










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.11 no.2 Bogotá July/Dec. 2009
ARTÍCULO DE INVESTIGACIÓN
Quantitative transient GUS expression in J-104 rice calli through manipulation of in vitro culture conditions.
Expresión transitoria GUS en callos de arroz (var. J-104) mediante la optimización de las condiciones de cultivo in vitro
Maylin Pérez Bernal 1 , Carlos Hernández 1, María Teresa Barceló 1, Magalis Delgado2 , Raúl Armas1
1Master in Plant Biology, Research Department. Center for Genetic Engineering and Biotechnology of Sancti Spiritus. P.O.Box 83, P.C. 60200. Sancti Spiritus, Cuba. maylin.perez@cigb.edu.cu (corresponding author).2Laboratory Technician.
Recibido: septiembre 15 de 2009 Aprobado: noviembre 12 de 2009
Abstract
This paper purposes suitable conditions for callus induction and co-cultivation with Agrobacterium tumefaciens of J-104 rice cultivar. It was evaluated the effect of different concentrations of 2.4-D and agar, and the inclusion of L-proline and L-glutamine in callus culture medium. The use of 2.5 mg/L 2.4-D and 0.8% agar allowed the highest percentage of embryogenic calli. Callus formation was improved considerably with 500 mg/L of L-proline and L-glutamine in the culture medium. Different factors were studied throughout co-cultivation of calli with A. tumefaciens: inoculation time, co-cultivation temperature, concentration of acetosyringone and co-cultivation period. Transient GUS expression was quantified by fluorometry in all co-cultivated calli. The best results were obtained with the following conditions: 10 min as inoculation time, 100µM acetosyringone in co-cultivation medium, temperature of 20°C, and 3 days as co-cultivation period.
Key words: Agar, callus, co-cultivation, fluorometric GUS activity.
Resumen
Se describen las condiciones óptimas para la callogénesis y cocultivo de callos con Agrobacterium tumefaciens de la variedad de arroz J-104. Se determinó el efecto de diferentes concentraciones de 2.4-D, agar y de L-prolina y L-glutamina en el medio de cultivo de callos. El uso de 2,5 mg/L de 2.4-D y 0,8% de agar permitió lograr el porcentaje más alto de callos embriogénicos. La formación de callos fue mejorada considerablemente con la adición de 500 mg/L de L-prolina e igual concentración de L-glutamina en el medio de cultivo. Se estudiaron diferentes factores en el cocultivo de los callos con A. tumefaciens: tiempo de inoculación, concentración de acetosiringona, temperatura y tiempo de cocultivo. Para comparar el efecto de cada factor sobre la expresión GUS se cuantificó la actividad transitoria mediante fluorimetría. Los valores más altos de actividad fluorimétrica fueron obtenidos con las siguientes condiciones: 10 min de inoculación, 100µM de acetosiringona en el medio de cocultivo y 3 días de cocultivo a 20 °C.
Palabras clave: agar, callo, cocultivo, actividad fluorimétrica GUS.
Introduction
Indica rice varieties are often considered to be sensitive to tissue culture and poorly responsive to genetic transformation, exhibiting culture-specific genotype differences that render them recalcitrant to transformation. Although the first reports of any rice transformation have been available since the late 1980s, the paucity of reports describing transgenic indica lines may confirm that indica varieties are difficult to transform (Lin and Zhang, 2005). It has been a constant endeavour to identify suitable explants to produce embryogenic calli, under appropriate culture condition, to maximise the callus yield.
Several studies have described the evaluation of a few indica lines for their totipotency and have demonstrated their transgenic ability (Datta et al., 2001; Lin and Zhang, 2005). Various growth regulators, amino acids and gelling agents have been assayed to improve frequency of callus formation, as a pre-requisite to select the cells carrying the transgenes, and subsequent recovery of whole plants with superior characters (Visarada and Sarma, 2002).
One of the growth regulators most commonly used to obtain embryogenic calli in cereals is 2.4-dichlorephenoxyacetic acid (2.4-D). This auxin generates DNA hypermethylation, which maintains the cells in a highly active mitotic stage and, therefore, in a proembryonic phase (Meneses et al., 2005). Gelling agent also plays an important role in callus induction. Regardless of the type of gelling agent used, increase of gel strength is associated with reduced water availability from the medium to the cultures (Klimaszewska et al., 2000).
Supplementation of amino acids in culture media has been reported to enhance somatic embryogenesis. The most frequently amino acids used in callus culture are glycine, asparagine, L-glutamine, L-proline and L-tryptophan (Kopertekh and Stribnaya, 2003; Saharan et al., 2004; Zaidi et al., 2006). Amino acids provide a source of reduced nitrogen, which is readily metabolized by plant cells, stimulating faster cell growth and development. Differential responses of organic nitrogen sources indicate the requirement of specific amino acids for specific events during in vitro morphogenesis. Therefore, the additional amino acids appear to have the potential to enhance the roles of suitable nitrogen sources (Sarker et al., 2007).
Factors involved in transformation mediated by Agrobacterium tumefaciens can be modified to transform important rice genotypes. The transfer of T-DNA and its integration into the plant genome is influenced by several A. tumefaciens and plant tissue specific factors, including plant genotype, explant, vectors-plasmid, bacterium inoculation, addition of vir-gene inducing synthetic phenolic compounds, culture media composition, tissue damage, suppression and elimination of A. tumefaciens infection after co-cultivation (Tyagi et al., 2007).
In Cuba all rice varieties are included into indica group. At present, J-104 is the most important commercial cultivar. Some biotic and abiotic factors affect its crop, with subsequent decrease of rice national production. Establishment of an efficient tissue culture system and genetic transformation procedure is necessary to enhance agronomical properties of J-104. But J-104 has been quite recalcitrant for tissue culture in respect to others Cuban varieties that have been studied before (Pérez Bernal et al., 2007a; 2008a; 2008b).
The purpose of this research was to establish appropriate conditions for callus induction and co-cultivation with Agrobacterium tumefaciens of J-104 rice variety. It was evaluated the effect of different concentrations of 2.4-D and agar, and the inclusion of L-proline and L-glutamine in callus culture medium. In addition, different factors were studied throughout co-cultivation of calli with Agrobacterium tumefaciens. Transient ß-D-Glucuronidase (GUS) expression in callus tissues was quantified by fluorometry method.
Materials and methods
Callus induction
Mature dehulled seeds, of J-104 Cuban rice variety, were surface sterilized with 70% ethanol for 1 min and rinsed with distilled water. Then, they were sterilized with 2.5% active chlorine solution, for 25 min by vigorous shaking. Treated seeds were rinsed twice with distilled water and blot to dry them onto filter paper. The surface sterilized seeds were placed on different culture media for callus induction and plant regeneration.
The N6 medium (Chu et al., 1975) was used for embryogenic callus induction, enrichment with 1g/L casein hydrolysate, 2.4-D (2, 2.5 or 3.0 mg/L) and agar (0.8; 1.0 or 1.3%). The 2.4-D treatments were combined with each one of agar concentrations, for a total of 9 treatments (table 1). Every of them consisted of 12 repetitions with 10 seeds each one.
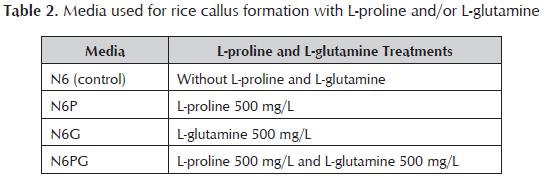
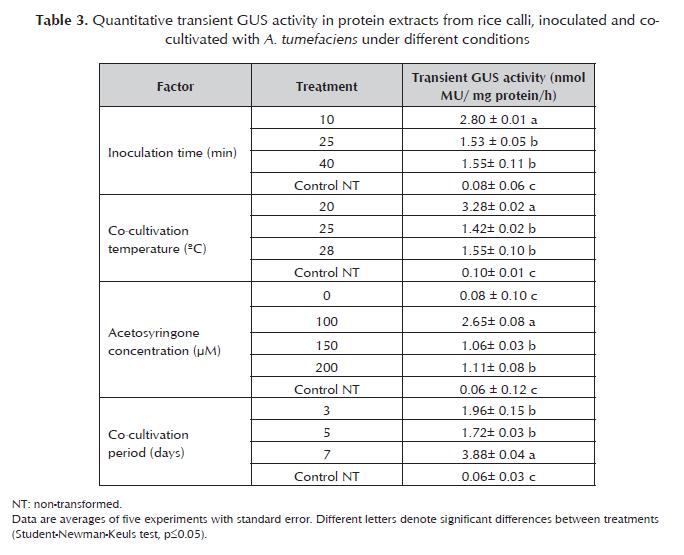
Callus induction medium, with the best combination of 2.4-D and agar, was modified by inclusion of L-proline or L-glutamine or both, in accordance with table 2. There were made 14 replications per treatment with 10 seeds each one, including a control without amino acids. The relative fresh weight growth (RFWG) of calli was determined using random samples of 100 calli pertreatment with L-proline and/or L-glutamine and control without amino acids. The pH of all media was adjusted to 5.7 before autoclaving at 120 °C for 20 min. Media were poured into Petri dishes. Cultures were maintained during 21 days, in the dark, at 28°C. In all experiments, embryogenic calli formation frequency (ECFF) (%) was calculated as follows: Bacterial strain and plasmid Agrobacterium tumefaciens strain EHA105, harbouring the binary vector pCAMBIA1301, was used to transform embryogenic rice calli. The vector contains a hygromycin-resistant gene (hpt) and uidA reporter gene, interrupted by castor bean (Ricinus communis) catalase 1 intron. Both genes are driven by the CaMV 35S promoter. A. tumefaciens strain was grown in 50 mL of AB liquid media (Hiei and Komari, 2006) with 50 mg/L kanamycin and 40 mg/L rifampicin, at 28±1 °C, in the dark, shaking at 250 rpm. Cultures were centrifuged at 3000 xg for 20 min, and Agrobacterium cells were resuspended in AAM medium (Hiei and Komari, 2006), to give optical density (600 nm) of 1.0. Acetosyringone was added at a concentration of 100 µM just prior to bacterium inoculation. Inoculation and co-cultivation of embryogenic calli with A. Tumefaciens Embryogenic calli were distributed and submerged in three flasks containing 20 mL of inoculums with optical density (600 nm) of 1.0. Inoculation was carried out during 10, 25 or 40 min, at 26 °C. The inoculum was pippeted out and infected calli were dried on sterile absorbent paper. Calli were put on co-cultivation medium (N62M), consisted of callus induction medium with 10 g/L glucose and pH 5.2. The following co-cultivation factors were assessed: Temperature. Calli were incubated during 3 days in the dark, at 20, 25 or 28 °C. Acetosyringone concentration. There were assayed 0, 100, 150 or 200 µM of acetosyringone in N62M medium. Calli were cultured on each medium during 3 days, in the dark, at 20 °C. Co-cultivation period. Calli were placed on N62M with 100 µM acetosyringone, in the dark, at 20 °C. They were incubated for 3, 5 or 7 days. Five replications, with 40 calli each one, were applied for all treatments during inoculation and co-cultivation. Non-transformed calli were included as control in all experiments. Antibiotic washes and quantitative GUS assay Following co-cultivation, calli were washed exhaustively with sterile distilled water, plus a final rinse with 500 mg/L cefotaxime during 10 min, and then were blotted on sterile absorbent paper. Calli from all co-cultivation conditions were analyzed by quantitative fluorometric GUS assay. The proteins were extracted from calli and the amount of protein was determined according to the Bradford assay with bovine serum albumin as standard. The protein concentration was quantified with a spectrophotometer at 620 nm. Protein was incubated with 1 mM MUG buffer at 37 °C according to Jeffersons protocol (Jefferson, 1987). The enzymatic reaction was measured by a fluorometer (model PR-521) with excitation at 365 nm and emission at 455 nm. The reaction was stopped each hour with Na2CO3 buffer. Fluorescence readings of the samples were utilized to generate a standard curve. Data from the curve were applied to calculate the amount of metylumbelliferone (MU) liberated by each sample. Values from the fluorescence assay were converted to moles of MU/h, and the quantitative GUS activity was expressed finally as nmol (MU)/mg protein/h. Statistical analysis Two way analysis of variance (ANOVA) was applied for embryogenic callus formation frequency. One way ANOVA was used for relative fresh weight growth of callus and for fluorometric GUS assay results. Multiple comparisons of means were applied by Student- Newman-Keuls Test (p=0.05). Analysis was carried out using version 11.5 of Statistical Package for Social Sciences. Results And Discussion Callus induction Primary calli were formed after 15 days of seed culture on callus induction media. They were characterized by having an intense yellow colour, small size and compact appearance. Small clusters of globular and translucent somatic embryos have developed in 60% of primary calli, after 25 days of culture. The highest frequency of embryogenic callus formation was obtained when the medium were solidified with 0.8% of agar (Figure 1 ). High agar concentration decreased the embryogenic response, regardless of the increase of the 2.4-D concentration. These results suggested that the concentration of agar in the medium was a critical factor for the embryogenic callus formation. Agar influences the callus induction by regulating the humidity of in vitro culture conditions. The effect of genotype may determine the concentration of agar to be used at each stage of in vitro culture. Zaidi et al. (2006) obtained the highest frequencies of callus induction of indica rice (cv. MDU 5) when the culture medium was solidified with 1.2% agar. Pérez Bernal (2007b) has reported that rice calli (cv. IACuba-28) cultured at 0.8% agar showed hiperhidricity. However, in the case of J-104 rice calli the decrease of agar concentration did not cause hiperhidricity, but improved the embryogenic quality of the calli. Perhaps the mobility and absorption of the culture medium nutrients were favoured with the lower agar concentration, stimulating the callogenesis in this cultivar. Moreover, agar could modify the availability of soluble substances by means of chemical interactions (Abdoli et al., 2007). Impact of amino acid supplement on callus growth There were not significant differences (p=0.05) in embryogenic callus formation frequency between N6, N6P and N6G. However, the growth of numerous proembryogenic structures and globular somatic embryos over calli cultured on N6PG was notable (Figure 2a). The effect produced by both amino acids suggested that amino acid interaction was involved in this instance. Significant differences (p=0.05) in relative fresh weight growth of callus were found between treatments with L-proline and/or L-glutamine respect to control without amino acids. Calli cultivated on N6PG exhibited a 2.61 fold increase in fresh weight in respect to calli cultivated on medium without amino acids. Most of callus random samples weighed more than 1.22 g when L-proline, L-glutamine or both were added to callus formation medium (Figure 2b). L-Proline and L-glutamine are commonly included as organic supplement in tissue culture media. The use of 500 mg/L of L-glutamine and L-proline, joined to casein hydrolysate, has been successful for the induction of two lines of wheat callus (Opabode, 2006). Zaidi et al. (2006) demonstrated that the addition of the amino acid L-tryptophan promoting callus induction and regeneration of numerous genotypes of indica rice cv. MDU 5. L-Tryptophan is an essential amino acid which acts as a precursor of the IAA, an important auxin for somatic embryogenesis in cereals (Zaidi et al., 2006) To date, casein hydrolysate was the exclusive amino acid source used for efficient callus induction in other Cuban rice varieties (Pérez Bernal et al., 2008a, 2008b). The present work has proved that the mixture of L-proline and L-glutamine as additional organic nitrogen supplement has significant utility for rice callus development. These amino acids provide a source of reduced nitrogen, which is readily metabolized by plant cells, stimulating faster cell growth (Sarker et al., 2007). Due to the complexity of amino acid metabolism, future research should investigate the properties of amino acids combination that play a major role in tissue culture of J-104 rice cultivar. Effects of inoculation and cocultivation factors on transient GUS expression Inoculation The maximum fluorometric GUS activity was obtained in callus inoculated for 10 min with A. tumefaciens (table 3). Extending inoculation beyond 25 min was not beneficial, due to tissue browning detected after callus dried onto filter paper. Transient GUS expression was improved by a short inoculation time. Further increases in the inoculation time tended to cause reductions in GUS expression, perhaps as a result of a decrease in cell viability. Tissue browning and necrosis following Agrobacterium infection is still a major obstacle in genetic transformation (Kuta and Tripathi, 2005). Calli might be hypersensitive to the bacterial culture if co-cultivation is extended (Pérez Bernal et al., 2008a). A correlation between the reduction in cell death and the improved transformation frequency has been necessary. Determining the optimum inoculation time and co-cultivation period is important, because if they are extended the tissues are colonized by Agrobacterium, and is more difficult to suppress the bacterium during the culture stages. Temperature The calli of Cuban rice varieties are routinely cultured at 28 °C (Pérez-Bernal et al., 2007a; 2008). But this research demonstrated that transient GUS activity markedly increased when the co-cultivation temperature was decreased from 28 to 20 °C. Significant differences in transient activity (p=0.05) were not detected between calli cultured at 25 °C and 28 °C. It was found that 20 °C was better as co-cultivation temperature supporting by the highest transient activity (table 3). This suggests that transient GUS activity depends of temperature. The optimal temperature for co-cultivation should be evaluated with each specific explants, cultivars and Agrobacterium strain involved. In rice, the co-cultivation temperature for most of the cultivars has ranged from 24 °C to 25 °C and, in some cases, 28 °C. Saharan et al. (2004) described the Agrobacterium- mediated transformation of two indica varieties. They used EHA 105 strain containing pCAMBIA 1301, the same used in the present study. But they affirmed that transient GUS expression found to be higher in calli co-cultivated at 28 °C, which is a different result compared to ours. Another studies reported 28.9% and 44.4% of transient GUS expression in indica rice calli of HKR-46 y HKR-126 cultivars, mediated by co-cultivation with Agrobacterium at 25 °C (Kuta and Tripathi, 2005). All these results suggest that researchers should take into account the effect of genotype to determine the optimal co-cultivation temperature. Acetosyringone concentration Co-cultivation of calli with Agrobacterium in the presence of acetosyringone, a vir gene inducer, has become a routine exercise in the transformation of recalcitrant crops such as rice, maize, barley and wheat (Ozawa, 2009). GUS expression was found in calli cocultivated in presence of acetosyringone at 100, 150 and 200 µM. When acetosyringone was omitted the value of fluorometric GUS activity was similar to background value of non-transformed control, indicating the nonexistence of GUS expression (table 3). This result means that J-104 rice calli need the stimulant effects of acetosyringone as vir inducer to get transient GUS expression. The majority of the successful reports on Agrobacterium-mediated transformation of rice have been based on Agrobacterium pre-induction and/or co-cultivation in the presence of acetosyringone. Hashizume et al. (2006) reported that transient expression of the gus gene was not observed in the absence of acetosyringone even when using a super virulent Agrobacterium strain. Hiei and Komari (2006) found that the level of transient GUS expression was small and stable transformed rice plants could not be regenerated when acetosyringone was omitted. The use of acetosyringone 100 µM allowed the maximum quantitative GUS expression in the present work. No significant differences in transient GUS expression were obtained with 150 µM and 200 µM of acetosyringone (table 3). It was observed a necrotic reaction in calli cultivated with 200 µM acetosyringone. These calli turned brown and wet during co-cultivation period and only a few amounts of them could be used for fluorometric GUS assay. The use of 200 µM acetosyringone for genetic transformation of IACuba-28 rice calli (Pérez Bernal et al., 2008a) did not cause the severe tissue damage observed in the J-104 calli. In this case the effect of genotype could determine the in vitro response. In addition, the increase of concentration from 100 µM to 200 µM caused GUS activity reduction, indicative of a possible harmful effect on Ti-DNA transfer (Amoah et al., 2001). This may reflect the observation that, whilst acetosyringone was effective at low concentration, it may act as a bacteriostatic at higher concentrations (Sheng and Citovsky, 1996). Co-cultivation period After 3 days of co-cultivation the calli grew normally and transient GUS expression was slightly improved in respect to calli co-cultivated during 5 days. Calli co-cultivated for 7 days yielded better results than those cocultivated for 3 and 5 days (table 3). But calli exhibited serious damages when they were co-cultivated for more than 5 days, coinciding with results obtained in IACuba-28 rice cultivar (Pérez Bernal et al., 2008a). The browning was caused by excess proliferation of Agrobacterium during co-cultivation, which decreases the frequency of transformation and callus viability (Ozawa, 2009). Kuta and Tripathi (2005) explained that Agrobacterium-induced necrosis often observed in target plant tissues is linked to hypersensitive defense reaction in plants to Agrobacterium infection. The plant defense mechanisms against Agrobacterium involve a rapid and large generation of reactive oxygen radicals in target plant cells, resulting to plant cell death. Therefore, optimization of co-cultivation period is a possible way for improving the efficiency of Agrobacterium mediated transformation of rice, especially in recalcitrant cultivars like J-104. Conclusions Agar concentration and amino acid supplement are critical factors for the embryogenic callus formation in J-104 rice cultivar. High agar concentration decreases the embryogenic response, regardless of the increase of the 2.4-D concentration. The combination of Lproline and L-glutamine has significant utility for increasing embryogenic callus formation frequency. The higher values of quantitative transient GUS expression in calli are obtained under the following conditions: 10 min as inoculation time, co-cultivation media with acetosyringone 100 µM, temperature of 20 °C, and 3 days as co-cultivation period. References 1 Abdoli, M., Moieni, A., Dehghani, H. 2007. Effects of cultivar and agar concentration on in vitro shoot organogenesis and hyperhydricity in Helianthus annuus L Pak J Bot 39: 31-35. [ Links ] 2 Amoah, B. K., Wu, H., Sparks, C.;, Jones, H. D. 2001. Factors influencing Agrobacterium-mediated transient expression of µidA in wheat inflorescence tissue. J Exp Bot 52: 1135-1142. [ Links ] 3 Chu, C. C., Wang, C. C., Sun, C. S., Hsu, C., Chu, C. Y., Bin, F. Y. 1975. Establishment of an efficient medium for anther culture of rice through comparative experiment on the nitrogen sources. Scientia Sinica 18: 659-668. [ Links ] 4 Datta, K., Tu, J. M., Oliva, N., Ona, I., Velazhahan, R., Mew, T. W., Muthukrishnan, S., Datta, S. K. 2001. Enhanced resistance to sheath blight by constitutive expression of infection related rice chitinase in transgenic elite indica rice cultivar. Plant Sci 160: 405-414. [ Links ] 5 Hashizume, F., Nakazaki, T., Tsuchiya, T., Matsuda, T. 2006. Effectiveness of genotype-based selection in the production of marker-free and genetically fixed transgenic lineages: ectopic expression of a pistil chitinase gene increases leaf-chitinase activity in transgenic rice plants without hygromycin-resistance gene. Plant Biotechnol 23: 349-356. [ Links ] Hiei, Y., Komari, T. 2006. Improved protocols for transformation of indica rice mediated by Agrobacterium tumefaciens. Plant Cell Tissue Org Cult 85 (3): 271-283. [ Links ] Jefferson, R. 1987. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol Biol Reporter 5:387-405. Klimaszewska, K., Bernier-Cardou, M., Cyr, D. R., Sutton, B. C. S. 2000. Influence of gelling agents on culture medium gel strength, water availability, tissue water potential, and maturation response in embryogenic cultures of Pinus strobus L. in vitro Cell Dev Biol 36 (4): 279-286. [ Links ] [ Links ] Kopertekh, L. G., Stribnaya, L. A. 2003. Plant Regeneration from Wheat Leaf Explants. Russian J Plant Physiol 50: 365-368. [ Links ] Kuta, D. D., Tripathi, L. 2005. Agrobacterium-induced hypersensitive necrotic reaction in plant cells: a resistance response against Agrobacterium-mediated DNA transfer. Afr J Biotechnology 4 (8): 752-757. [ Links ] Lin, Y. J., Zhang, Q. F. 2005. Optimizing the tissue culture conditions for high efficiency transformation of indica rice. Plant Cell Rep 23: 540-547. [ Links ] Meneses, A., Flores, D., Muñoz, M., Arrieta, G., Espinoza, A. M. 2005. Effect of 2,4-D, hydric stress and light on indica rice (Oryza sativa) somatic embryogenesis. Rev Biol Trop 53 (3-4): 361-368. [ Links ] Opabode, J. 2006. Agrobacterium-mediated transformation of plants: emerging factors that influence efficiency. Biotechnol Mol Biol Rev 1 (1): 12-20. [ Links ] Ozawa, K. 2009. Establishment of a high efficiency Agrobacterium- mediated transformation system of rice (Oryza sativa L.). Plant Sci 176: 522-527. [ Links ] Pérez Bernal, M., Delgado, M., Hernández, C., Armas, R. 2007a. Morphological evaluation of shoots regenerated from hygromycin-resistant rice callus (cv. IACuba-28). Colombian Journal of Biotechnology 9 (1): 35-40. [ Links ] Pérez Bernal, M. 2007b. Transformación genética de la variedad de arroz IACuba-28 mediante Agrobacterium tumefaciens. Magíster Scientiae Thesis. Biology Faculty, University of Havana. 33 p. [ Links ] Pérez Bernal, M., Coll, Y.; Pujol, M., Armas, R., Alfonso, J., Hernández, C.; González, A., Abreu, D., Valdivia, O., Delgado, M. 2008a. Las condiciones de cocultivo determinan la eficiencia de transformación de arroz (var. IACuba-28) mediante Agrobacterium tumefaciens. BioTecnología 12 (3): 21-33. [ Links ] Pérez Bernal, M., Delgado, M., Hernández, C., Armas, R. 2008b. Indirect organogenesis from shoot apical meristems of Cuban rice cultivar Reforma. Cultivos Tropicales 29 (1): 23-28. [ Links ] Saharan, V., Yadav, R. C., Yadav, R. N., Ram, K. 2004. Studies on improved Agrobacterium mediated transformation in two indica rice (Oryza sativa L.). Afr J Biotechnology 3 (11): 572-575. [ Links ] Sarker, K. K., Kabir, A. H., Sharmin, S. A., Nasrin, Z., Alam, M. F. 2007. Improved somatic embryogenesis using L-asparagine in wheat (Triticum aestivum L.). Sjemenarstvo 24: 187-196. [ Links ] Sheng, J., Citovsky, V. 1996. Agrobacterium-plant cell DNA transport: have virulence proteins, will travel. The Plant Cell 8: 1699-1710. [ Links ] Tyagi, H., Rajasubramaniam, S., Dasgupta, I. 2007. Regeneration and Agrobacterium mediated transformation of a popular indica rice variety ADT39. Current Science 93 (5): 678-683. [ Links ] Visarada, R. S., Sarma, N. P. 2002. Qualitative assessment of tissue culture parameters useful in transformation of indica rice. Current Science 82 (3): 343-346. [ Links ] Zaidi, M. A., Narayanan, M., Sardana, R., Taga, I., Postel, S., Johns, R., McNulty, M., Mottiar, Y., Mao, J., Loit, E., Altosaar, I. 2006. Optimizing tissue culture media for efficient transformation of different indica rice genotypes. Agronomy Research 4 (2): 563-575. [ Links ] 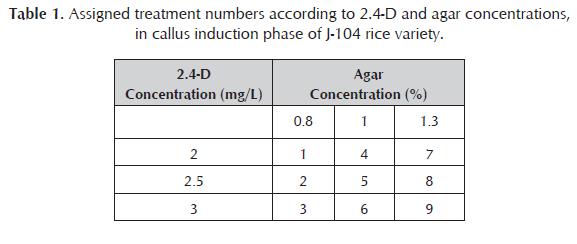



{kind=link}