Los primeros y los de más antigua utilización son los basados en la morfología de la planta, los cuales permiten una primera observación, a simple vista, de sus atributos principales para analizar las diferencias entre las accesiones en estudio. Estos métodos son relativamente económicos y constituyen la base de la caracterización de las muestras presentes en los bancos de germoplasma (Rallo et al., 2002).
Los métodos citogenéticos y moleculares (bioquímicos y basados en el ADN) permiten analizar las diferencias entre los cromosomas, las proteínas o el ADN de las plantas (Avise, 1994), y se utilizan para complementar los estudios donde los marcadores morfológicos han proporcionado resultados limitados; así como para acelerar los programas de mejoramiento (Persson, 2001).
La genética basada en los marcadores moleculares está siendo integrada en varios programas de investigación y juega un papel importante en el mejoramiento de plantas (Oliveira et al., 2008). Los marcadores moleculares tienen un rango amplio de aplicaciones, entre éstas: el mapeo y marcaje de genes, los estudios de diversidad genética, las relaciones entre y dentro de las diferentes especies, y la distribución de esa diversidad entre regiones geográficas, entre otras (Valdés-Infante et al., 2008).
Las técnicas de Polimorfismo de Longitud de Fragmentos Restringidos (RFLP) (Aradhya et al., 1999), Polimorfismo del ADN Amplificado al Azar (RADP) (Stiles, 1993; Jobin-Décor et al., 1997; Cattaneo, 2001), y Polimorfismo de Longitud de Fragmentos Amplificados (AFLP) (Van Droogenbroeck et al., 2002; Kim et al., 2002; Morales et al., 2004), han sido aplicadas en papaya mostrando ser adecuadas para la determinación de relaciones genéticas y estudios de diversidad. Por otra parte, el empleo de los microsátelites (SSR) se considera apropiado para detectar y cuantificar la variabilidad genética en dicha especie (Pérez et al., 2006), y la determinación de sexo en plantas de papaya (Ma et al., 2004; Oliveira et al., 2007; Sánchez y Núñez, 2008). Estas técnicas se utilizan para complementar los estudios donde los marcadores morfológicos han proporcionado resultados limitados, o para acelerar los programas de mejoramiento (Meerow et al., 2003).
En este sentido, AFLP es reconocida como una técnica molecular confiable y eficiente comparada con los marcadores RFLP, RADP o microsatélites (Vos et al., 1995; Powell et al., 1996; Pejic et al., 1998; Kim et al., 2002) y su ventaja principal es el gran número de bandas polimórficas que genera (Rafalski et al., 1996).
El objetivo del presente trabajo fue estimar la variabilidad genética entre accesiones de papaya pertenecientes al banco de germoplasma de este frutal en Cuba, mediante el polimorfismo generado por los AFLP.
Materiales y métodos
Material vegetal
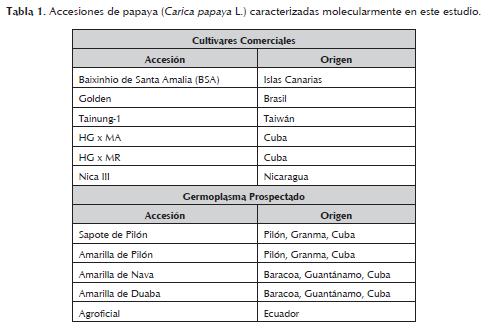
Se utilizaron doce accesiones de papayas seleccionadas al azar y pertenecientes al banco de germoplasma del IIFT en Cuba. Las accesiones corresponden a cultivares comerciales, materiales introducidos de diferentes orígenes: Brasil, Islas Canarias (Cultivares del Grupo Solo), Nicaragua, Ecuador, Costa Rica, Taiwán (cultivares del Grupo Formosa), México, y germoplasma prospectado en Cuba (tabla 1).
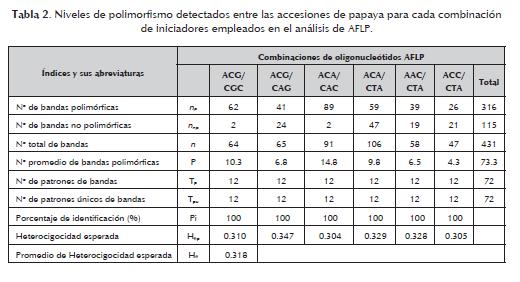
2. Número de bandas no polimórficas (nnp). 3. Número total de bandas (n = np+ nnp).
4. Número promedio de bandas polimórficas por unidad de ensayo (P= np /u).
5. Número de patrones de bandas identificados por combinación de oligonucleótidos (Tp).
6. Número de patrones únicos de bandas identificados por combinación de oligonucleótidos (Tpu).
7. Porcentaje de identificación (Pi=( Tpu / Tp)x100).
8. Heterocigocidad esperada (Hep) del loci polimórfico: He = 1 - Εpi2 donde pi es la frecuencia de alelos en el imo alelo y la media aritmética de la heterocigosidad del loci polimórfico: 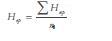 , donde n es el número de marcadores analizados. En todos los casos, los valores fueron corregidos por el tamaño de la muestra como:
, donde n es el número de marcadores analizados. En todos los casos, los valores fueron corregidos por el tamaño de la muestra como: 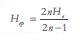 (12) Media de la heterocigocidad esperada de los loci polimórficos: He= (ΕHep)/ u.
(12) Media de la heterocigocidad esperada de los loci polimórficos: He= (ΕHep)/ u.
Se estimó la similitud genética entre todos los genotipos estudiados, para ello se utilizó el subprograma SIMQUAL, empleando como coeficiente Dice (Dice, 1945) y como método de agrupamiento, el Método de las Medias Aritméticas por Grupo no Ponderadas (UPGMA) por presentar los valores cofenéticos más elevados. La conformación de los grupos en los análisis de conglomerados, y la determinación de los posibles grupos de diversidad se apoyó mediante el método de remuestreo utilizando el programa Winboot (1998). Los cálculos fueron realizados mediante el paquete estadístico NTSys-PC (versión 2.1).
Resultados y discusión
En el estudio de las accesiones de papaya con las seis combinaciones de iniciadores se obtuvo un total 431 bandas con un 73,3% de polimorfismo. Esto corrobora la eficiencia de los AFLP para generar un gran número de marcadores por corrida como lo refiere Valdés-Infante (2009). La tabla 2 muestra los resultados relacionados con el polimorfismo para cada combinación.
El número de bandas polimórficas varió de 26 a 89 con las combinaciones E-ACC/ MCTA y E-ACA/MCAC, respectivamente. El porcentaje de polimorfismo por combinación varió de 48,7 a 97,8%. Dichos resultados fueron superiores a los señalados por Kim et al. (2002), quienes utilizando nueve combinaciones obtuvieron 186 bandas polimórficas (42%) en un total de 445 bandas detectadas en un estudio de diversidad genética en 63 accesiones de Carica papaya L. El número de patrones de bandas totales fue de 72, y el número de patrones únicos fue de 12. Estos niveles de polimorfismo fueron menores a los obtenidos en un estudio similar realizado en guayabo (Psidium guajava L.), donde se registraron valores de 150 y 45, respectivamente (Valdés-Infante et al., 2008). El número total de patrones de bandas identificados se mantuvo igual en todas las combinaciones utilizadas, con un porcentaje de identificación alto por combinación (Pi=100%). Esto sugiere que estas seis combinaciones pudieran ser empleadas para estudios de variabilidad genética en accesiones de papaya. En el caso del número de patrones de banda únicos (Tpu) mostró el mismo valor en las seis combinaciones, 12 por unidad de ensayo, lo que comparado con el total de patrones de bandas detectado sugiere la efectividad de estos oligonucleótidos en la identificación de todos los genotipos evaluados (tabla 2). Esto explica la potencialidad de los AFLP para estudios de identificación en este frutal; resultados similares han sido obtenidos en aguacatero (Persea americana Mill.) por Rodríguez et al. (2008), en mango (Mangifera indica L.) por Capote (2007) y en el guayabo (Psidium guajava L.) por Valdés- Infante et al. (2008). De manera general, la heterocigocidad promedio esperada fue 0,31, resultados que corroboran los valores obtenidos para la heterocigocidad esperada por combinación. La combinación E-ACG/M-CAG mostró el mayor valor de heterocigocidad esperada (He = 0,34), mientras que con las combinaciones E-ACA/ M-CAC y E-ACC/ M-CTA se obtuvo el menor valor (He = 0,30). El coeficiente de similitud de Dice varió de 0,299 a 0,785, con un promedio global de 0,542. Este valor promedio fue superior (0,880) en el estudio conducido por Kim et al. (2002), como era de esperarse por el mayor número de combinaciones de iniciadores (9) y accesiones de papaya analizadas (63). Por otra parte, los valores de similitud obtenidos muestran a Nica-III como la accesión con mayor heterogeneidad genética al presentar el valor más bajo (0,299). En la figura 1 se muestran las relaciones genéticas entre las 12 accesiones de papaya evaluadas, se puede apreciar que el marcador AFLP permitió la discriminación de todas las accesiones. El dendrograma presenta tres grupos de diversidad bien definidos y dos accesiones externas (figura 1). El Grupo I (incluye accesiones introducidas del Grupo Solo Sunset, Golden y el híbrido Tainung-01); el Grupo II (incluye una accesión local y dos cultivares comerciales cubanos HGxMR y HGxMA) y el Grupo III (incluye tres accesiones locales y una accesión introducida de Ecuador - Agroficial). Dentro de las accesiones externas se encuentra el cultivar comercial Baixinhio de Santa Amalia (BSA) y el cultivar introducido de Nicaragua (Nica-III). El cultivar BSA constituye una accesión externa en el dendrograma generado utilizando los datos AFLP, lo que puede deberse a que fue originada a partir de una mutación natural del cv Sunrise Solo en la localidad de Espirito Santo, Brasil (Dantas et al., 2002). Se observa la asociación de Golden y Sunset (figura 1, Grupo I), aspecto que potencia la afirmación de que son cultivares interrelacionados y pertenecientes a la línea Solo. El cultivar Sunset constituye una línea hermana del cv Sunrise, originada a partir de un mismo parental y, a su vez, la posición del cultivar Tainung-01 entre las accesiones de este grupo no resulta sorprendente, si se tiene en cuenta que este híbrido fue obtenido a partir del cruce de una selección de papaya de Costa Rica y el cv Sunrise. Las accesiones HGxMR y HGxMA pertenecientes al Grupo II corroboran sus relaciones de parentesco, ambos genotipos fueron obtenidos a partir del cruzamiento con el cultivar Hortus Gold y Maradol. Además, la ubicación en este grupo de la accesión local Sapote de Pilón, seleccionada por un productor en la región de Pilón (provincia Granma), mostró una similitud genética de 0,59, lo que pudiera indicar que este material está compartiendo parte de su genoma a través de los cruzamientos por polinización abierta con el mismo progenitor común de ambos cultivares, teniendo en consideración que el progenitor Maradol constituye el cultivar más extendido comercialmente en el país. Las accesiones locales Amarilla de Nava, Amarilla de Duaba y Amarilla de Pilón, la mayoría colectadas a partir de diferentes productores de papaya en la región oriental de Cuba formaron el Grupo III, sugiriendo una estrecha relación genética entre estos materiales, lo que pudiera estar indicando un posible origen común en estas accesiones. En el caso específico de la accesión introducida de Ecuador, no se conoce información alguna relacionada con sus parentales, por lo cual se hace necesario desarrollar estudios adicionales con marcadores moleculares codominantes como los SSR para determinarlo. Igualmente, resultó efectiva la introducción del genotipo foráneo Nica-III con el objetivo de ampliar la base genética de la colección, el cual tiene un origen geográfico diferente al de las accesiones presentes en el banco de germoplasma, lo cual se ve reflejado en su ubicación en el dendrograma. No obstante, se requiere la introducción de nuevos materiales con un origen más diverso y características morfológicas más distintivas. De forma general, los resultados presentados demuestran que entre las accesiones de papaya existe diversidad genética, lo cual constituye un reflejo del origen que presentan los genotipos analizados a partir de la introducción de materiales foráneos y la polinización abierta de un grupo de materiales selectos. Por tanto, se recomienda seguir retomando la prospección y selección de accesiones locales, así como la introducción de nuevos genotipos foráneos, como dos vías fundamentales para aumentar la diversidad genética presente en el banco de germoplasma de papaya. Valdés Infante (2009) y Taamalli et al. (2006) arribaron a estas mismas conclusiones en la caracterización de accesiones de Psidium guajava L. y Olea europaea L., respectivamente. Los resultados obtenidos y discutidos en este trabajo coinciden y reafirman lo reportado por Rodríguez y Arencibia (2002), respecto a lo poderosa que resulta la técnica AFLP, y los marcadores moleculares en general, para la detección y evaluación de la variación genética en colecciones de germoplasma, así como en los estudios de biodiversidad. Además, también corrobora lo planteado en la literatura, de que los marcadores moleculares constituyen una herramienta moderna y poderosa para el viejo arte de la selección (Coto y Cornide, 2002). Conclusiones Las combinaciones de iniciadores AFLP utilizadas en este estudio permitieron la diferenciación de todas las accesiones evaluadas. Los resultados evidencian la utilidad de este tipo de marcadores en la identificación de genotipos, y en estudios de diversidad genética en papayo, información que puede ser de gran utilidad en el diseño de métodos y estrategias de mejoramiento genético. Las seis combinaciones evaluadas pudieran ser empleadas para estudios de variabilidad genética en accesiones de papaya, por el alto porcentaje de identificación por combinación (Pi = 100%) obtenido. La diversidad genética observada entre las accesiones de papaya sugiere que la prospección y selección de accesiones locales, así como la introducción de nuevos genotipos foráneos, constituyen dos vías fundamentales para aumentar la variabilidad en el banco de germoplasma del cultivo. Referencias bibliográficas 1 Alonso, M., Ramos, R., Tornet, Y. 2007. Caracterización y evaluación de los recursos genéticos de papaya (Carica papaya L.). CitriFrut 24 (1): 38-42. [ Links ] 2 Aradhya, M. K., Manshardt, R. M., Zee, F., Modern C. W. 1999. A phylogenetic analysis of the genus Carica L. (Caricaceae) based on restriction fragment length variation in a cpDNA intergenic spacer region. Genet Resour Crop Evol 46: 579-586. [ Links ] 3 Badillo, V. M. 2000. Carica L. vs. Vasconcella St. Hil. (Caricaceae) con la rehabilitación de este último. Ernstia 10: 74-79 [ Links ] 4 Belaj, A., Satovis, Z., Cipriano, G., Baldono, L., Testolin, R., Rallo, L., Trujillo, I. 2003. Comparative study of the discriminating capacity of RADP, ALP and SSR markers and of their effectiveness in stablishing genetic relationships in olive. Theor Appl Genet 107: 736- 744. [ Links ] 5 Capote, M. 2007. Caracterización morfoagronómica y molecular de variedades comerciales de mango (Mangifera indica L.) en Cuba. Tesis de Maestro en Ciencias. Facultad de Biología, UH. 114 p. [ Links ] 6 Cattaneo, L. F. 2001. Evaluación de la diversidad genética y análisis de generaciones en papaya (Carica papaya L). Tesis de Doctorado en Producción Vegetal, Universidad Estadual del Norte Fluminense-UENF. Campos dos Goytacazes, RJ. 95 pp. [ Links ] 7 Coto, O., Cornide, M. T. 2002. Principales aplicaciones de los marcadores moleculares. En: Marcadores Moleculares. Nuevos horizontes en la genética y la selección de plantas. La Habana: Ed. Félix Varela. pp. 92-110. [ Links ] 8 Dice, L. R. 1945. Measures of the amount of ecological association between species. Ecology 26: 297-302. [ Links ] 9 Doyle, J. J., Doyle, J. L. 1990. Isolation of plant DNA from fresh tissue. Focus 12: 13-5. [ Links ] 10 Jobin-Décor, M. P., Graham, G. C., Henry, R. J., Drew, R. A. 1997. RADP and isozyme analysis of genetic relationship between Carica papaya and wild relatives. Genet Resour Crop Evol 44: 471-477. [ Links ] 11 Kim, M. S., Moore, P. H., Zee, F., Fitch, M. M. M., Steiger, D. L., Manshardt, R. M., Paull, R. E., Drew, R. A., Sekioka, T., Ming, R. 2002. Genetic diversity of Carica papaya as revealed by AFLP markers. Genome 45: 503-512. [ Links ] 12 Ma, H., Moore, P. H., Liu, Z., Kim, M. S., Yu, Q., Fitch, M. M. M., Sekioka, T., Paterson, A. H., Ming, R. 2004. High density linkage mapping revealed suppression of recombination at the sex determination locus in papaya. Genetics 166: 419-436. [ Links ] 13 Meerow, A. W., Wisser, R. J., Brown, J. S., Kuhn, D. N., Schnell, R. J. Broschat, K. 2003. Analysis of genetic diversity and population structure within Florida coconut (Cocos nucifera L.) germplasm using microsatellite DNA, with special emphasis on the Fiji dwarf cultivar. Theor Appl Genet 106: 715-726. [ Links ] 14 Morales, A. R., Medina, D. L., Yaguache, B. D. 2004. Genetic diversity, phylogeny and geographic distribution of the genus Vasconcella in Southern Ecuador. Lyonia 7 (2): 15-27. [ Links ] 15 Oliveira, E. J., Dantas, J. L. L., Castellen, M. da S., Lima, D. S., Barbosa, H. S. e Motta, T. B. N. 2007. Marcadores moleculares na predição do sexo em plantas de mamoeiro. Pesq Agropec Bras 42 (12): 1747-1754. [ Links ] 16 Oliveira, E. J., Dantas, J. L. L., Castellen, M. da S., Machado, A. D. 2008. Identificação de microssatélites para o mamoeiro por meio da exploração do banco de dados de DNA. Rev Bras Frutic 30 (3): 841-845. [ Links ] 17 Paris. 2005. Consensus document on the biology of papaya (Carica papaya). OECD Environment, Health and Safety Publications. Series on Harmonisation of Regulatory Oversight in Biotechnology, 33. 65p. [ Links ] 18 Pejic, J., Ajmone-Marsan, P., Kozumplik, V., Castiglioni, P., Tamarino, G., Motto, M. 1998. Comparative analysis of genetic similarity among mayze inbred lines detected by RFLPs, RADPs, SSRs and AFLPs. Theor Appl Genet 97: 1248-1255. [ Links ] 19 Pérez, J. O., Dambier, D., Ollitrault, P., D Eeckenbrugge, G. C., Brottier, P., Froelicher, S., Risterucci, A-M. 2006. Microsatellitte markers in Carica papaya L.: isolation, characterization and transferability to Vasconcellea species. Molecular Ecology Notes 6: 212-217. [ Links ] 20 Powell, W., Morgante, M., Andre, C., Hanafey, M., Vogel, J., Tingey, S., Rafalski, A. 1996. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol Breed 2: 225-238. [ Links ] 21 Rafalski, J. A., Vogel, J. M., Morgante, M., Powell, W., André, C., Tingey, S. V. 1996. Generating and using DNA markers in plants. In: B, Birren; E. Lai (eds). Non Mammalian Genomic Analysis: A Practical Guide. London: Academic Press. pp.75-134. [ Links ] 22 Rodríguez, M., Arencibia, A. (2002). Principales tipos de marcadores del polimorfismo de los ácidos nucleicos. Técnicas Analíticas. En: Marcadores Moleculares. Nuevos horizontes en la genética y la selección de las plantas. La Habana: Editorial Félix Varela. pp. 13-31. [ Links ] 23 Rodríguez, N. N., Fuentes, J. L., Coto, O., Fuentes, V. R., Ramírez, I. M., Becker, D., Rodríguez, I., Rohde, W. 2008. Comparison of morphological traits, isozymes and DNA markers for estimating the discriminating capacity and informativeness in cuban avocado (Persea americana Mill.) accessions. Revista CENIC Ciencias Biológicas 39 (3): 2008. [ Links ] 24 Rohde, W., Kullaya, A., Rodríguez, A., Ritter E. (1995). Genetic analysis of Cocos nucífera L. by PCR amplification of spacer sequences separating a subset of copialike Eco RI repetitive elements. J Genet & Breed 49: 179-186. [ Links ] 25 Sánchez, E., Núñez, V. M. 2008. Evaluación de marcadores moleculares tipo SCAR para determinar sexo en plantas de papaya (Carica papaya L.). Revista Corpoica – Ciencia y Tecnología Agropecuaria 9 (2): 31-36. [ Links ] 26 Sneath, P. H. A. and Sokal R. R. 1973. Numerical Taxonomy. San Francisco: Freeman. 573 pp. [ Links ] 27 Stiles, J. I., Lemme, C., Sondur, S., Morshidi, M. B., Manshardt, R. M. 1993. Using randomly amplified polymorphic DNA for evaluating genetic relationships among papaya cultivars. Theor Appl Genet 85: 697-701. [ Links ] 28 Storey, W. B. 1969. Pistillate papaya flower: A morphological anomaly. Science 163: 401-405 [ Links ] 29 Taamalli, W., Geuna, F., Banfi, R., Bassi, D., Daoud, D., Zarrouk, M. 2006. Agronomic and molecular analyses for the characterisation of accessions in Tunisian olive germplasm collections. Electronic Journal of Biotechnology. 10 p. [ Links ] 30 Valdés-Infante, J. 2009. Utilización de caracteres morfoagronómicos y de marcadores de ADN para el desarrollo de una metodología que contribuya al mejoramiento genético del guayabo (Psidium guajava L.) en Cuba. Tesis de Doctor en Ciencias Biológicas. Facultad de Biología, UH. 110 p. [ Links ] 31 Valdés-Infante, J., Rodríguez, N. N., Velásquez, B., Rivero, D., Martínez, F., Risterucci, A-M., Billotte, N., Becker, D., Rohde, W. 2008. Comparison of the polymorphism level, discriminating capacity and informativeness of morph-agronomic traits and molecular markers in guava (Psidium guajava L.). In: II Symposium International of Guava and other Myrthaceas, Mérida, México, 2008. [ Links ] 32 Van Droogenbroeck, B., Breyne, P., Goetghebeur, P., Romeijn-Peeters, E., Kind, T., Gheysen, G. 2002. AFLP analysis of genetic relationships among papaya and its wild relatives (Caricaceae) from Ecuador. TAG Theoretical and Applied Genetics 105 (2-3): 289-297. [ Links ] 33 Vos, P., Hogers, R., Blecker, M., Reijans, M., Van de Lee T., Hornes M., Fritjers, A., Pot, J., Peleman, J., Kuiper, M., Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleid Acids Research 23: 4407- 4414. [ Links ] 34 Rallo, P., Belaj, A., De La Rosa, R., Trujillo, I. 2002. Marcadores moleculares (en línea). Córdoba, España. http://www.extremadura21.com/caudal 6 p. [ Links ] 35 Avise, J. C. 1994. Molecular markers, Natural History and Evolution. New York: Chapmas y Hall. 511 p. [ Links ] 36 Persson, H. 2001. Estimating Genetic Variability in Horticultural Crop Species at different Stages of Domestication. Doctoral Thesis. 30 p. [ Links ] 37 Marcucci, S., Acuña, C., Torales, S., Zelener, N., Pathaver, P., López, G., Harrand, L., Hopp, E. 2001. Evaluación de la Variabilidad Genética en huertos semilleros de especies de Eucalyptus. http://www.inta.gov.ar/ediciones/idia/forest/genetica10.pdf 7 p. [ Links ]