Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.11 n.2 Bogotá jul./dez. 2009
ARTÍCULO DE INVESTIGACIÓN
Evaluación del efecto del medio MS y la temperatura en el desarrollo de propágulos de Marchantia polymorpha L. (Marchantiaceae) bajo condiciones in vitro y ex vitro
Evaluating the effect of MS medium and temperature in developing Marchantia polymorpha L. (Marchantiaceae) propagules in in vitro and ex vitro conditions
Jaime Alonso Pedroza-Manrique 1 , Mauricio Caballero Arévalo 2
1Biólogo, Esp. M. Sc.; Profesor Asociado, Universidad Distrital Francisco José Caldas. jpedroza@udistrital.edu.co2Licenciado en Biología, Universidad Distrital Francisco José Caldas.
Recibido: septiembre 9 de 2009 Aprobado: noviembre 19 de 2009
Resumen
En este trabajo de investigación se propagaron, bajo condiciones in vitro, propágulos de Marchantia polymorpha evaluando su desarrollo vegetativo mediante observaciones y registros semanales a cada cultivo por tratamiento en función del tiempo de desarrollo. En este contexto, se evaluó cualitativamente la forma y las características del talo, las cuales se valoraron mediante observaciones y registros semanales. Se analizaron los resultados cuantitativos mediante un análisis de varianza con diseño al azar y arreglo factorial 4 x 2, mediante la prueba estadística de Fisher. Las condiciones apropiadas de cultivo para propágulos de M. polymorpha se establecieron en una concentración de 25% de sales minerales Murashige y Skoog (1962), incubados a una temperatura de 25 ± 1 °C. Luego de 13 semanas de desarrollo bajo condiciones in vitro, se adaptaron a condiciones naturales mediante un cambio de sustrato, y controlando la temperatura y humedad en el lugar de desarrollo. En esta etapa se evaluó la sobrevivencia de plantas durante 10 semanas; posteriormente al cambio de condiciones, y como característica cualitativa, se tuvo en cuenta el vigor del talo. Este protocolo de propagación de M. polymorpha, pionero en Colombia, es un modelo que permite conservar y cultivar de manera masiva diferentes especies de briófitos, especialmente aquellos que se encuentran en vías de extinción en nuestro país.
Palabras clave: briófitos, cultivo in vitro, cultivo ex vitro.
Abstract
Marchantia polymorpha propagules were propagated in this research in in vitro conditions to assess their growth in terms of development time. Development rate and contamination regarding each treatment were also evaluated by weekly observations and crop records for each experimental unit per treatment. Talus shape and characteristics were also qualitatively evaluated by weekly observations and records. Fishers statistical test was used for analysing quantitative results by variance analysis with random design and 4 x 2 factorial arrangement. Appropriate conditions for cultivating M. polymorpha propagules were established at 25% Murashige and Skoog mineral salt concentration (1962), incubated at 25°C ± 1°C. After 13 weeks development in in vitro conditions, they were adapted to natural conditions by changing development site substrate and controlling temperature and humidity. Plant survival was evaluated for 10 weeks during this stage. Conditions were then changed and (as qualitative characteristic) talus vigour was taken into account. In was also determined that the ex vitro cultivation level should allow for gradual adjustment to new humidity, temperature and substrate conditions, taking special care that these conditions were not so altered as to become insurmountable. This pioneering M. polymorpha propagation protocol in Colombia is a model for the future maintenance and mass development of different bryophyte species, especially those which are endangered in our country.
Key words: bryophyte, in vitro culture, ex vitro culture.
Introducción
En el mundo, la conservación de briófitos ha tomado vital importancia desde hace más de cien años, esta preservación se ha llevado a cabo en el lugar donde habitan estos organismos in situ; sin embargo, debido a la dificultad de su estudio, al lento crecimiento y fragilidad del hábitat donde se hallan, se ha optado por la conservación ex situ, siendo la propagación bajo condiciones in vitro la más usada en especial en países europeos (Constantino, 1994; Elías et al., 2001; Hernández, 2007). La técnica de cultivo de tejidos vegetales in vitro se ha utilizado ampliamente para la propagación, conservación e investigación de plantas vasculares, especialmente criptógamas en vías de extinción (Chaparro et al., 1997; Barba et al., 2001; Pedroza, 2008). En Londres se ha investigado el cultivo de briófitos bajo condiciones in vitro desde hace cerca de cien años; dentro de las múltiples investigaciones realizadas se han propagado especies en riesgo de extinción, de interés comercial, y especies poco comunes, estableciendo metodologías precisas para su propagación, tales como Funaria, Ceratodon y Sphagnum, entre otras (Duckett et al., 2004). En Colombia no se han realizado investigaciones de esta índole, puesto que el interés de muchos investigadores es la propagación de plantas de interés comercial, o bien conservar plantas en riesgo de extinción pertenecientes a la flora fanerógama, donde sólo se han evaluado comunidades de briófitos y se ha determinado el riesgo de extinción para más de 60 especies, sin aplicarse ninguna estrategia de conservación, teniendo en cuenta que en varios ecosistemas como los páramos altoandinos tienen una larga historia de disturbio y un estrecho vínculo con las actividades humanas, porque los modelos productivos, desarrollados e implementados en la geografía nacional, llevan siglos ocasionando graves alteraciones en los bosques tropicales, transformando grandes extensiones para el desarrollo de monocultivos, ganadería y la minería a cielo abierto (Parra et al., 1999; Linares y Uribe, 2002; Orrego y Uribe, 2004; Lagos et al., 2008; Insuasty, et al., 2009; Mosquera, 2009). Estas perturbaciones generan alteraciones en las comunidades vegetales altoandinas (Cupitra, 2009).
En esta investigación se pretende establecer la propagación y conservación del desarrollo de propágulos de la hepática Marchantia polymorpha L., mediante la evaluación de diferentes concentraciones de sales minerales, para determinar cuál es la concentración más apropiada, porque los briófitos propagados bajo condiciones in vitro toleran diversidad de medios de cultivo y diferentes concentraciones. Por esta razón se eligió el medio Murashige y Skoog (1962) debido a que contiene una formulación básica que sirve de fuente de nutrientes a gran variedad de plantas, además es usado en el cultivo de helechos, los cuales se relacionan un poco con los briófitos, aunque este medio tiene un contenido elevado de sales (Hurtado y Merino, 1987; Pierik, 1990). Los 19 nutrientes esenciales para la mayoría de plantas, incluyendo los briófitos, se encuentran en estos medios de cultivo, tanto los micronutrientes como los macronutrientes (Taiz y Zeiger, 1998; Jaques, 1998; Glime, 2006).
La propagación de briófitos mediante técnicas de cultivo de tejidos vegetales abrirá nuevas posibilidades en la conservación de estos organismos, porque se podrán establecer metodologías de cultivo para cada especie. Por otra parte, servirá de herramienta para la propagación masiva de especies de interés comercial, la obtención de hepáticas y musgos libres de patógenos, así como la conservación del germoplasma de especies raras o de crecimiento tardío como sucede con los antoceros; asimismo, permitirá realizar revisiones de algunas especies y subespecies, las cuales pueden ser incluidas en un nuevo taxón de acuerdo con las características de los tejidos y las células en cultivo porque la vegetación cumple un papel fundamental en la mayor parte de los ecosistemas (Magdaleno, 2009).
Materiales y métodos
La caracterización cuantitativa y cualitativa del desarrollo de propágulos de Marchantia polymorpha L., se obtiene realizando los siguientes pasos:
- a) Colección del material vegetal. 20 briófitos de M. polymorpha fueron colectadas manualmente en zona cercana al edificio de los laboratorios de la sede Macarena B de la Universidad Distrital Francisco José de Caldas, localizada en la Macarena, Bogotá, D. C. (figura 1).

Con respecto a la aclimatación a condiciones naturales, tan pronto se finalizó la semana 13 de evaluación in vitro se procedió a adaptar las plantas a condiciones ex vitro. Esto se realizó tomando suelo fresco, el cual se tamizó con ayuda de un colador para separar las partes grandes, medianas y pequeñas del suelo. En el fondo de los semilleros se colocaron las partes más grandes, seguidas de las medianas y, por último, se colocaron las partes más pequeñas del suelo, luego se regaron con agua hasta alcanzar la capacidad de campo. Posteriormente, se sacó cada planta del frasco de cultivo, se lavó con agua destilada y a continuación se colocó en el suelo del semillero.
Para mantener la humedad se cubrieron los semilleros con su correspondiente cubierta transparente, la cual mantenía la humedad relativa y permitía a la vez el paso de luz. Estos semilleros a su vez se ubicaron dentro de un invernadero en el cual la temperatura promedio era de 22 ± 2 °C y la humedad de 90%. Posteriormente se pudo observar que las plantas continuaron con su desarrollo normal y se inició con la evaluación ex vitro durante 10 semanas. Se observó el aspecto y vigor que presentaba el talo, teniéndose en cuenta cuatro valores cualitativos: excelente (E), bueno (B), regular (R) y malo (M).
Análisis estadístico
Se analizaron los datos numéricos que se obtuvieron del diseño al azar con arreglo factorial 2 x 4. Este arreglo significa dos temperaturas de incubación (19 ± 1 °C y 25 ± 1 °C) y cuatro concentraciones de sales minerales Murashige y Skoog (100, 50, 25 y 0%), para un total de ocho tratamientos que evaluaron el porcentaje de desarrollo y tamaño de las propágulos de M. polymorpha.
Cada tratamiento tuvo 15 repeticiones lo que totalizó 120 unidades experimentales que fueron evaluadas mediante análisis de varianza con la prueba de Fisher, de acuerdo con el siguiente modelo: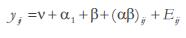 , donde v es la media geométrica, a y b las variables por evaluar, y Eij el error experimental (Steel y Torrie, 1985).
, donde v es la media geométrica, a y b las variables por evaluar, y Eij el error experimental (Steel y Torrie, 1985).
Resultados y discusión
En la evaluación cuantitativa y cualitativa del desarrollo de los propágulos de Marchantia polymorpha L M., el efecto de las temperaturas de incubación y las concentraciones del medios MS presentó diferencias estadísticas altamente significativas (p<0,01) entre los ocho tratamientos evaluados, que a continuación son analizados.
Establecimiento bajo condiciones in vitro de M. polymorpha L. M.
La influencia del material vegetal escogido para realizar la experimentación, en este caso los propágulos, determina en buena parte los resultados obtenidos, debido a diversos factores. Como primera instancia está el genotipo, teniendo como referente la capacidad de regeneración, que en este caso está determinada por la capacidad de multiplicación celular que tiene el propágulo, la cual solo se presenta cuando las condiciones ambientales son apropiadas, estas condiciones las obtuvo de cada uno de los tratamientos, los cuales proveían de temperatura adecuada, y de concentración de sales minerales. Por otra parte, los propágulos están especializados en la propagación vegetativa y se encuentran protegidos al interior de los conceptáculos, por tanto poseen un tejido que no ha sido expuesto a las condiciones del medio y, además, no ha presentado actividad metabólica. Adicionalmente, su estado vegetativo para los propágulos permite que se desarrolle mucho más fácil que las esporas en estado generativo (Pierik, 1990; Jensen et al., 1998; Molphe et al., 1999; Strasburger et al., 2004; Wickett y Goffinet, 2008).
El éxito en el desarrollo y crecimiento de los propágulos también se atribuye a que los medios de cultivo fueron solidificados con ayuda de Phytagel y no con Agar como suele hacerse en la mayoría de cultivos in vitro, esto se hizo debido a que esta sustancia gelificante es menos tóxica para el tejido cultivado, debido a la menor cantidad de impurezas; asimismo, es más eficaz para el desarrollo de hepáticas, sumado a que permite ver más fácilmente las plantas y su proceso de crecimiento, y los signos de presencia de contaminantes (Montoya, 1991; Duckett et al., 2004).
Esta especie, al igual que la mayoría de hepáticas talosas, se desarrolla generalmente sobre suelo o arcilla húmeda, y cuando un propágulo inicia su crecimiento tan sólo necesita de suficiente humedad relativa, la cual se presentaba en todos los tratamientos al 100%, especialmente en aquellos que estaban incubados a 25 ± 1 °C en el Fitotrón (tratamientos 1-4), esta humedad estaba relacionada con la luz y la temperatura, por tanto no era necesario regularla. También necesita un sustrato que le provea nutrientes cuando la planta tenga un grado de madurez mayor, el cual fue sustituido por el medio de cultivo con sales Murashige y Skoog (1962), que es adecuado para diversas especies vegetales, incluidos los briófitos (Scagel, 1973; Hurtado y Merino, 1987; Vasil, 1984).
La contaminación que presentaron algunas unidades experimentales a lo largo de la experimentación puede atribuirse a diversos factores inherentes a la planta, puesto que el material vegetativo, generalmente, contiene gran cantidad de microorganismos en su superficie (Hurtado y Merino, 1987). Ante esta situación, las muestras que presentaron desarrollo estaban libres de patógenos, porque el proceso de desinfección con hipoclorito de sodio es muy efectivo para eliminar los agentes presentes en los propágulos y cualquier parte del tejido de la hepática, en especial las esporas de bacterias, las bacterias en estado vegetativo, protozoarios, hongos, algas y virus (Pedroza y Tupaz, 2008).
Evaluación cuantitativa
Cuando se inició este proceso investigativo, el propágulo se observó de coloración verde hialino a modo de lóbulos redondeados, aunque algunos propágulos permanecían sin desarrollarse.
Con respecto a la evaluación del tamaño y las características morfológicas de las plantas, en los resultados obtenidos semanalmente la mayoría de las plantas presentaban un crecimiento de 1 a 2 mm por semana, el cual era verificado teniendo como punto de referencia su parte apical, puesto que M. polymorpha tiene un grupo de células meristemáticas situadas en esta parte las cuales permiten el crecimiento del gametofito. Este crecimiento era relativamente rápido, puesto que en general las hepáticas tienen un crecimiento lento, aunque algunas especies en condiciones favorables pueden crecer hasta 30 cm al año (Scagel, 1973; Abbayes et al., 1989).
Se observó que desde la primera hasta la tercera semana de evaluación, el crecimiento es gradual aumentando de 3 a 12 unidades experimentales por tratamiento, lo que demuestra la rápida respuesta que tienen los propágulos ante el medio de cultivo en que se hallan. Asimismo, se observó que la cantidad de propágulos que se desarrollaron se mantiene constante hasta la última semana de evaluación in vitro, es decir, hasta la semana 13.
Todos los tratamientos permitieron que se desarrollaran los propágulos incubados en el interior de cada unidad experimental; sin embargo, presentaron diferencias en cuanto al tiempo de respuesta y desarrollo. Al cabo de la primera semana, en los tratamientos 4 y 8 se evidenció un crecimiento inmediato. En el tratamiento 4 (0% medio MS y 25 ± 1 °C), 14 de las 15 unidades experimentales permitieron el desarrollo de propágulos lo cual indica que estos tratamientos activan el desarrollo de los propágulos más rápidamente. El tratamiento ocho (0% medio MS y 19 ± 1 °C) por su parte, permitió el crecimiento de seis unidades experimentales en la primera semana y durante las 12 semanas siguientes siete unidades experimentales lograron desarrollar los propágulos en su interior (figura 2).
El tratamiento 3 (25% medio MS y 25 ± 1 °C) evidenció un rápido desarrollo de propágulos en cada una de sus unidades experimentales, puesto que en la segunda semana, 11 unidades experimentales desarrollaron propágulos. Posteriormente, en la tercera semana, aumentó a 14 unidades experimentales y al final de la evaluación, en la semana 13, las 15 unidades experimentales en total permitieron el desarrollo de la planta. Los propágulos de M. polymorpha, al ser estructuras de reproducción vegetativa, poseen una ventaja porque crecen directamente sin pasar por la fase de protonema, que en condiciones naturales se origina por la germinación de la espora, lo cual permitió evidenciar su desarrollo a simple vista en cada unidad experimental. Esta condición a su vez hizo posible que el desarrollo de la planta fuera mucho más rápido, sumado a la asepsia existente en los tratamientos, los cuales facilitaron el desarrollo de talos libres de patógenos (Izco et al., 2004; Hurtado y Merino, 1987).
En los tratamientos 1 y 5, la concentración de sales minerales Murashige y Skoog (1962) en el medio de cultivo era del 100%, esto revela que recibían la misma cantidad de nutrientes; sin embargo, hay diferencia en el volumen de la planta o cantidad de biomasa, y en el tamaño del talo.
Es por ello que la influencia de la temperatura determinó esa diferencia entre los tratamientos, siendo 25 ± 1 °C la temperatura que permitió un desarrollo superior de las plantas incubadas en el tratamiento 1, debido a que los briófitos en condiciones in vitro se desarrollan bien entre 5 y 25 °C; no obstante, para especies tropicales se recomienda entre 20 y 25 °C (Duckett et al., 2004; Vasil, 1984).
En los tratamientos 2 y 6, la concentración de sales minerales Murashige y Skoog (1962) en el medio de cultivo era del 50%. No obstante, se observan plantas con mejor vigor y mayor biomasa que las anteriores, notándose así que un medio diluido a la mitad presenta mejores resultados, puesto que los requerimientos de minerales no son tan altos, provocando un mayor potencial hídrico que favorece su desarrollo vegetativo.
Adicionalmente, el desarrollo era evidentemente superior en el tratamiento dos, lo cual de nuevo se relaciona con la temperatura de incubación. Que estos tratamientos permitieran un aumento de biomasa aceptable se atribuye al hecho no solo de las condiciones de cada tratamiento en sí, sino a que estas condiciones facilitaron la acción de las citoquininas que la planta sintetiza; este fitorregulador de crecimiento está presente en sus células, al igual que en muchas plantas vasculares y, en este caso, se encargó de incrementar la multiplicación celular, que se puede evidenciar en el aumento de biomasa (Hurtado y Merino, 1987; Pedroza, 2008).
En los tratamientos 3 y 7, la concentración de sales minerales Murashige y Skoog (1962) en el medio de cultivo era del 25%, esta concentración tuvo el mejor efecto en el desarrollo de los propágulos puesto que, al observar las plantas, el vigor, la biomasa y la coloración de los talos era notoriamente superior al de los demás tratamientos. Pero tal como sucedió con dichos tratamientos, existe una clara diferencia entre el tratamiento incubado a temperatura de 25 ± 1 °C, el cual tiene un talo con mayor biomasa, y el incubado a 19 ± 1 °C, que presenta un talo muy delgado y de gran longitud. Generalmente, los cultivos se mantienen en una temperatura constante entre los 20-30 °C. En este contexto, los termoperiodos y los pretratamientos de temperatura (incluyendo una temperatura de enfriamiento) han mostrado efectos en la morfogénesis. Dentro del mismo tejido han sido observados diferentes óptimos de temperatura para la formación de brotes y el enraizamiento (Cheng y Voqui, 1977; Rumary y Thorpe, 1984).
En atención a que en esta experimentación no se utilizaron los fitorreguladores de crecimiento, en el tratamiento 7 se ve una clara acción de las auxinas que la propia planta sintetizó, porque la elongación del talo se debe a la acción de este fitorregulador de crecimiento que actúa directamente en la elongación celular, evidenciándose un crecimiento por alargamiento. En este contexto, es importante tener en cuenta que la proliferación de los diferentes explantes que se utilizan en el cultivo in vitro puede lograrse con el uso de reguladores de crecimiento como componentes principales de un medio de cultivo previamente establecido y un adecuado manejo del proceso in vitro cuando en los explantes evaluados no existe un balance de fitorreguladores (Pedroza, 2008).
Adicionalmente, en el tratamiento 3 se ve una acción combinada de auxinas como de citoquininas, al igual que con los tratamientos 2 y 6, por su notorio aumento de biomasa semanal. Este aumento también se puede atribuir a la concentración adecuada de Nitrógeno presente en el medio de cultivo, el cual tiene una influencia directa sobre el aumento de la biomasa y elongación del talo (Salemaa et al., 2008; Sabovljevic et al., 2003).
En los tratamientos 4 y 8, la concentración de sales minerales Murashige y Skoog (1962) en el medio de cultivo era del 0%, lo que indica que los nutrientes obtenidos por las plantas los extraían del azúcar inmerso en el medio (myoinositol) y del gelificante en sí, en este caso Phytagel, puesto que contiene pequeñas cantidades de micronutrientes (Duckett et al., 2004; Pedroza, 2008). Además, es evidente la deficiencia de nutrientes que sufrieron las plantas durante su desarrollo, puesto que tenían poco vigor en el talo, su coloración verde casi no era aparente, además de su corta longitud y área de cobertura. Es importante tener en cuenta que las hepáticas suelen crecer de manera apropiada en medios de cultivo libres de nutrientes, incluso en ocasiones tienen mejor desarrollo que los tratamientos con algún tipo de solución de nutrientes en el medio. Si bien esto puede ser algo inadvertido, la falta de crecimiento de algunos propágulos en el tratamiento 8, también se puede atribuir a que sufrieron un daño severo en su tejido al momento de ser sometidos al proceso de desinfección, y por tanto no pudieron iniciar el desarrollo (Vasil, 1984).
Evaluación cualitativa
El registro de características cualitativas del talo evidenció un leve desarrollo el cual se pudo observar por el aumento en volumen del propágulo, convirtiéndose en el talo de la planta, en esta etapa inicial también se observaron pequeños rizoides que crecían de forma desorientada sobre la superficie del medio de cultivo. Este desarrollo estuvo coadyuvado porque el propágulo, al igual que el gametofito, contiene en su interior células mucilaginosas y oleaginosas que le proveen nutrientes para iniciar su desarrollo (Scagel, 1973; Bold et al., 1989; Abbayes et al., 1989; Gimeno et al., 2001; Wickett y Goffinet, 2008). Además, se esperaba una respuesta positiva a los tratamientos, puesto que estas plantas se reproducen fácil y apropiadamente de forma vegetativa de cualquier parte de la planta, pero debido a que se usó una estructura especializada, como los propágulos, que tienen la capacidad de producir un gametofito de forma rápida y efectiva, los resultados obtenidos dan muestra de la confianza de la técnica de cultivo in vitro.
De acuerdo con las características del talo que fueron observadas y registradas, se determinaron 2 tipos de crecimiento: talo lobulado y talo acintado, estas formas de crecimiento podían cambiar a medida que se desarrollaba la planta, además de ser formas alternas una entre otra (figura 3). El desarrollo de los propágulos se iniciaba generalmente con un aumento del volumen del propágulo que se observaba en forma lenticular, sin embargo, esta fase era muy corta, porque el propágulo adquiría una forma lobulada que podía permanecer durante varias semanas o bien tener un alargamiento, lo que cambiaba la forma del talo a una forma acintada.
Igualmente, dentro del desarrollo de los propágulos era evidente la simetría dorsoventral característica de las hepáticas talosas como M. polymorpha, puesto que sobre la cara ventral se desarrollaban los rizoides y sobre la dorsal el tejido fotosintético. Adicionalmente, se generaliza la forma del gametofito a una forma laminar laciniada que es característica de este tipo de talos (Scagel, 1973). No obstante, estas formas de crecimiento son variadas, puesto que presentan diversidad de acuerdo con los medios donde vivan, si se cambia la humedad o el sustrato, la forma de la planta cambia, es por ello que las formas de los briófitos se usan para determinar la naturaleza de un microclima.
De otra parte, el talo podía presentar tres formas de crecimiento con respecto al sustrato, como se observa en la figura 4: 1) talo postrado, el cual permanecía con la superficie ventral del talo completamente adherida al medio de cultivo y solamente las partes apicales presentaban un levantamiento; 2) talo levemente erguido, el cual desde el inicio del desarrollo presentaba una inclinación causada por los rizoides, que no penetraban en el medio y que mantenían el talo un poco hacia arriba; 3) talo erguido, el cual desde el inicio del crecimiento presentaba una disposición casi perpendicular con respecto al sustrato, al igual que el talo levemente erguido; esta posición se debía a que los rizoides mantenían el talo en esa posición, aunque cuando el talo adquiría un mayor desarrollo, esta posición variaba un poco.
Otra posible causa de los crecimientos erguidos y levemente erguidos es la polaridad del propágulo, pues esta determina la dirección hacia donde se debe dirigir el crecimiento. También se puede atribuir a la dureza del medio de cultivo en algunas unidades experimentales y tratamientos, puesto que los rizoides, en ese caso, no pudieron penetrar la superficie del medio y por tanto no pudieron fijarse como lo hacen en medio natural, es decir, no se extendieron dentro del sustrato ni tampoco paralelamente sobre él, característica que es inherente a los patrones de crecimiento de los rizoides, por tanto, ejercieron una presión sobre el talo, de tal manera que lo obligaron a tener este tipo de inclinación. Adicionalmente, el dosel de los briófitos a pequeña escala contribuye al flujo de agua en los ecosistemas, por tanto es muy importante en la diversidad microbiana y de microinvertebrados (Scagel, 1973; Sastre et al., 2009).
A pesar de esto, los rizoides tienen la función importante de la conducción capilar de agua, independientemente de la posición que adopten con respecto al sustrato (Schofield, 1996; Abbayes et al., 1989). Un método que pudo evitar el tipo de crecimiento erguido y levemente erguido era usar cajas de Petri como recipientes de cultivo, puesto que son los más convenientes en cuanto al establecimiento, manipulación y manutención del material vegetal proveniente de los briófitos, además de que pudo evitar que el talo se elongara en esas posiciones, haciendo que se adhiriera de forma casi total al sustrato durante su desarrollo (Duckett et al., 2004).
El tratamiento 1 evidenció crecimiento a partir de la segunda semana, sin embargo, el talo presentó pocos cambios al transcurrir la evaluación, solo hasta la sexta semana comenzaron a evidenciarse cambios de estructura y aumento de volumen de una manera eficiente, y en las últimas 3 semanas aumentó el área de cobertura sobre el sustrato y las ramificaciones dicotómicas
El tratamiento 2 demostró crecimiento a partir de la segunda semana, el talo presentó cambios notorios a partir de la semana 4, adquiriendo un aspecto vigoroso y con ramificaciones dicotómicas que posteriormente continuarían aumentando el volumen y la cobertura del talo sobre el medio. A partir de la semana 8, el talo de la planta adquirió un vigor excelente, presentando un desarrollo apropiado, gran cantidad de rizoides y un talo con una coloración uniforme, que finalmente daría como resultado una planta de muy buen porte, lista para el trasplante al finalizar la evaluación bajo condiciones in vitro
El tratamiento 3, al igual que los dos tratamientos anteriores, evidenció crecimiento a partir de la segunda semana, el talo presentó cambios muy notorios semana a semana, el desarrollo de las unidades experimentales que se hallaban en este tratamiento fue muy rápido, con cambios evidentes con respecto a los demás tratamientos. éste fue, sin duda, uno de los mejores tratamientos de la experimentación. El talo tuvo un aspecto vigoroso a partir de la sexta semana, aumentando su volumen y grosor casi el doble cada semana. Presentó un desarrollo apropiado, gran cantidad de rizoides y un talo con una coloración uniforme, incluso al finalizar la evaluación bajo condiciones in vitro las partes apicales del talo que se encontraban más adheridas al sustrato comenzaron a adherirse a las paredes del frasco de cultivo (figura 5).
El tratamiento 4 evidenció crecimiento a partir de la primera semana, siendo uno de los primeros en mostrar respuesta al mismo. Sin embargo, en este tratamiento los propágulos se caracterizaron con pocos cambios de desarrollo porque su evolución fue mínima y con algunas deficiencias.
Además de esto, la planta no aumentó el área de cobertura sobre el sustrato. Por esta razón, se consideró que es uno de los tratamientos con menor efectividad sobre el desarrollo de los propágalos.
El tratamiento 5 favoreció el crecimiento a partir de la segunda semana, el talo presentó cambios notorios progresivos a partir de la cuarta semana; el grosor del talo, su vigor y área de cobertura aumentaban semana a semana de una manera gradual sin presentar cambios abruptos. Las plantas tuvieron un tamaño relativamente pequeño comparado con otros tratamientos en que el vigor del talo era similar. Al finalizar la evaluación las plantas de este tratamiento, aunque no tuvieron un tamaño ideal, tenían un vigor suficiente para ser traspasadas a condiciones naturales.
El tratamiento 6 reveló crecimiento a partir de la segunda semana, el talo presentó cambios notorios a partir de la quinta semana, adquiriendo un talo ramificado, vigoroso y con un volumen adecuado. La cobertura del talo sobre el sustrato era algo limitada debido a que su crecimiento era erguido, lo cual impedía que se postrara sobre el medio de cultivo. Sin embargo, la planta adquirió un vigor excelente, presentando un desarrollo apropiado, gran cantidad de rizoides y una coloración uniforme en el talo.
El tratamiento 7 evidenció crecimiento a partir de la segunda semana, pero en contraposición a los demás tratamientos en esta semana tenía un tamaño que hacía los talos muy evidentes a simple vista. En este tratamiento se observa claramente cómo, de una semana a la siguiente, el cambio morfológico del talo es muy amplio, y en particular, el talo crece en forma erguida (figura 6).
El tratamiento 8 presentó pocos cambios, el desarrollo de su talo fue muy lento, con escasos rizoides y un vigor regular del talo. Además de esto, el grosor del gametofito y el área de cobertura eran mínimos, debido a que la planta estaba creciendo de forma erguida. A partir de la novena semana la planta comenzó a perder la coloración verde, presentando una coloración marrón. Adicional a esto, el talo no aumentó de manera sustancial su volumen ni su tamaño. Por esta razón, se consideró que es el tratamiento con menor efectividad sobre el desarrollo de los propágalos.
En términos generales, los resultados obtenidos permiten señalar que el tratamiento más efectivo para el cultivo de propágulos de M. polymorpha fue el tratamiento 3, en el desarrollo de los propágulos, de acuerdo con el objetivo de esta investigación. De hecho, se logró el 100% de propágulos desarrollados en este tratamiento en la semana 13, mientas que los tratamientos 1, 4 y 7 alcanzaron el 93,3% de desarrollo vegetativo, el tratamiento 2 el 73,3%, los tratamientos 5 y 6 el 66,6%, y el tratamiento 8 el 46,6% (figura 7).
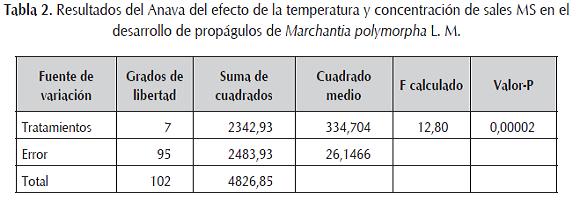
En este contexto, el Anava evidencia que la varianza de respuesta de los tratamientos (Medio MS y temperatura) tiene una diferencia estadísticamente significativa entre la media de respuesta entre un nivel de tratamiento y otro, con un nivel del 95,0% de confianza (tabla 2).
Evaluación de la aclimatación y desarrollo en condiciones ex vitro de M. polymorpha Se inició la evaluación con 94 vitrogametofitos los cuales estaban en su correspondiente contenedor del semillero. Se determinó la sobrevivencia a las nuevas condiciones ambientales mediante observación del vigor del talo y su capacidad fotosintética, la cual se comprobaba por la presencia de coloración verde en el talo del gametofito. Cuando la planta carecía de esta pigmentación (clorosis), era indicio de que había perdido la clorofila y, por tanto, su capacidad de mantener vivo el tejido, razón por la cual se concluía que la planta no se adaptó a las condiciones ex vitro. Semanalmente se realizó este registro, el cual no indicó pérdidas de plantas durante la evaluación, salvo en la semana 10, en la cual murieron 24 plantas posiblemente debido a un aumento fuerte en la temperatura que superó los 30 °C en el invernadero, que es irremediable para las plantas o que causa grandes daños en su estructura. De hecho, los estudios realizados en las últimas décadas han establecido las potenciales amenazas del cambio climático para la biodiversidad (Duckett et al., 2004; Wickett y Goffinet, 2008; Jácome y Vieira, 2009). Durante la fase de aclimatación al nuevo sustrato las plantas continuaron con el desarrollo de forma adecuada, tal como lo venían haciendo desde la fase in vitro, incluso las plantas que tenían pocas probabilidades de sobrevivir, debido a la poca biomasa de su talo, lograron aumentar su tamaño y mejorar su vigor. Al momento de cambiar el sustrato se notó que en la parte basal del talo se hallaba una formación negruzca, la cual contenía las partes necrotizadas del talo, es decir, las partes más veteranas del gametofito (Abbayes et al., 1989). Este material necrotizado evidencia la formación de un nuevo sustrato, el cual es la base sobre la cual continúa la sucesión vegetal, puesto que esta materia orgánica puede acumular agua y la liberación de bióxido de carbono a la atmósfera, sales minerales sobre el cual puede llegar una planta vascular y adaptarse a este nuevo sustrato (Scagel, 1973; Strasburger et al., 2004; Cuevas et al., 2009). Otra de las características que se notó en las plantas en la evaluación bajo condiciones in vitro es que los talos presentaban una superficie muy delgada, la cual se debía a que las paredes de las células de la epidermis se encuentran poco cutinizadas, además, los briófitos como M. polymorpha carecen de tejidos vasculares, presentan células capaces de lignificar debido a la alta humedad que se presenta en cada unidad experimental, razón por la cual las plantas estaban más susceptibles a no sobrevivir al traspaso a nuevas condiciones naturales, puesto que la cutícula protege el tejido interno de la planta y evita la obstrucción de los poros de la superficie. De hecho, las plantas no vasculares son utilizadas como bioindicadores gracias a sus características morfológicas. La ausencia de cutícula y sistema excretor permite que tanto agua y nutrientes, como agentes contaminantes sean absorbidos por todo el organismo (Novo, 2008; Simijaca et al., 2009). Adicionalmente, las plantas que se encontraban en muy mal estado fisiológico fueron traspasadas al nuevo medio a fin de proveerle un sustrato similar, en el que finalmente las plantas se renovaron. Es importante tener en cuenta que los briófitos tienen la característica inherente de colonizar diversidad de sustratos, algunos muy inusuales, los cuales no serían colonizados por plantas vasculares, lo que les confiere cierta ventaja en este procedimiento; de hecho, M. polymorpha forma asociaciones simbióticas con especies de cianobacterias siendo las especies de Nostoc las más frecuentemente encontradas (Pierik, 1990; Schofield, 1996; Frahm, 2003; Mayz, 2004). En este procedimiento es muy alta la tasa de mortalidad de plantas, puesto que implica un cambio un poco destemplado, al cual muchas plantas no logran aclimatarse y por tanto no sobreviven. Es por ello que posterior al cambio de sustrato se controlaron los factores ambientales, haciéndolos intermedios al cultivo in vitro y al cultivo ex vitro, sin olvidar mantener la ecofisiología propia de estas plantas en la que debe predominar la humedad; asimismo, es especialmente importante que la capacidad de retención de agua del nuevo sustrato sea la adecuada. Además, uno de los requisitos fundamentales que debe cumplir el sustrato para su utilización es la sanidad. Cuando no se almacenan o manejan correctamente, pueden contaminarse y provocar serios daños a las plantas durante la aclimatización (Hurtado y Merino, 1987; Abbayes et al., 1989; Schofield, 1996; Suárez, 2005; Pedroza, 2008; Wickett y Goffinet, 2008). Las plántulas in vitro son con frecuencia mixotróficas o heterotróficas (Grout, 1988), aunque la fijación del carbón puede ser estimulada al incrementar la intensidad de luz y la concentración de CO2 (Kozai, 1991). De esta manera, durante la aclimatación se debe desarrollar un metabolismo autotrófico. Las plántulas in vitro crecen normalmente en una densidad de flujo de fotones fotosintéticos baja. Un incremento repentino en la intensidad de luz durante la aclimatización orienta a la fotoinhibición y sirve como una fuente estresante (Van Huylenbroeck, 1994; Van Huylenbroeck et al., 1995). Todos los vitrogametofitos se adaptaron bien, de hecho los provenientes de los tratamientos 2, 3, 6 y 7 presentaron un desarrollo permanente, sin embargo a partir de la semana 8, de forma aleatoria algunas plantas fueron atacadas por algunos patógenos. Se percibió que los talos presentaban zonas mordidas, las cuales fueron atribuidas a moluscos del género Arion, encontrados ocasionalmente en los contenedores. Además de estos ataques, se presentaron zonas con clorosis, debido posiblemente a un déficit de nutrientes, y otras con necrosis, que igualmente pueden atribuirse a déficit nutricional o a una elevada temperatura ambiental. A pesar de esto, la secuencia de las plantas evidenció que el crecimiento en esta etapa se acelera, demostrando la efectividad de las nuevas condiciones y el buen estado de la mayoría de gametofitos. El tratamiento 3 facilitó gametofitos con variados estados de vigor de talo, la mayoría en estado bueno al momento del trasplante, algunas con vigor del talo excelente. Permanecieron con este vigor, mejorando progresivamente; pasaron a un estado excelente a partir de la sexta semana, y continuaron con este mismo vigor hasta la novena semana. En la décima semana, al igual que sucedió con el tratamiento anterior (2), el talo presentó necrosis general en todo el gametofito (figura 8). En términos generales, los gametofitos obtenidos bajo condiciones in vitro presentaron notorias diferencias, especialmente en cuanto a su desarrollo in vitro, el cual determinó su facilidad de aclimatación a condiciones ex vitro, por ello se encontraron diversas respuestas ante el nuevo sustrato y las condiciones ambientales a las cuales fueron sometidas las plantas. En esta fase, cada planta tuvo una aclimatación diferente, y por tanto, sus características no eran tan homogéneas como en la fase in vitro. Debido a que todos los tratamientos se sometieron a condiciones similares, lo que se evaluó principalmente fue la capacidad para adaptarse al nuevo medio, teniendo como referencia el vigor del talo. Además, se observó que las plantas que provenían de condiciones in vitro con mayor biomasa tuvieron más probabilidad de sobrevivir. Es por ello que los tratamientos 2, 3, 6 y 7, mantuvieron un buen vigor del talo en casi toda la evaluación, debido principalmente a que al poseer mayor biomasa, tienen mayor cantidad de tejido parenquimático, el cual permite que el gametofito pueda regenerarse o continuar el desarrollo, debido a la presencia de reservas nutricionales (Abbayes et al., 1989). En los gametofitos del tratamiento 3 también se observó la presencia de conceptáculos hacia los márgenes del talo, como es característico en esta especie, lo cual es un indicio de la eficiencia del tratamiento y la madurez que llega a alcanzar la planta (Schofield, 1996). La sobresaliente biomasa presente en los tratamientos, especialmente en los 2 y 3, no solo es indicio de un buen desarrollo, sino que además facilita en el talo el almacenamiento de agua, puesto que tanto a nivel simplasto y apoplasto, la hepática mantiene el agua que recibe del entorno húmedo (Frahm, 2003; Strasburger et al., 2004). Adicionalmente, los gametofitos provenientes de los tratamientos 1 y 5 iniciaron con un vigor muy regular en casi todas las unidades experimentales, y con el transcurrir de la evaluación fueron mejorando; a pesar de esto, al menos la mitad de las unidades experimentales de ambos tratamientos lograron adaptarse a estas condiciones. Por último, los gametofitos provenientes de los tratamientos 4 y 8, tenían un vigor del talo malo y no se predecía un buen resultado en cuanto a aclimatación; no obstante, los gametofitos del tratamiento 4 mantuvieron un desarrollo general regular, sin aumento significativo de la biomasa, y los gametofitos del tratamiento 8 lograron finalizar la evaluación con talos buenos y algunos excelentes. La mortandad de plantas en la semana 10, en la cual finalizaba la evaluación de la fase ex vitro, fue relativamente alta, puesto que el 20% de los talos se murieron. Estos gametofitos tenían un aspecto seco, sin embargo, al tocarlos se evidenciaba que estaban húmedos y el talo no estaba acartonado (como es característico de una planta seca), además presentaban una coloración marrón negruzco. Estos resultados indican que un aumento violento de temperatura, que superó los 30 °C dentro del invernadero, fue la causa de este suceso, no obstante es una circunstancia extraña, puesto que las hepáticas pueden tolerar la desecación y reponerse cuando las condiciones vuelven a ser favorables, a menos que el periodo de deshidratación del gametofito haya sido muy extenso. Incluso las M. polymorpha suelen enrollarse hacia dentro con el fin de soportar la desecación, aunque esta característica no se evidenció en ninguna de las plantas. La cifra de plantas muertas no incluye aquellas que estaban parcialmente necrotizadas, ni las que tenían el talo destrozado por ataques de patógenos, los cuales pudieron ser prevenidos al esterilizar el suelo antes del trasplante (Schofield, 1996; Duckett et al., 2004; Oliver et al., 2005). De acuerdo con Hernández (2007), las técnicas ex situ se configuran actualmente como una eficaz medida para la protección de especies y ecosistemas en sus hábitats naturales. Su carácter complementario no altera su función imprescindible a la hora de garantizar la conservación de la diversidad vegetal en el planeta. Se ha producido un profundo cambio en sus objetivos, métodos y perspectivas: son medidas preferentemente adoptables en todos los países del mundo, que no alteran sino refuerzan la soberanía y accesibilidad a los recursos genéticos. Manejan ahora técnicas innovadoras, desde la biología molecular a los sistemas de información geográfica, conservan cada vez más íntegramente la variación genética de las especies, estimulan la cooperación internacional, y son una respuesta muy eficaz ante problemas tan serios para la biodiversidad como el cambio climático. Las oportunidades que les brinda el nuevo marco internacional regulado por convenios y tratados como Cites, IT (FAO), y especialmente CBD, junto a las oportunidades que les pueden brindar el desarrollo del Régimen Internacional de Acceso y Distribución de Beneficios, de la Estrategia Mundial de Conservación de Plantas y la respuesta al Cambio Global, deben ser causa de un inmediato refuerzo institucional que permita a los jardines botánicos, herbarios, arboretos y, más singularmente, a los bancos de germoplasma, gozar del apoyo y reconocimiento adecuado sin menoscabo ni tergiversación de sus fines y objetivos. En términos generales, es importante tener en cuenta que la propagación y conservación de los briófitos es un procedimiento indispensable para evitar su extinción y la afectación ambiental de nuestro planeta; de hecho, el establecimiento del protocolo de micropropagación de M. polymorpha es una alternativa de gran interés en la recuperación y el mantenimiento de nuestros ecosistemas afectados. La diversidad de una comunidad depende tanto de la diversidad de especies como de la diversidad de las interacciones ecológicas (Melgarejo, 2009; Zamora, 2009). En este contexto, los briófitos como M. polymorpha rápidamente colonizan los sustratos desnudos que de acuerdo con las características físicas de pendiente, micrositio disponible, especies y formas de crecimiento, confieren diverso grado de compactación y estabilidad al sustrato, en consecuencia controlan el impacto de la gota de lluvia y atenúan la erosión, ya que tienen la capacidad de retener el agua, proteger los suelos, y ser el hábitat de miles de especies de microfauna, entre otras (Bolaños et al., 2009; Rivera y Linares, 2009). Adicionalmente, una de las herramientas con mayor potencial para la gestión de conservación es la restauración de poblaciones, a fin de conectar la restauración con la biología de conservación de una manera directa (Escudero e Iriondo, 2009). Conclusiones En esta investigación se evidenció que M. polymorpha L. se desarrolla adecuadamente en condiciones in vitro. Todos los tratamientos permitieron que se desarrollaran los propágulos incubados en el interior de cada unidad experimental, aunque presentaron diferencias en cuanto al tiempo de respuesta y desarrollo, siendo su crecimiento más rápido y notorio en aquellas que se encontraban incubadas a 25 ± 1 °C de temperatura, con una concentración de sales minerales MS de 25%. En la evaluación cuantitativa y cualitativa de la longitud de los talos se encontró una diferencia significativa entre los tratamientos. En este contexto, el tratamiento 3 fue el que presentó el 100% de desarrollo vegetativo con mejores características puesto que el vigor de las plantas fue excelente y su biomasa era notoriamente superior a la encontrada en los gametofitos de los demás tratamientos. De la misma manera, se observó que las plantas que provenían de condiciones in vitro con mayor biomasa tuvieron mayor probabilidad de sobrevivir. Es por ello que los tratamientos 2, 3, 6 y 7, mantuvieron un buen vigor del talo en casi toda la evaluación, debido principalmente a su mayor biomasa, la cual tiene más cantidad de tejido parenquimatoso, el cual permite que el gametofito pueda regenerarse o continuar el desarrollo debido a las reservas nutricionales. No obstante, al finalizar la evaluación se presentó una muerte masiva de plantas por causas que se atribuyen a un aumento de la temperatura ambiental. Agradecimientos Este trabajo fue financiado por la Facultad de Ciencias y Educación de la Universidad Distrital Francisco José de Caldas. Referencias bibliográficas 1 Abbayes, H., Chadefaud, M., Feldmann, J., De Ferré, Y., Gaussen, H., Grassé P., Prevot, A. R. 1989. Botánica: Vegetales inferiores. España: Reverté. 746 p. [ Links ] 2 Barba, A., Luna, B., Romero, J. 2001. Micropropagación de plantas. México: Trillas. 107 p. [ Links ] 3 Bolaños, G. Y., Feuillet, C., Bolaños, A. C. 2009. Diversidad de briófitos terrestres de un fragmento de bosque nublado en el Km 18 vía Cali-Buenaventura, Valle del Cauca. San Juan de Pasto: Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombina de Botánica. pp. 120. [ Links ] 4 Bold, H., Alexopoulos, C., Delevoryas, T. 1989. Morfología de las plantas y los hongos. España: Omega. 911 p. [ Links ] 5 Chaparro, B., Marquinez, X., Caro, A. 1997. Micropropagación in vitro de Vallea stipularis. Colombia: CAR. 33 p. [ Links ] 6 Cheng, T., Voqui, T. 1977. Regeneration of Douglas fir plantlets through tissue culture. Science, 198: 306- 307. [ Links ] 7 Constantino, S. 1994. Propagación in vitro de un helecho arborescente (Dicksonia sellowiana Hooker 1844) amenazado: una alternativa para su preservación ex situ. Bogotá, Trabajo de grado (Biólogo). Pontificia Universidad Javeriana, Facultad de Ciencias Básicas, Departamento de Biología. 157 p. [ Links ] 8 Cuevas, R. M., Hidalgo, C., Etchevers, J., Campo, J. 2009. Dinámica de la fracción activa de la materia orgánica del suelo en bosques tropicales estacionalmente secos a lo largo de un gradiente de precipitación. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. p. 35. [ Links ] 9 Cupitra, O. S. 2009. Perturbaciones en la diversidad vegetal por pérdida de cobertura por implementación de modelos productivos en los municipios de Timba y Suárez en el departamento del Cauca. Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. p. 64. [ Links ] 10 Duckett, J., Burch, J., Fletcher, P., Matcham, H., Read, D., Russell, A., Pressel, S. 2004. in vitro cultivation of bryophytes: a review of practicalities, problems, progress and promise. Journal of Bryology 26: 3-20. [ Links ] 11 Elías, M. J., Sánchez, J. A., García, J. M. 2001. Hepáticas y antocerotas (Marchantiopsida y Anthocerotopsida) de Salamanca (España). Botánica Complutensis 25: 207-212. [ Links ] 12 Escudero, A., Iriondo, J. A. 2009. Restauración de poblaciones de plantas amenazadas. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. p. 67. [ Links ] 13 Frahm, J. 2003. Manual of tropical bryology. Tropical Bryology 23. 194 p. [ Links ] 14 Gimeno, C., Puche, F., Segarra, J. G., Laguna, E. 2001. Modelo de conservación de la flora briológica en la comunidad Valenciana: microrreservas de la flora criptogámica. Botánica Complutensis 25: 221-231. [ Links ] 15 Glime, J. 2006. Bryophyte Ecology: Physiological Ecology. Estados Unidos: Michigan Technological University. 593 p. [ Links ] 16 Grout, B. W. 1988. Photosynthesis of regenerated plantlets in vitro and the stresses of transplanting. Acta Hort 230: 129-135. [ Links ] 17 Hernández, J. E. 2007. Jardines botánicos y bancos de germoplasma: la conservación ex situ frente a las perspectivas de una estrategia para la conservación de las plantas. Ambienta. pp. 40-46. [ Links ] 18 Hurtado, D., Merino, M. 1987. Cultivo de tejidos vegetales. México: Trillas. 232 p. [ Links ] 19 Insuasty, J., Vargas, J. O., Cárdenas, C., Rojas, O., Gómez, P. A. 2009. Buscando estrategias de restauración ecológica en los páramos: el caso de áreas afectadas por pastoreo, parque nacional natural (PNN) Chingaza, Colombia. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. pp. 115-116. [ Links ] 20 Izco, J., Barreño, E., Bruguéz, M., Costa, M., Devesa, J., Fernández, F., Gallardo, T., Llimona, X., Salvo, E., Talavera, S., Valdés, B. 2004. Botánica. 2 ed. España: McGraw Hill. 906 p. [ Links ] 21 Jácome, J., Vieira, M. I. 2009. Establecimiento de sitios piloto para el establecimiento del impacto del cambio en la biodiversidad de ecosistemas de alta montaña en Colombia. Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. pp. 70. [ Links ] 22 Jaques, M. 1988. Multiplicación vegetativa y cultivo in vitro. Los meristemos y la organogénesis. España: Mundi- Prensa. 232 p. [ Links ] 23 Jensen, W., Salisbury, F., Ross, C .W. 1988. Botánica. Colombia: McGraw Hill. 762 p. [ Links ] 24 Kozai, T. 1991. Photoautotrophic Micropropagation. In vitro Cell Dev Biol 27: 47-51. [ Links ] 25 Lagos, M. I., Sáenz, F. A., Morales, M. A. 2008. Briófitos reófilos de tres quebradas del páramo de Mamapacha, Chinavita (Boyacá, Colombia). Acta Biológica Colombiana 13 (1): 143-160. [ Links ] 26 Linares, E., Uribe, J. 2002. Libro Rojo de briófitas de Colombia. Colombia: Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Ministerio del Medio Ambiente. 170 p. [ Links ] 27 Magdaleno, F. 2009. Restauración de bosques riparios de ríos y humedales. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. p. 26. [ Links ] 28 Mayz, J. 2004. Fijación biológica de nitrógeno. Revista UDO Agrícola 4 (1): 1-20. [ Links ] 29 Melgarejo, L. M. 2009. Bioprospección. Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. p. 12. [ Links ] 30 Molphe, E., Ramírez, R., Núñez H., Ochoa, N. 1999. Introducción al cultivo de tejidos vegetales. México: Universidad Autónoma de Aguascalientes. 179 p. [ Links ] 31 Montoya, L. 1991. Cultivo de tejidos vegetales. Colombia: Universidad Nacional de Colombia, Facultad de Ciencias Agropecuarias, Departamento de Agronomía. 77 p. [ Links ] 32 Mosquera, F. 2009. Pérdida, transformación y alteración de las comunidades vegetales altoandinas por la ganadería extensiva en el sector de la cuenca alta del río Caquetá, municipio de San Sebastián, departamento del Cauca. Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. pp. 75-76. [ Links ] 33 Murashige, T., Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473-97. [ Links ] 34 Novo, E. 2008. Lignificación en cultivos celulares de gimnospermas basales. Trabajo de grado de Doctorado en Ciencias Biológicas. Departamento de Biología Animal, Biología Vegetal y Ecología de la Universidad de A. Coruña en colaboración con el Departamento de Biología Vegetal (U.D. Fisiología Vegetal) de la Universidad de Murcia. pp. 43-44. [ Links ] 35 Oliver, M., Velten, J., Mishler, B. 2005. Desiccation tolerante in bryophytes: a reflection of the primitive strategy for plant survival in dehydrating habitats? Integrative & Comparative Biology 45: 788-789. [ Links ] 36 Orrego, O., Uribe, J. 2004. Hepáticas (Marchantiophyta) del departamento del Quindío, Colombia. Biota Colombiana 5 (2): 209-216. [ Links ] 37 Parra, J., Posada, J., Callejas, R. 1999. Guía ilustrada de los briofitos del Parque Arví Piedras blancas. Colombia: Universidad de Antioquia. 162 p. [ Links ] 38 Pedroza, J. 2008. Aplicaciones del cultivo de tejidos vegetales en condiciones in vitro. Bogotá, D. C.: Editorial Universidad Distrital FJC. 350 p. [ Links ] 39 Pedroza, J., Tupaz, W. 2008. Micropropagación de Ilex kunthiana Triana & Planchon (Aquifoliaceae), una especie de gran importancia en programas de revegetalización. Revista Colombiana de Biotecnología 10 (2): 72-84. [ Links ] 40 Pierik, R. 1990. Cultivo in vitro de las plantas superiores. España: Mundi-Prensa. 326 p. [ Links ] 41 Rivera, D., Linares, E. 2009. Biocostras y restauración de taludes y canteras del páramo de Chingaza. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. p. 117. [ Links ] 42 Rumary, C., Thorpe, T. 1984. Plantlet formation in black and white spruce. in vitro techniques. Can J For Res 14: 322-326. [ Links ] 43 Sabovljevic, M., Bijelovic, A., Dragicevic, A. 2003. in vitro culture of mosses: Alonia aloides, Ceratodon purpureus, Eurhynchium praelongum and Grimmia pulvinata. Turkish Journal of Botany 27: 441-446. [ Links ] 44 Salemaa, M., Mäkipää, R., Oksanen, J. 2008. Differences in the growth response of three bryophyte species to nitrogen. Environmental Pollution 152: 82-91. [ Links ] 45 Salisbury, F., Ross, C. 2000. Fisiología Vegetal: Desarrollo de las plantas y fisiología ambiental. Madrid: Thomson editores Paraninfo, S. A., pp. 573-574. [ Links ] 46 Sastre, I., Acosta, D., Santos, C. 2009. ¿Es la estructura del dosel de briófitos útil para predecir la diversidad de microinvertebrados de un bosque nublado tropical? Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. p. 20. [ Links ] 47 Scagel, R. 1973. El reino vegetal: los grupos de plantas y sus relaciones evolutivas. España: Omega. 659 p. [ Links ] 48 Schofield, W. 1996. Introduction to Bryology. Estados Unidos: Macmillan. 431 p. [ Links ] 49 Simijaca, D. F., Morales, M. E., Porras, S., Vargas, D. L. 2009. Plantas no vasculares como bioindicadores de contaminación atmosférica en Tunja (Boyacá, Colombia). Libro de Resúmenes: V Congreso Colombiano de Botánica. Asociación Colombiana de Botánica, San Juan de Pasto. p. 143. [ Links ] 50 Steel, R., Torrie, J. 1985. Bioestadística: Principios y procedimientos. Colombia: McGraw Hill. 622 p. [ Links ] 51 Strasburger, E., Noll, F., Schenck, H., Schimper, A. 2004. Tratado de botánica. 35 ed. España: Omega. 1134 p. [ Links ] 52 Suárez, A. 2005. Contribución al conocimiento de las hepáticas epífitas en un bosque nublado del municipio de Gachantivá (Boyacá). Bogotá: Trabajo de grado (Licenciado en Biología). Universidad Distrital Francisco José de Caldas. Facultad de Ciencias y Educación. Proyecto Curricular de Licenciatura en Biología. 118 p. [ Links ] 53 Taiz, L., Zeiger, E. 1998. Plant physiology. 2 ed. Sunderland: Sinanuer Associates, Inc. 560 p. [ Links ] 54Van Huylenbroeck, J. M. 1994. Influence of the light stress during the acclimatization of in vitro plantlets. En: Struik, P., Vredenberg, J., RenkemaJ., Parlevliet, J. (eds.). Plant production on the threshold of a new century. Netherlands: Kluwer Academic Publischers. pp. 451-453. [ Links ] 55 Van Huylenbroeck, J. M., Huygens, H., Deberg, P. C: 1995. Photoinhibition during acclimatization of micropropagated Spathiphyllum petite plantlets. in vitro Cell Dev Biol 31: 160-164. [ Links ] 56 Vasil, I. 1984. Cell culture and somatic cell genetic of plants. Volume I. Orlando, United States: Academic Press. 645 p. [ Links ] 57 Wickett, N., Goffinet, B. 2008. Origin and relationships of the myco-heterotrophic liverwort Cryptothallus mirabilis. Botanical Journal of the Linnean Society 156: 1-12. [ Links ] 58 Zamora, R. 2009. Las interacciones ecológicas como motores de la restauración de la biodiversidad en hábitats degradados. Libro de Resúmenes: I Congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. p. 25. [ Links ]