Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.12 no.1 Bogotá Jan./June 2010
ARTÍCULO DE INVESTIGACIÓN
Estandarización de un bioensayo y evaluación preliminar de tres formulaciones comerciales de Bacillus thuringiensis sobre Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae)
Standardising a bioassay for the preliminary evaluation of three commercial Bacillus thuringiensis formulations against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae)
Lorena Ramírez1 Natalia Ramírez1, Luz Stella Fuentes2,Jaime Jiménez2,Javier Hernández-Fernández 1
1Facultad de Ciencias, Carrera de Biología Ambiental, Genética, biología molecular y bioinformática, Laboratorio de Biología Molecular, Universidad Jorge Tadeo Lozano, Bogotá, D.C., Colombia. javier.hernandez@utadeo.edu.co Autor para correspondencia.
2Control Biológico de Plagas, Centro de Biosistemas, Universidad Jorge Tadeo Lozano, Chía, Cundinamarca, Colombia
Recibido: junio 18 de 2009 Aprobado: abril 15 de 2010
Resumen
La polilla del tomate (Tuta absoluta Meyrick; Lepidoptera: Gelechiidae) es una de las plagas más devastadoras del tomate en Colombia y países suramericanos, produciendo pérdidas de hasta el 100% en cultivos sin protección. En 2009, T. absoluta se detectó en España, Portugal y países del mediterráneo, además de Inglaterra, Bulgaria y Alemania. Para su control se utilizan insecticidas químicos que generan resistencia e impacto ambiental y de salud. La alternativa de utilizar biopesticidas contra esta plaga es de importancia creciente. En este estudio se evaluaron cinco métodos de bioensayo para medir adecuadamente la toxicidad sobre larvas de T. absoluta de tres productos comerciales: Dipel®, XenTary® y Turilav®, formulaciones a base de Bacillus thuringiensis (Bt). El método de Inmersión del folíolo, con el producto Dipel®, causó el 100% de mortalidad de larvas y 96% de supervivencia del testigo; este método presentó diferencias significativas al segundo (F=0,025, p>0,05) y cuarto (F=0,0018, p>0,05) día después de la aplicación (DDA). El método de Aspersión foliar por aerógrafo produjo 100% de mortalidad de larvas con Dipel® al segundo DDA (F=7,94x10-10, p> 0,05), y produjo diferencias significativas también al cuarto DDA (F=3,45x10-6, p>0,05). Los métodos Foliolos sumergidos y Medio de cultivo provocaron una alta mortalidad en el control por lo que fueron rechazados. El uso de Dipel®, XenTari® y Turilav® en concentración de 1,25 g/L causó entre 80-100% de mortalidad entre el segundo y octavo DDA en tres métodos evaluados válidos (1, 2, 3), además corrobora la actividad biológica de B. thuringiensis sobre este insecto plaga.
Palabras clave: Tuta absoluta, Bacillus thuringiensis, Dipel®, XenTary®, Turilav®
Abstract
The tomato moth (Tuta absoluta Meyrick; Lepidoptera: Gelechiidae) is one of the most devastating tomato pests in Colombia and South-American countries, producing losses of up to 100% in unprotected crops. T. absoluta was detected in Spain, Portugal and Mediterranean countries in 2009, as well as England, Bulgaria and Germany. Chemical insecticides are used for controlling it; however, they produce resistance and an environmental and human health impact. Finding an alternative to using biopesticides against this pest is becoming increasingly important. This study evaluated five bioassay methods measuring three commercial products toxicity on T. absoluta larvae: Dipel, XenTary and Turilav Bacillus thuringiensis (Bt) -based formulations. The leaf dipping bioassay method caused 100% larvae mortality with Dipel, the control group having 95% survival rate. The other products showed significant differences on the 2nd (F=0.025, p>0.05) and 4th (F=0.0018, p>0.05) days after application (DAA). The leaf spray airbrush method produced 100% larvae mortality with Dipel on the 2nd DAA, having significant differences from the other products tested on 2nd (DAA F=7.94 x 10-10, p>0.05 ), 4th (F=3.45x10-6, p>0.05 ) and 8th (F=1.07x10-5, p>0.05 ) DAA. Submerged leaflet and culture medium methods caused high mortality in controls and were thus rejected. A variation of the leaflet immersion method was standardised. The three commercial products produced high mortality in Lab conditions regarding T. absolute larvae control at 1.25 g/L concentration, thereby corroborating the biological activity of B. thuringiensis against this insect pest.
Key words: Tuta absoluta, Bacillus thuringiensis, Dipel, XenTary, Turilav (the last 3 being commercial insecticide brand names).
Introducción
La polilla del tomate Tuta absoluta (Meyrick) es considerada una de las principales plagas endémicas en plantas de tomate Solanum lycopersicum en los países sudamericanos (Suinaga et al., 1999; Torres et al., 2001), entre estos, Argentina, Bolivia, Brasil, Colombia, Chile, Ecuador, Paraguay, Perú, Uruguay y Venezuela, ocasionando daños que se estiman del orden del 40 al 100% en cultivos sin protección (Cely et al., 2006; Larraín, 1986; Giustolin et al., 2001). En Colombia, la plaga se ha diseminado por los principales departamentos productores de tomate: Cundinamarca, Santander, Valle, Caldas, Huila, Risaralda, Antioquia, Atlántico y La Guajira (Corporación Colombia Internacional, 2006).
Más recientemente, en 2006, T. absoluta fue reportada en Islas Canarias (España), en 2009, en Portugal y países del mediterráneo, además de Inglaterra, Bulgaria y Alemania, y 2010 en Israel (Tuta absoluta Information Network, 2010), convirtiéndose en una plaga cosmopolita de consecuencias inciertas.
Las larvas de T. absoluta infestan hojas, frutos y tallos de la planta. El daño es ocasionado cuando las larvas del insecto se alimentan del mesófilo de los folíolos haciendo minas y afectando la capacidad fotosintética de la planta y, de esta manera, reducen la producción. Sin embargo, las larvas también pueden penetrar el fruto causando cuantiosas pérdidas (García, 2002; Colombo and Berta, 2005; Corpoica, 2008).
Tuta absoluta presenta una metamorfosis completa y posee un alto potencial reproductivo produciendo entre 10-12 generaciones por año. El ciclo biológico se completa en 29-38 días dependiendo de las condiciones ambientales. El desarrollo se cumple hasta en 76 días a 14° C, 39 a 19,7 °C y 23 a 27,1 °C (Barrientos et al., 1997; Vélez, 1997). Los adultos son nocturnos y usualmente durante el día permanecen sobre los folíolos (Vélez, 1997).
Para obtener una producción sostenible de tomate, los agricultores recurren a prácticas de control químico severas, utilizando mezclas de insecticidas altamente tóxicos, en dosis y frecuencias elevadas. Lo anterior está causando graves alteraciones en el ambiente, en la salud humana y animal, y en la calidad de vida de los consumidores. Además, se estaría provocando un incremento de la tasa de aparición de insectos resistentes. En efecto, existen antecedentes acerca del desarrollo de resistencia a insecticidas clorados y fosforados, e incluso hacia los piretroides deltametrina y esfenvalerato (Salazar y Araya, 2001).
Desde principios del siglo pasado, el control biológico surgió como una nueva forma de controlar plagas, como alternativa al empleo de plaguicidas sintéticos, con la ventaja de ser efectivo y no causar daños al medioambiente. El microorganismo de mayor uso a nivel mundial es la bacteria Bacillus thuringiensis (Bt) una bacteria aeróbica, gram-positiva, formadora de esporas y productora de inclusiones paraesporales durante la fase de esporulación. Esta inclusión contiene proteínas insecticidas de cristal (ICPS) (Höfte y Whiteley, 1989), también denominadas d-endotoxinas, las cuales son codificadas por los genes cry (Schnepf et al., 1998). Bt es un patógeno de insectos cuya actividad es atribuida, amplia o completamente (dependiendo del insecto) a las ICPS, y su importancia radica en su toxicidad contra larvas de insectos-plaga de los órdenes lepidóptera, coleóptera, díptera, himenóptera, homóptera, ortóptera y malófaga, y aún en otros organismos tales como platelmintos, nemátodos, protozoos y ácaros (Hofte y Whiteley, 1989; Feitelson et al., 1992, 1993; Bravo et al., 1998; Su et al., 2007). Incluso han mostrado actividad sobre células cancerígenas (Mizuki et al., 2000; Akio et al., 2004).
El desarrollo de las formulaciones insecticidas elaboradas a base de la bacteria Bt es una tecnología centenaria que ha recibido un fuerte impacto en décadas recientes. La mezcla de esporas y cristales, que es el principio activo de estas preparaciones comerciales, ha sido objeto de estudios constantes, y en ellos se destaca la búsqueda de cepas cada vez más potentes o mejoradas (Rosas-García, 2008). Los productos de este microorganismo presentan algunas características importantes que justifican su uso, como la ausencia de toxicidad sobre los seres humanos, en enemigos naturales de diversas plagas, en otros vertebrados y en plantas (Rosas-García, 2008).
El éxito de estas formulaciones se ha visto reflejado en la gran variedad de productos comerciales que existen, solo las preparaciones por aspersión comprenden aproximadamente el 2% del mercado global de los insecticidas, y aunque no ha sido fácil su introducción en el mundo de la agricultura, la aplicación de los cultivos con formulaciones tradicionales de Bt está constituyendo la estrategia de elección de los agricultores orgánicos, ya que se utilizan para una producción agrícola más sana, limpia y de mayor calidad.
La presente investigación se realizó con el fin de estandarizar la cría, realizar un bioensayo sobre larvas de la polilla del tomate T. absoluta y determinar la eficacia de tres productos comerciales cuyo ingrediente activo son esporas y cristales de cepas de B. thuringiensis: Dipel®, XenTari® y Turilav®.
Materiales y métodos
Plantas utilizadas: se utilizaron semillas de tomate variedad Milano Tropic (Impulsemillas). Se sembraron en bandejas con celdillas de 15 cm de profundidad conteniendo turba (material orgánico), se colocaron por cinco días en cuarto oscuro a una temperatura de 25°C, hasta la germinación. Posteriormente, se dejaron a temperatura ambiente en el semillero por tres semanas. Las plántulas desarrolladas se trasplantaron a materas de 18 cm de alto.
Cría del insecto plaga T. absoluta: se recolectaron polillas adultas de T. absoluta en Chía, Cundinamarca, Colombia (4°52´00.01´´ N, 74°03´57.31´´O) en cultivos de tomate mantenidos en condiciones de invernadero (temperatura 27±13°C, HR 40-90%). Se estableció una cría en jaulas de 80 x 60 x 60 cm, recubiertas con velo suizo. En una jaula se colocaron tres materas con plantas de tomate con aproximadamente 60 adultos en proporción sexual 1:1. Las plantas con las posturas de huevos se trasladaron a otra jaula. Después de la eclosión, las larvas se clasificaron de acuerdo con su ciclo metamórfico en 1er, 2do, 3er y 4to instar. Las larvas de 4to instar se utilizaron para infestar nuevas plantas de tomate dentro de jaulas, y de esta forma mantener la cría (González, 1989). Plantas de mes y medio de edad se infestaron con polillas adultas y después de dos días se encontraron infestadas únicamente en los folíolos superiores, con una buena cantidad de huevos por planta.
Las jaulas establecidas para la cría del insecto estuvieron sometidas a temperaturas variables, altas durante el día (27° C) y bajas durante la noche (13° C), condición que prolongó y produjo variabilidad en el ciclo reproductivo de las polillas, llegando a estar entre 50-60 días, resultado muy similar al descrito en anteriores estudios (Vélez, 1997; Estay, 2000; Fernández y Montagne, 1990; González, 1989; López, 1991). Para la realización de los bioensayos se utilizaron larvas de 2do instar.
Productos biológicos utilizados: se utilizaron tres productos comerciales Dipel®, XenTari® y Turilav®. Los dos primeros son producidos por la casa comercial Valent BioSciences Corporation (USA), constituidos por las cepas de Bt var. kurstaki y aizawai respectivamente. Turilav® es producido por la empresa Laverlam S.A (Cali, Colombia), constituido por la cepa Bt var. kurstaki.
Experimentos para la estandarización del bioensayo
Los tres insecticidas biológicos comerciales se prepararon resuspendiendo 1,25 g de producto en un litro de agua de acuerdo con la recomendación de la casa comercial fabricante (Valent BioSciences Corporation, USA). En los experimentos se utilizaron estas diluciones en un diseño completamente al azar. En los métodos 1 a 4 se utilizaron cuatro réplicas con cinco larvas cada una y en el 5 tres réplicas con 21 larvas. La mortalidad se evaluó cada 24 h por ocho días utilizando un estereoscopio Advanced Optical, y se corrigió utilizando la fórmula de Henderson y Tilton (1955). La mortalidad corregida para los tres productos se sometió a un análisis de varianza (Anova) y prueba de comparación de medias de Tukey (P<0,05).
Se evaluaron cinco métodos para la estandarización del bioensayo, estos fueron:
1. Inmersión de folíolos: los folíolos de tomate fenológicamente similares se lavaron, secaron y sumergieron por 10 min en las diluciones insecticidas por separado. Luego se dejaron secar a temperatura ambiente y se introdujeron en cajas de Petri sobre papel absorbente húmedo. Para mantener hidratado el folíolo se le colocó un algodón húmedo. Después, los folíolos se infestaron con cinco larvas de 2do instar utilizando un pincel de pelo de marta. Se utilizó un grupo control sin tratamiento (testigo absoluto) (adaptado de Niedman y Meza-Basso, 2006).
2. Aspersión foliar por aerógrafo: este experimento se realizó igual que el experimento anterior variando únicamente la forma de aplicación de los bioinsecticidas sobre los folíolos en los diferentes tratamientos. Se utilizó un aerógrafo con el que se aplicó 1 ml de la dilución de los tres bioinsecticidas por separado sobre el haz y el envés de los folíolos de tomate a una distancia de 15 cm, procurando cubrir toda la superficie.
3. Frascos con folíolos de tomate con pecíolo sumergido: a manera de florero se colocaron folíolos de plantas de tomate (folíolos con un tallo pequeño) en el interior de otro frasco plástico pequeño, el cual fue cubierto con un dedo de guante quirúrgico para evitar que las larvas cayeran al agua. Los productos biológicos por evaluar se sirvieron utilizando el método de Inmersión de las hojas. Luego, los frascos se colocaron dentro de otro recipiente plástico y se sellaron con tapa.
4. Medio de cultivo extracto de folíolos de tomate: se preparó una dieta artificial utilizando 50 g de folíolos de tomate; 0,65 g agar; 1,65 g ácido ascórbico; 1,65 g ácido cítrico y 0,25 g Dipel® y se disolvieron en 50 ml de agua destilada. Esta mezcla se homogeneizó en una licuadora Osterizer y luego se sirvió en cajas de Petri. Los demás procedimientos se realizaron igual que el de Inmersión de folíolos. Se evaluó únicamente la dilución del producto comercial Dipel®.
5. Recipientes plásticos como unidades experimentales: este método es una variación del método 1, Inmersión del folíolo, y se propuso para homogeneizar el montaje del bioensayo, ya que al hacerlo por separado en cajas de Petri hay mayor manipulación y, por tanto, mayor probabilidad de error. Se utilizaron recipientes plásticos de 28 x 16 x 4 cm, el sistema de aplicación de los productos empleados fue inmersión. Después los folíolos se dejaron secar a temperatura ambiente y con un pincel de pelo de marta se colocaron 21 larvas por tratamiento. Se utilizaron tres repeticiones por tratamiento incluyendo al testigo absoluto (control).
Resultados y discusión
Estandarización del sistema de bioensayo
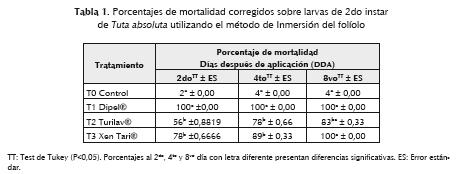
La evaluación del método 1, Inmersión de folíolos, produjo el efecto esperado sobre las larvas de 2do instar de Tuta absoluta, observándose alta toxicidad al segundo día después de la aplicación (DDA) con un 100% de mortalidad en el tratamiento T1 (producto comercial Dipel®). De igual manera, en los tratamientos T2 (Turilav ®) y T3 (XenTari®) se observó un incremento de la mortalidad a partir del segundo DDA; sin embargo, solo hasta el octavo DDA se evidenció un 100% de mortalidad de las larvas en el T3 y 83% en T2 (Tabla 1). El análisis estadístico presentó diferencias significativas entre los tratamientos evaluados al segundo (F=0,0249, p>0,05), cuarto (F=0,001768, p>0,05) y octavo DDA (F=0,00007456, p>0,05); el test de Tukey presentó diferencias entre T1 y T2-T3 al segundo y cuarto DDA; al octavo DDA no se presentaron diferencias entre T1-T3, pero sí de estos dos con T2 (Tabla 1).
El testigo absoluto (T0) produjo 4% de mortalidad, menor al máximo permitido para bioensayos (Beegle, 1990). En este experimento los biopesticidas utilizados se distribuyeron homogéneamente en los folíolos, lo que permite confiabilidad de los resultados. Sin embargo, la cantidad de unidades experimentales contando las repeticiones es grande, lo que redunda en alta manipulación de las larvas, mayor tiempo empleado y costo del bioensayo. En el método 2, Aspersión foliar por aerógrafo, los tratamientos evaluados mostraron toxicidad sobre larvas de Tuta absoluta a partir del 2do DDA, con un 100% de mortalidad en T1 (Tabla 2). Sin embargo, aunque los tratamientos T2 y T3 incrementaron su eficacia después de la aplicación, solo hasta el 8vo DDA causaron 85 y 69% de mortalidad respectivamente (Tabla 2). El análisis estadístico presentó diferencias significativas entre el T1 y los demás tratamientos al 2do DDA (F=7,94 x 10-10 p>0,05), diferencia corroborada con el test de Tukey, que evidencia diferencias cercanas entre los promedios de supervivencia de los testigos T2 y T3 en comparación con el T1 (Tabla 2). Igualmente, al 4to DDA se presentaron diferencias significativas entre el T1 y T2-T3 (F=3,455 x 10-6 p>0,05) (Tabla 2). Al 8vo DDA se presentaron diferencias entre T1 y T3 (F=1,073 x 10-5 p>0,05) pero no con el T2 (Tabla 2). El T0 (grupo control) causó el 3% de mortalidad, lo que valida este bioensayo (Tabla 2). Sin embargo, en este experimento la inoculación de los biopesticidas sobre los folíolos no fue homogénea. Se registró el peso de las hojas antes y después de la aspersión obteniéndose los pesos promedio de producto asperjado en cada uno de los experimentos, estos fueron: 1.754, 1.143 y 0.479 ml para T1, T2 y T3 respectivamente. Por esto, tal vez las diferencias en mortalidad presentadas con el método 1, Inmersión de folíolos, que para el producto XenTari® (T3) son altas, ya que redujo la mortalidad en 31%. Para los otros dos bioinsecticidas la mortalidad no presentó diferencias. El método 3, Frascos con foliolos sumergidos de plantas de tomate, causó el 100, 29 y 33% de mortalidad de las larvas, en T1, T2 y T3 respectivamente. El control presentó el 67% de mortalidad, lo que invalida este sistema de bioensayo (Figura 1). En este experimento el modelo de florero no es estable para las larvas lo que ocasiona una mortalidad alta en el control. Sin embargo, en T2 y T3 se produjo una disminución en la mortalidad con respecto a los dos métodos anteriores, debida tal vez a la posición vertical de los folíolos. T1 causó una mortalidad del 25% al 1er DDA, mientras que T2 y T3 no causaron mortalidad de las larvas. Al 3er DDA, T1 causó 71% de mortalidad, respecto al 9 y 14% de T2 y T3, respectivamente. Aunque este experimento no fue válido, sí nos permite proponer la hipótesis de que el producto comercial Dipel® (T1) es el bioinsecticida menos variable y el más rápido en su acción tóxica sobre larvas de T. absoluta, entre los tres productos evaluados El método 4, Medio de cultivo extracto de hojas de tomate, causó una mortalidad del 100% en todos los grupos (T1, T2 y T3, incluyendo el grupo control T0) presentando adicionalmente contaminación, resultado que invalidó este método. Al parecer, el medio de cultivo formulado presentó una concentración alta de productos de folíolos de tomate y esto pudo generar la mortalidad del 100% de las larvas. En algunas especies de tomate, como L. hirsutum y L. glabratum, han sido reportadas altas concentraciones de alfa tomatina, un alcaloide tóxico para varios lepidópteros entre ellos, T. absoluta (Giustulin et al., 2001). Es posible que los folíolos de L. esculentun utilizados en este estudio produzcan este alcaloide en tan bajas concentraciones que no alcanzan normalmente a presentar toxicidad sobre larvas de T. absoluta, pero posiblemente sí al concentrarlas en el medio de cultivo, que se preparó al 100% de concentración de hojas de tomate (50 g/hojas en 50 ml de H2O). El análisis de estos cuatro métodos permitió concluir que aquellos aceptables para realizar los bioensayos con T. absoluta fueron el 1, Inmersión de folíolos, y el 2, Aspersión con aerógrafo, ya que la mortalidad del testigo fue inferior al 10% por lo cual estos dos métodos se consideran adecuados (Beegle, 1990). Sin embargo, el método 1 fue el que presentó resultados claros, confiables y reproducibles. Los métodos 3 y 4, con mortalidades del testigo del 33 y 100%, fueron invalidados (Beegle, 1990). Se decidió optimizar el método estandarizado, para ello se utilizó el método 5, Recipientes plásticos como unidades experimentales, con el fin de disminuir la manipulación de las larvas y el tiempo empleado para la lectura del bioensayo. Las larvas de T. absoluta son tremendamente susceptibles a la manipulación y a los cambios físicos y ambientales a los que son sometidas. Este método mostró resultados muy parecidos al método 1, Inmersión de folíolos, 3% de mortalidad del control y 100, 87 y 100% de mortalidad de larvas de T. absoluta en los T1, T2 y T3 respectivamente (Figura 2). El análisis estadístico presentó diferencias significativas entre los tratamientos evaluados al 2do (F=0,00217, p>0,05), y 4to DDA (F=0,001652, p>0,05), el test de Tukey mostró diferencias significativas entre T1 y T2-T3 al segundo y cuarto DDA; al octavo DDA no se presentaron diferencias entre ninguno de los tratamientos. Igualmente, el producto comercial Dipel® (T1) mostró más rapidez en su actividad tóxica, ya que en el segundo día causó el 100% de mortalidad de las larvas, contra el 75% de Turilav ® (T2) y 52% de XenTari® (T3) (Figura 2). Actividad de los formulados comerciales La supervivencia en el testigo es el parámetro fundamental para tener en cuenta en un bioensayo en condiciones controladas tendiente a evaluar uno o varios factores de mortalidad sobre un insecto específico. Si la mortalidad en el testigo alcanza un nivel superior al 10% límite fijado como aceptable, los resultados no pueden tenerse en cuenta (Beegle, 1990). De los resultados obtenidos se desprende la posibilidad de emplear los formulados basados en cepas de Bt evaluados para el control de T. absoluta. Al parecer, el complejo espora cristal base de los biopesticidas utilizados es capaz de provocar la detención de la alimentación de las larvas de la polilla del tomate, produciendo la muerte y por tanto su control. Es posible proponer la hipótesis de que T. absoluta posee receptores de membrana en el intestino medio capaces de reconocer una o varias proteínas Cry de Bt (Gill et al., 1992; De Maagd et al., 2001). Estudios previos ya habían descrito actividad de Bt sobre esta polilla (Theoduluz., 1997; Giustolin et al., 2001; Theoduluz et al., 2003; Niedmann y Meza-Basso, 2006). El producto comercial Dipel® tiene como ingrediente activo la misma cepa de Bt que el producto Turilav®, la subespecie kurstaki (Bt.k) serotipo H3a3b. Sin embargo, parece existir diferencia en la efectividad de los dos productos. De acuerdo con los resultados preliminares presentados, Dipel® es más rápido en su acción tóxica sobre las larvas de T. absoluta. Solo en cuanto al porcentaje de mortalidad es un poco más bajo Turilav® (83%). La cepa de Bt del producto Turilav® es Bt var. aizawai (Bt.a) serotipo H7 que expresa las toxinas Cry1Ba, Cry1Ca y Cry1Da diferentes a las expresadas por Bt.k,que son Cry1Aa, Cry1Ab, Cry1Ac, Cry1Ia, Cry2Aa y Cry2Ab; al parecer, varias proteínas tienen actividad sobre larvas de T. absoluta, ya que los tres formulados evaluados tienen actividad tóxica, pero difieren entre las proteínas que expresan. Niedmann y Meza-Basso (2006) describieron actividad tóxica sobre larvas de T. absoluta utilizando aislados nativos chilenos que expresaban Cry1Ac. Theodoluz et al. (2003), realizando clonación de genes de Bt y produciendo la proteína recombinante Cry1Ab describieron actividad de esta toxina sobre este mismo insecto plaga. Estas evidencias experimentales están de acuerdo con los resultados obtenidos en este estudio, ya que Dipel® y Xen- Tari® contienen estas dos toxinas y mostraron un control potencial de las larvas de T. absoluta. Turilav® también mostró un control significativo sobre este insecto plaga. En consecuencia, podemos concluir que los productos comerciales de Bt evaluados controlan larvas de 2do instar de T. absoluta en condiciones controladas de laboratorio. El uso de Dipel®, XenTari® y Turilav® en concentración de 1,25 g/L causaron 80-100% de mortalidad entre los 2-8 DDA en tres métodos evaluados válidos (1, 2, 5), además, corrobora la actividad biológica de B. thuringiensis sobre este insecto plaga. Agradecimientos Los autores agradecen a la Dirección de Investigaciones de la Universidad Jorge Tadeo Lozano por apoyar el desarrollo completo del presente estudio. Referencias bibliográficas 1 Akio, I., Yasuyuki, S., Sakae, K., Yoshitomo, K., Kyoko, K., Kenjiro et al. 2004. A Bacillus thuringiensis Crystal Protein with Selective Cytocidal Action to Human Cells. The J of Biol Chemistry 272: 21282-21286. [ Links ] 2 Barrientos, Z., Apablaza, H., Norero, S., Estay, P. 1997. Threshold temperature and thermal constant for development of the South American tomato moth, Tuta absoluta (Lepidoptera, Gelechiidae). Ciencia e Investigacion Agraria 25, 133-137 (in Spanish). [ Links ] 3 Beegle, C. 1990. Bioassays methods for quantification of Bacillus thuringiensis delta–endotoxin. En: Analytical chemistry of Bacillus thuringiensis. American Chemical Society. USA. [ Links ] 4 Bravo, A., Sarabia, S., López, L., Ontiveros, H., Abarca, C., Ortiz et al. 1998. Characterization of cry genes in a Mexican Bacillus thuringiensis strain collection. Appl and Environ Microbiol 64 (12): 4965-4972. [ Links ] 5 Cely, L., Cantor, F., Rodríguez, D., Cure, J. 2006. Niveles de daños ocasionados por diferentes densidades de Tuta absoluta (Lepidóptera: Gelechiidae) en tomate bajo invernadero. Resúmenes Socolen Reencuentro con la Entomología en el Eje Cafetero. XXXIII. Manizales 26, 27 y 28 de julio. Congreso de Entomología. [ Links ] 6 Colombo, M., Berta, D. 2005. Los EJEMPLARES TIPO de Eumeninae (Hymenoptera: Vespidae) depositados en la colección del Instituto Fundación Miguel Lillo (IFML), Argentina. Rev Soc Entomol Argent 63 (3): 23-35. [ Links ] 7 Corpoica (Corporación Colombiana de Investigación Agropecuaria) 2008. Librería virtual agropecuaria. Jorge Jaramillo: Tomate Bajo invernadero. Julio 2008. Bogotá, Colombia. http:// intranet.corpoica.org.co Fecha de consulta: 24 de julio de 2009. [ Links ] 8 Corporación Colombia Internacional. 2006. Ministerio de Agricultura, acerca de certificación agroecológica. Buenas prácticas agrícolas. http://certificacion74.blogspot.com/2006/04/qu-es-lacorporacin-colombia.html. Fecha de consulta: 20 de julio de 2008. [ Links ] 9 De Maagd, R., Bravo, A., Crickmore, N. 2001. How Bacillus thuringiensis has evolved specific toxin to colonize the insect world. Rev Trend Genet 17: 193-199. [ Links ] 10 Estay, P. 2000. Polilla del Tomate Tuta absoluta (Meyrick). Informativo La Platina 9:1-4. [ Links ] 11 Feiltelson, J., Payne, J., Kim, L. 1992. Bacillus thuringiensis: Insects and beyond Biotechnol 10: 271-275. [ Links ] 12 Feiltelson, J. 1993. The Bacillus thuringiensis family tree. Advanced engineered pesticides. 63-71. [ Links ] 13 Fernández, S., Montagne, A. 1990. Biología del Tomate, Scrobipalpula absoluta (Meyrick) (Lepidoptera: Gelechiidae). Boletín de Entomología Venezolana 5 (12): 89-99. [ Links ] 14 García, F. 2002. Manejo biológico de plagas en tomate. En control biológico, componente fundamental del manejo integrado de plagas en una agricultura sostenible. Memorias del I curso taller internacional. Programa nacional Manejo integrado de plagas Corpoica. [ Links ] 15 Gill, S., Nowles, E., Pietrantonio, P. 1992. The mode of action of Bacillus thuringiensis endotoxins. Annu Rev Entomol 37: 615-36. [ Links ] 16 Giustolin, T., Vendramin, J., Alves, S., Viera, A., Pereira, R. 2001. Susceptibility of Tuta absoluta (Meyrick) (Lep., Gelechiidae) reared on two species of Lycopersicon to Bacillus thuringiensis var. kurstarki. J of Appl Entomol 125: 551-556. [ Links ] 17 González, R. 1989. Insectos y ácaros de importancia cuarentenaria en Chile. Santiago, Ograma S.A. 310. [ Links ] 18 Henderson, C., Tilton, E. 1955. Tests with acaricides against the brown wheat mite. J Econ Entomol 48: 157-161. [ Links ] 19 Höfte, H., Whiteley, H. 1989. Insecticidal cristal proteins of Bacillus thuringiensis. Microbiol Review 53: 242- 255. [ Links ] 20 Larraín, P. 1986. Plagas del tomate. Investigación y Progreso Agropecuario 39. p. 30-35. [ Links ] 21 López, E. 1991. Polilla del tomate: problema crítico para la rentabilidad del cultivo de verano. Empresa y Avance Agrícola 1 (5): 6-7. [ Links ] 22 Mizuki, E., Park, Y., Saitoh, H., Yamashita, S., Akao, T., Higuchi, K., Ohba, M. 2000. Parasporin, a human leukemic cell-recognizing parasporal protein of Bacillus thuringiensis. Clinical and Diagnostic Laboratory Immunology 7: 625-634. [ Links ] 23 Niedmann, L., Meza-Basso, L. 2006. Evaluación de cepas nativas de Bacillus thuringiensis como alternativa de manejo integrado de la polilla del tomate Tuta absoluta (Meyrick); Lepidopetera: Gelechiidae en Chile. Agricultura Técnica 66 (3): 235-246. [ Links ] 24 Rosas-García, N. 2008. Avances en el desarrollo de formulaciones insecticidas a base de Bacillus thuringiensis.Revista Colombiana de Biotecnología 10: 49-63. [ Links ] 25 Salazar, E., Araya, J. 2001. Respuesta de la polilla del tomate, Tuta absoluta (Meyrick), a insecticidas en Arica. Agricultura Técnica 61 (4): 429-435. [ Links ] 26 Schnepf, E., Crickmore, N., van Rie, J., Lereclus, D., Baum, J., Feitelson, J. 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol Mol Biol Review 62: 775-806. [ Links ] 27 Su, X., Shu, C., Zhang, J., Huang, D., Tan, J., Song, F. 2007. Identification and Distribution of Bacillus thuringiensis Isolates from Primeval Forests in Yunnan and Hainan Provinces and Northeast Region of China. Agricultural Sciences in China 6: 1343-1351. [ Links ] 28Suinaga, F., Picanço, M., Jham, G., Brommonschenkel, H. 1999. Causas químicas de resistência de Lycopersicon peruvianum (L.) a Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Anais da Sociedade Entomológica do Brasil 28: 313-321. [ Links ] 29 Theoduluz, C., Roman, P., Bravo, J., Padilla, C., Vásquez, C., Meza-Cepeda, L., Meza-Basso, L. 1997. Relative toxicity of native Chilean Bacillus thuringiensis strain against Scrobiplapuloides absoluta (Lepidoptera:Gelechiidae). J Appl Microbiol 82: 462-468. [ Links ] 30 Theoduluz, C., Vega, A., Salazar, M., González, E., Meza- Basso, L. 2003. Expression of a Bacillus thuringiensis d-endotoxin cry1Ab gene in Bacillus subtilis and Bacillus licheniformis strains that naturally colonize the phylloplane of tomato plants (Lycopersicon esculentum, Mills). J Appl Microbiol 94: 375-381. [ Links ] 31 Torres, J., Faria, C., Evangelista, J., Pratissoli, D. 2001. Within plant distribution of the leaf miner Tuta absoluta (Meyrick) immatures in processing tomatoes, with notes on plant phenology. International J of Pest Management 47: 173-178. [ Links ] 32 Tuta absoluta Information network. http://www.tutaabsoluta.com. com. (Revisión: marzo 10 de 2010). [ Links ] 33 Vélez, R. 1997. Plagas agrícolas de impacto económico en Colombia: bionomía y manejo integrado. Editorial Universidad de Antioquia, Medellín. p. 379-385. [ Links ]