Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.12 n.1 Bogotá jan./jun. 2010
ARTÍCULO DE INVESTIGACIÓN
Estudio de diversidad de accesiones de guayabo (Psidium guajava L.) mediante el marcador molecular ISTR
Studying the diversity of guava (Psidium guajava L.) accessions using ISTR molecular markers
Narciso N. Rodríguez Medina1 , Juliette Valdés-Infante Herrero2 , Miguel Angel Keb Llanes3 , Aileen OConnor Sánchez4 , Wolfgang Rohde5
1Biólogo, Doctor en Ciencias Biológicas, Investigador Titular, Instituto de Investigaciones en Fruticultura Tropical. La Habana. Cuba. isabel.garcia@infomed.sld.cu
2Bióloga, Doctora en Ciencias Biológicas, Investigador Agregado, Instituto de Investigaciones en Fruticultura Tropical. La Habana. Cuba. mejoramiento@iift.cu. Autor para correspondencia.
3Técnico, Centro de Investigaciones Científicas de Yucatán. Mérida, Yuc., México. mkeb@latinmail.com
4Doctora en Ciencias Biológicas y Biotecnología de plantas, Investigador asociado C, Centro de Investigaciones Científicas de Yucatán. Mérida, Yuc., México. aileen@cicy.mx
5Ingeniero Químico, Profesor Doctor Max-Planck Institut für Züchtungsorschung (MPIZ), Köln, Germany. rohde@mpiz-koeln.mpg.de
Recibido: marzo 2 de 2010 Aprobado: junio 23 de 2010
Resumen
Los estudios de variabilidad genética resultan útiles para el manejo racional del material, tanto para su conservación como para el mejoramiento. La Repetición de Secuencias Inversas Marcadas (ISTR) es una técnica basada en la PCR que permite el estudio de la diversidad genética de individuos y poblaciones; identificación de cultivares, entre otras aplicaciones. En este sentido, el objetivo del presente trabajo fue estudiar la diversidad de accesiones de guayabo empleando este marcador molecular. Para el análisis de los datos se generó una matriz de ausencia-presencia de las bandas polimórficas, a partir de la cual se desarrolló un análisis de agrupamiento basado en el coeficiente de Jaccard y el método UPGMA para la construcción del dendrograma con el programa NTSYS-pc. La evaluación de los genotipos de guayabo con el marcador ISTR generó un total de 52 bandas polimórficas, las cuales permitieron diferenciar todos los materiales evaluados, corroborando la utilidad de esta técnica para la identificación de accesiones en la especie. El análisis de agrupamiento permitió evidenciar la formación de cuatro grupos de diversidad definidos y la presencia de cuatro accesiones externas. Las diferencias encontradas en el agrupamiento de las accesiones por ISTR respecto a las obtenidas previamente por AFLP y SSR sugieren que el elegir la técnica más apropiada para determinados estudios puede resultar un proceso difícil. Los resultados discutidos en este trabajo indican la necesidad de realizar estudios integrados en el banco de germoplasma de este cultivo para lograr un manejo más racional de la base genética del mismo.
Palabras clave: identificación de genotipos, parentesco, polimorfismo.
Abstract
Genetic variability studies can be useful for the rational management of material as well as its conservation and improvement. Inverse sequence-tagged repeat (ISTR) is a PCR-based technique which allows individuals and populations genetic diversity to be studied, cultivars to be identified, etc. The present work was thus aimed at using this molecular marker to study guava accession diversity. An absence-presence matrix for polymorphic bands was generated for data analysis; cluster analysis was developed from this, based on Jaccards coefficient and the UPGMA method for dendrogram construction using NTSYS-pc software. Guava genotype evaluation with the ISTR marker generated 52 polymorphic bands, allowing all the materials being evaluated to be differentiated, thereby corroborating this techniques use for identifying accessions in the specie. Cluster analysis revealed the formation of four defined diversity groups and the presence of four external accessions. The differences encountered for accession clustering with ISTR regarding those previously obtained by AFLP and SSR suggested that choosing the most appropriate technique for determinate studies can be difficult. The results indicated the need for carrying out integrated studies on this crops germplasm bank to ensure more suitable management.
Key words: Genotype identification, relatedness, polymorphism.
Introducción
El guayabo (Psidium guajava L.) ha adquirido recientemente una mayor importancia en las áreas tropicales y subtropicales del mundo, debido principalmente a su alto contenido de ácido ascórbico y por lo rentable de su cultivo (Coelho de Lima et al., 2002; Vasco et al., 2003; Sanabria et al., 2005).
Los estudios de variabilidad genética resultan útiles para el manejo racional del material, tanto para su conservación como para el mejoramiento genético de la especie (Marcucci et al., 2001; Frankham et al., 2003; Tapia et al., 2005). En este sentido, en la literatura hay varias referencias acerca del uso de marcadores de ADN para algunos de los miembros de la familia Myrtaceae a la cual pertenece el guayabo (Aitken et al., 1998; Dettori y Palombi, 2000; Rossetto et al., 2000). Por otra parte, se han alcanzado importantes resultados mediante el empleo de marcadores del tipo Polimorfismo del ADN Amplificado al Azar (RAPD, de sus siglas en inglés), Polimorfismo de la Longitud de Fragmentos Amplificados (AFLP, de sus siglas en inglés) y Secuencias Simples Repetidas (SSR, de sus siglas en inglés) en colecciones del germoplasma de este frutal en México, Colombia, Venezuela, India y Cuba (Hernández et al., 2003; Reveles et al., 2003; Rueda, 2003; Valdés-Infante et al., 2003; Rodríguez et al., 2004; Rodríguez et al., 2005; Sharma et al., 2005; Sanabria et al., 2006; Hernández-Delgado et al., 2007; Aranguren y Fermin, 2007).
La Repetición de Secuencias Inversas Marcadas (ISTR) es una técnica basada en la Reacción en Cadena de la Polimerasa (PCR, de sus siglas en inglés) que utiliza secuencias repetitivas denominadas retrotransposones como cebadores. Estas secuencias se encuentran ampliamente distribuidas por todo el genoma de eucariontes y permiten el estudio de la diversidad genética de individuos y poblaciones, la identificación de cultivares, entre otras aplicaciones (Rohde, 1996; Valdés-Infante y Rodríguez, 2003; Capote, 2007). En el caso específico del guayabo, sólo existe la referencia del uso de este marcador molecular para el estudio de parentales y descendientes del cruce Enana Roja Cubana x Bangkok (Rodríguez et al., 2003). El objetivo del presente trabajo fue estudiar la diversidad de accesiones de guayabo empleando la técnica ISTR.
Materiales y métodos
Material vegetal
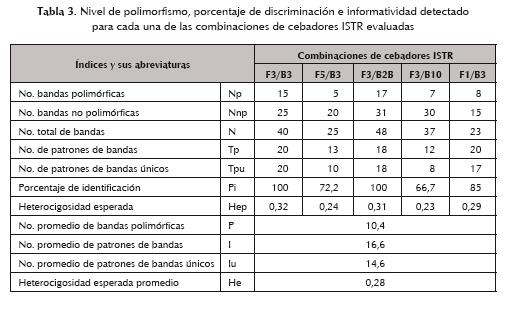
El material vegetal utilizado para la caracterización morfoagronómica (tabla 1) forma parte de la colección de guayabo (Psidium guajava L.) de la Unidad Científico-Tecnológica de Base (UCTB) de Alquízar, provincia La Habana, perteneciente al Instituto de Investigaciones en Fruticultura Tropical (IIFT) de Cuba. Las accesiones provienen de diferentes fuentes: plantas introducidas, prospecciones realizadas en distintas localidades del país y selecciones producidas por polinización abierta, fundamentalmente de los cultivares N6, Suprema Roja, Indian Pink y Perú Roja.
La colección se encuentra ubicada en los 22° 47 de latitud norte y los 82° 31 de longitud oeste a 11 m sobre el nivel del mar y con una topografía llana de pendiente 0. Los suelos sobre los que se asienta corresponden al tipo ferralítico rojo compactado y ferralítico rojo hidratado, y tienen un pH entre 5,5 y 6,5 (Rodríguez et al., 1999). La misma fue confeccionada siguiendo un diseño completamente aleatorizado, que incluye dos réplicas para cada accesión. Aislamiento y purificación del ADN genómico Se extrajo el ADN genómico procedente de 5,0 g de material foliar, a través de una modificación sugerida por Ramírez et al. (2004) al tampón de extracción CTAB (Polovinilpirrolidona al 2%, 25 mM de ascorbato de sodio, 25 mM de bisulfito de sodio y 10 mM de dietilditocarbamato) del protocolo original descrito por Doyle y Doyle (1990). Posteriormente, se purificaron con columnas NucleoSpin (Machery y Nagel, 2002) para ADN de alto peso molecular. La concentración y calidad del ADN precipitado fue determinada por electroforesis en gel de agarosa al 0,7% y comparada con el marcador de concentración 1 kb (GIBCOBRL, Netherlands). Análisis con el marcador molecular ISTR Se utilizó el material vegetal referido en la tabla 1 para la caracterización de la diversidad a través de este tipo de marcador de ADN. Se evaluaron cinco combinaciones de cebadores: F5/B3; F3/B2B; F3/B10; F3/B3 y F1/B3 (tabla 2), siguiendo el protocolo descrito en la literatura por Rohde (1996). La reacción en cadena de la polimerasa (PCR) fue desarrollada en un volumen final de 20 µL que contenían 10-50 ng de ADN; 0,2 mM de dNTP; 2,5 mM de MgCl2; buffer de PCR 1X (GIBCO-BRL, Groningen, Netherlands); 10 pM de cada cebador de AFLP y 1U de Taq ADN polimerasa (GIBCO-BRL). El programa empleado es el siguiente: 95 °C, 3 min; 95 °C, 30 s; 45 °C, 30 s; 72 °C, 2 min, con repetición de 40 ciclos del paso 2 al 4 y un paso final de extensión de 72 °C, 10 min. Los productos de la PCR generados fueron desnaturalizados por calentamiento durante 5 min a 94° C en presencia de formamida (98% formamida desionizada, 10 mM EDTA, 0,025% bromofenol azul y 0,025% xilencianol). Se aplicaron alícuotas de 3 µL en un gel desnaturalizante de poliacrilamida al 4%, empleando el tampón de corrida TBE 1X para la electroforesis, la cual se llevó a cabo a pH 8,9 y 40 W. Después de la corrida, los geles fueron revelados por tinción con sales de plata, según el protocolo referido por Valadez y Günter (2000). Análisis de los datos Para el análisis de los datos se generó una matriz de ausencia (0)-presencia (1) de las bandas polimórficas. A partir de esta matriz se calcularon los siguientes parámetros de acuerdo con lo sugerido por Belaj et al. (2003): 1. Número de unidades de ensayo (u: producto de la amplificación de la PCR obtenida con una pareja de oligonucleótidos). 2. Número de bandas polimórficas (np). 3. Número de bandas no polimórficas (nnp). 4. Número total de bandas (n= np+ nnp). 5. Número promedio de bandas polimórficas por unidad de ensayo (np/u). 6. Número de patrones de bandas identificados por combinación de oligonucleótidos (Tp). 7. Número de patrones de bandas únicos identificados por combinación de oligonucleótidos (Tpu). 8. Porcentaje de identificación (Pi = (Tpu/Tp)x100). 9. Número promedio de patrones de bandas identificados (I). 10. Número promedio de patrones únicos de bandas identificados (Iu). 11. Heterocigosidad esperada (Hep) del loci polimórfico: Con estos datos de ausencia presencia de bandas también se desarrolló un análisis de agrupamiento, en el cual se tuvo en cuenta el valor cofenético más alto para la selección del coeficiente y del algoritmo de agrupamiento por emplear. Se utilizó el coeficiente de Jaccard (1908) entre cada par de accesiones y el método de las medias aritméticas por grupo no ponderadas (UPGMA, Unweighted Pair-group Method Arithmetic Averages) para la construcción del dendrograma. Se empleó el paquete estadístico NTSYS-pc (versión 2.1; Rohlf, 2001) para la realización del mismo. Se utilizó el método del remuestreo para confirmar la veracidad de los grupos, así como el cálculo de los valores de similitud entre posibles grupos con este mismo propósito. Resultados y discusión La evaluación de los genotipos de guayabo con las cinco combinaciones de cebadores ISTR analizadas generó un total de 173 bandas, de las cuales el 30% son polimórficas. Se detectaron de 5 a 17 bandas polimórficas para las combinaciones de cebadores F5/B3 y F3/ B2B, respectivamente (tabla 3). La cantidad de bandas detectadas corrobora lo referido en la literatura por diversos autores respecto a la eficiencia de esta técnica para generar un gran número de marcadores en un período de tiempo corto (Rohde, 1996; Capote, 2007), y para la identificación de accesiones en el guayabo (Rodríguez et al., 2003). Las combinaciones F3/B3 y F3/B2B mostraron el mayor número de bandas, de las cuales el 60% y el 55% respectivamente fueron polimórficas. La combinación F1/B3, aunque mostró un menor número de bandas polimórficas, tiene un nivel de polimorfismo (53%) comparable a las dos mencionadas anteriormente. Las restantes parejas (F5/B3 y F3/B10) mostraron valores bajos de polimorfismo (25% y 23%, respectivamente) (tabla 3). Los cebadores ISTR evaluados permitieron generar un total de 52 fragmentos polimórficos con los cuales fue posible diferenciar todos los materiales analizados. Resultados similares fueron obtenidos por Ramírez et al. (2003), Rodríguez et al. (2003), Dueñas (2005), Capote (2007) y Alonso et al. (2008) durante el estudio de accesiones de aguacatero (Persea americana Mill.), guayabo, género Musa, mango y cocotero (Cocos nucifera L.), respectivamente, empleando este mismo marcador molecular. El 100% de los patrones de bandas identificados por las combinaciones F3/B3 y F3/ B2B fueron únicos (tabla 3), por lo que resultaron altamente eficientes para la diferenciación de todos los genotipos analizados y son recomendables, por tanto, para este tipo de análisis en el guayabo. El resto mostraron valores de intermedios a altos de porcentaje de identificación, a pesar del bajo número de bandas polimórficas detectado para algunas de ellas. De manera general, la heterocigosidad esperada fue baja para todas las combinaciones evaluadas, lo cual influyó en el valor medio detectado (tabla 3). Esto está en correspondencia con la naturaleza fundamentalmente autógama que tiene el guayabo, además de que puede constituir un reflejo del grado de parentesco que existe entre muchas de estas accesiones (Subramanyan e Iyer, 1993). Resultados similares fueron referidos por Valdés-Infante (2009) en la caracterización de un mayor número de accesiones pertenecientes a la colección cubana de guayabo, así como por Alonso et al. (2008) durante el estudio de accesiones de cocotero y por Capote (2007) en mango. Las combinaciones con los valores más altos fueron F3/B3 y F3/B2B (0,32 y 0,31 respectivamente), lo que es de esperar si se tiene en cuenta que mostraron los mayores valores de porcentaje de discriminación y de polimorfismo (tabla 3). Teniendo en cuenta lo comentado, son recomendables no sólo para la identificación varietal sino para la realización de estudios de diversidad en el cultivo. Por otra parte, la figura 1 muestra las relaciones genéticas entre las diferentes accesiones estudiadas. En la misma se puede apreciar la formación de cuatro grupos de diversidad definidos y la presencia de cuatro accesiones externas (EEA 18-40, BG 76-23, Peru roja y Microguayaba). Los valores de similitud entre grupos variaron del 28% al 36%, confirmando la veracidad de los mismos. Rodríguez et al. (2003) obtuvieron resultados similares al observar la formación de dos grupos de diversidad en la caracterización de descendientes y parentales del cruce Enana Roja Cubana x Bangkok utilizando marcadores ISTR. En el grupo I se encuentra la accesión Suprema roja, introducida de la Florida, agrupada con los dos materiales locales Belic L-123 y EEA 1-23. Según Cañizares (1981), en los inicios del mejoramiento genético de esta especie en el país se seleccionaron frutos con características deseables del cultivar Indian pink, cuyas semillas se sembraron y sus descendientes fueron designados con las letras Belic, seguidas de un número. Por otra parte, este mismo autor señala que la accesión EEA 1-23 también se deriva de la Indian pink, por lo que no resulta sorprendente que ambos genotipos estén formando un grupo. Los mismos fueron obtenidos a partir de polinización abierta, por lo que se desconoce el otro posible parental. No obstante, estos resultados contrastan con los obtenidos por AFLP (Valdés-Infante et al., 2003; Rodríguez et al., 2004) y por SSR (Rodríguez et al., 2005) en el estudio de diversidad de este mismo banco de germoplasma, donde la Suprema Roja se encuentra separada de la EEA 1-23 (AFLP) o en el grupo contiguo a la misma (SSR). De igual forma, en el grupo II se encuentran las accesiones locales BG 73-7 y EEA 6-19 con Seychelles, procedente de Islas Seychelles, y con N7 sin semillas, una mutación somática del cultivar N7 introducido desde la Florida a finales de la década de los cincuenta, según lo referido por Cañizares (1981). Respecto a la asociación de EEA 6-19 con Seychelles, se obtuvieron estos mismos resultados utilizando combinaciones de cebadores SSR, lo cual contrasta con lo observado por AFLP, donde la EEA 6-19 es una accesión externa en el dendrograma (Valdés-Infante et al., 2009). La agrupación de la N7 sin semillas con EEA 6-19 y Seychelles, dos genotipos con características moleculares y morfoagronómicas diferentes, confirma la naturaleza distintiva de estos materiales y sugiere la necesidad de preservarlos en la colección por representar una fuente peculiar de variabilidad. Por otra parte, la asociación de estos genotipos con la BG 73-7 no mostró igual relación en el análisis con AFLP y SSR, donde se ubicó bien separada de estas accesiones (Valdés- Infante, 2009). Resulta interesante destacar la localización de Cotorrera, Ibarra y Homero No. 1 en el grupo III. Los tres son genotipos locales y procedentes de prospecciones realizadas en diferentes regiones del país. Cotorrera es la forma silvestre del guayabo en Cuba; mientras que Ibarra es una mutación somática del cultivar N6, introducido desde la Florida. La misma fue identificada en la región de Sancti Spíritus, por el mejorador Juan Ibarra, según lo referido por Cañizares (1981). Esta hipótesis fue corroborada por Rodríguez et al. (2005) y Valdés-Infante (2009) con la utilización de los AFLP y los SSR. La caracterización molecular de este germoplasma con estos otros dos marcadores moleculares también coincide en la asociación de Ibarra y Homero No.1, no siendo así para el caso de la Cotorrera (Rodríguez et al., 2005; Valdés-Infante, 2009). No obstante, este genotipo se encuentra ampliamente distribuido por todo el país, por lo que no resulta sorprendente que pueda estar compartiendo información con accesiones procedentes de prospecciones locales. El grupo IV está formado por los genotipos locales BG 76-6, BG 73-10, Belic L-120, Belic L-207, BG 73-8, y la accesión introducida de la Florida N6. En la década de los sesenta se sembraron semillas de frutos extraordinarios de los cultivares N6 y Suprema roja, los cuales fueron designados con las letras BG, según lo referido por Cañizares (1981). En este grupo se pueden encontrar algunos de estos representantes que pudieran ser hijos de N6. Por otra parte, los denominados Belic son procedentes de polinización abierta, aún cuando se conoce que su progenitor femenino es Indian pink (Cañizares, 1968). De forma general, el agrupamiento de estas accesiones se corresponde con el obtenido por SSR y, en algunos casos, con el de AFLP en la caracterización previa de este banco de germoplasma, excepto para el caso de N6 (Rodríguez et al., 2005; Valdés- Infante, 2009). Con relación a las accesiones externas, nuevamente la Microguayaba se ubica separada del resto, resultado que coincide con el referido por Valdés-Infante et al. (2003), Rodríguez et al. (2004) y Rodríguez et al. (2005) con la utilización de marcadores AFLP, caracteres morfoagronómicos y cebadores SSR, respectivamente. Esto corrobora la hipótesis de que esta accesión pueda ser una subespecie de Psidium guajava L. (Valdés-Infante, 2009). De igual modo, Peru roja fue prospectada en la Isla de la Juventud, y por sus características morfológicas distintivas ha sido utilizada en los programas de mejoramiento genético de esta especie, para la selección de genotipos promisorios procedentes de polinización abierta (Cañizares, 1968 y 1981). Por otra parte, BG 76-23 también fue identificada como una accesión externa cuando se emplearon cebadores SSR en la caracterización de este germoplasma (Rodríguez et al., 2005). A su vez, EEA 18-40 o Enana Roja Cubana se relacionó con un conjunto de accesiones locales tanto por AFLP como por SSR (Valdés-Infante et al., 2003; Rodríguez et al., 2005), resultado que contrasta con el encontrado en el presente estudio. Sin embargo, esta accesión reúne un conjunto de características morfológicas distintivas que han hecho de ella el principal cultivar en explotación comercial y el patrón por modificar en los programas de mejoramiento. Por tanto, no resulta sorprendente que pueda portar información genética diferente. Las diferencias encontradas en el agrupamiento de las accesiones por ISTR respecto a las obtenidas previamente por AFLP y SSR sugieren que el elegir la técnica más apropiada para determinados estudios puede resultar un proceso difícil, por lo que a menudo se hace necesario una combinación de las mismas para obtener la información que se desea, lo que coincide con lo referido por Mohammadi y Prasanna (2003). Lo comentado en este trabajo indica la necesidad de realizar estudios integrados en el banco de germoplasma de este cultivo para lograr un manejo más adecuado del mismo. Esto garantiza un uso más racional de la base genética del guayabo para futuras generaciones, teniendo en cuenta el alto grado de parentesco que existe entre la mayoría de las accesiones presentes en la colección; por lo que la obtención de grupos de diversidad que orienten los programas de cruzamientos contribuye a aumentar la variabilidad y a disminuir el riesgo de erosión genética en la especie. Conclusiones La técnica ISTR constituye una herramienta de gran la utilidad para la identificación de accesiones y el análisis de la diversidad en el guayabo. La identificación de accesiones externas con características distintivas pone en evidencia la utilidad de los ISTR para la selección de material de interés para conservar por representar una fuente peculiar de variabilidad. La integración de la información obtenida por ISTR, con la referida previamente para los marcadores AFLP y SSR, garantizará un manejo más adecuado del banco de germoplasma y de la diversidad existente para futuras generaciones. Referencias bibliográficas 1 Aitken, K., J. Botero., R. Zwart., R. Teasdale. 1998. Detection of genetic diversity using RAPD markers in the genus Melaleuca. Acta. Hortc (ISHS) 461: 209-218. [ Links ] 2 Alonso, M., J. R. Cueto, Y. Santos, R. Llauger, M. Rodríguez, W. Rohde. 2008. Estimación de la variabilidad genética entre ecotipos de cocotero presentes en Cuba por ISTR. Rev Colomb Biotecnol X (2): 6-13. [ Links ] 3 Aranguren, Y., G. Fermín. 2007. Variabilidad de guayabas nativas en Venezuela por marcadores moleculares y UPOV. II simposio Internacional Fruticultura Tropical y Subtropical 2007. 8 p. [ Links ] 4 Belaj, A., Z. Satovis, G. Cipriano; L. Baldono; R. Testolin; L. Rallo, I. Trujillo. 2003. Comparative study of the discriminating capacity of RADP, ALP and SSR markers and of their effectiveness in stablishing genetic relationships in olive. Theor Appl Genet 107: 736-744. [ Links ] 5 Cañizares, J. 1981. Mejoramiento de la guayaba (Psidium guajava L.) por selección masal. Ciencia y Técnica en la Agricultura 4 (3-4): 7-23. [ Links ] 6 Cañizares, Z. J. 1968. La guayaba y otras frutas Myrtáceas. Ed. Revolucionaria. Instituto Cubano del Libro. 87 p. [ Links ] 7 Capote, M. 2007. Caracterización morfoagronómica y molecular de variedades comerciales de mango (Mangifera indica L.) en Cuba. Tesis de Maestro en Ciencias. Facultad de Biología. UH. 114 p. [ Links ] 8 Coêhlo de Lima, M. A., Simâo de Assis, J., Gonzaga, L. 2002. Caracterizacâo dos frutos de goiabeira e selecâo de cultivares na regiâo do submédio Sâo Francisco. Rev Bras Frutic 24 (1): 273-276. [ Links ] 9 Dettori, M. T., M. T. Palombi. 2000. Identification of Feijoa sellowiana Berg accessions by RAPD markers. Scientia Horticulturae 86: 279-290. [ Links ] 10 Doyle, J. J., J. L Doyle. 1990. Isolation of plant DNA from fresh tissue. Focus 12 (1): 13-15. [ Links ] 11 Dueñas, F. 2005. Determinación de la variabilidad genética en clones y somaclones de bananos y plátanos del género Musa. Tesis en opción al Título de Máster en Biología Vegetal. UH. 75 p. [ Links ] 12 Frankham, R., J. D. Ballou, D. A. Briscoe. 2003. Introduction to Conservation Genetics. Cambridge: Cambridge University Press. 100 p. [ Links ] 13 Hernández, D. S., J. Martínez., S. Padilla, N. Mayek. 2003. Diversidad genética de Psidium sp en la región Calvillo- Cañonnes, México. Primer Simposio Internacional de la guayaba. p. 71-83. [ Links ] 14 Hernández-Delgado, S., J. S. Padilla-Ramírez, A. Nava- Cedillo, N. Mayek-Pérez. 2007. Morphological and genetic diversity of Mexican guava germplasm. Plant Genetic Resources 5: 131-141. [ Links ] 15 Jaccard, P. 1908. Nouvelles rescherches sur la dsitribution florale. Bull Soc Vaud Sci Nat 44: 223-270. [ Links ] 16Machery and Nagel. 2002. NucleoSpinR Extract 2 in 1 Protocol. Germany. p. 3-13. [ Links ] 17 Marcucci, S., C. Acuña, S. Torales, N. Zelener, P. Pathaver, G. López et al. 2001. Evaluación de la Variabilidad Genética en huertos semilleros de especies de Eucalyptus. http://www.inta.gov.ar/ediciones/idia/forest/genetica10.pdf 7 p. [ Links ] 18Mohammadi, S. A., B. M. Prasanna. 2003. Analysis of genetic diversity in crop plants – Salient statistical tools and considerations. Crop Sci 43: 1235-1248. [ Links ] 19 Ramírez, I. M., N. N. Rodríguez, J. Valdés-Infante, M. Capote; D. Becker, W. Rohde. 2004. Isolation of genomic DNAs from the tropical fruit trees avocado, coconut, guava and mango for DNA marker application. Cultivos Tropicales 25: 33-38. [ Links ] 20 Ramírez, I., J. L. Rodríguez, N. N. Rodríguez, J. R. Cueto, W. Rohde. 2003. DNA polymorphism in Cuba varieties of avocado (Persea americana Mill.) as detected by Inverse Sequence Tagged Repeat (ISTR). Cultivos Tropicales 23 (3): 75-85. [ Links ] 21 Reveles, L. R., L. A. Saenz, E. Esparza, F. J. Cabral. 2003. Polimorfismo de ADN genómico en 12 selecciones de guayabo (Psidium guajava L.) del banco de germoplasma del campo experimental Los Cañones. Primer Simposio Internacional de la guayaba. p. 248- 252. [ Links ] 22Rodríguez, N. N., J. Valdés-Infante., D. Becker, B. Velázquez, O. Coto, E. Ritter, W. Rohde. 2004. Morphological, agronomic and molecular characterization of guava accessions (Psidium guajava L.) in Cuba. Journal of Genetic and Breeding 58 (79-90). [ Links ] 23 Rodríguez, N. N., Valdés-Infante, J., Becker, D., Velázquez, B., González, G., Sourd et al. 2005. Characterization of guava accessions by SSR markers, extension of the molecular linkage map, and mapping of QTLs for vegetative and reproductive characters. In Pathak, R. K., Singh, G., Kishun, R., Chandra, E. (eds.). Proceedings of the First International Guava Symposium, Lucknow, India, December 5-8, 2005. Louvain: ISHS Belgique, p. 201-215. International Guava Symposium. 1, 2005-12-05/2005-12-08, Lucknow, Inde. [ Links ] 24 Rodríguez, N. N., G. González, A. Simón, R. Jiménez, O. Mas, M. Morenza. 1999. Recursos genéticos del aguacatero (Persea americana Mill.) en Cuba. I. Prospección, colecta, establecimiento de la colección, selección y caracterización de cultivares. 17 (1-2-3): 27-32. [ Links ] 25 Rodríguez, N. N., J. Valdés-Infante, W. Rohde, D. Becker, G. González, V. Fuentes, B. Velásquez, D. Sourt. 2003. Molecular and morph-agronomic characterization of guava (Psidium guajava L.) hybrids population. Taller Internacional sobre Biotecnología Biotecnología Vegetal BioVeg 2003. p. 56-65. [ Links ] 26 Rohde, W. 1996. Inverse sequence tagged repeat (ISTR) analysis: a novel and universal PCR-based technique for genome analysis in the plant and animal kingdom. J Genet And Breed 50: 249-261. [ Links ] 27 Rohlf, F. J. 2001. NTSYS-PC, numerical taxonomy and multivariate análisis system. Version 2.1. Exeter Software, Setauket, New York. http://www.exetersoftware.com/cat/ntsyspc.htlm. 30 p. [ Links ] 28 Rossetto, M., F. C. Harris, A. McLauchlan, R. J. Henry, P. R. Baverstock, L. S. Lee. 2000. Interspecific amplification of tea tree (Melaleuca alternifolia-Myrtaceae) micosatellite loci-potential implications for conservation studies. Australian Journal of Botany 48: 367-373. [ Links ] 29 Rueda, A. 2003. Caracterización molecular del banco de germoplasma de Guayaba Psidium spp. del Centro de Investigación de Corpoica Palmira. Trabajo de Grado (Ing. Agr.) Universidad Nacional de Colombia. 65 p. [ Links ] 30 Sanabria, H., M. García, H. A. Díaz, J. Muñoz. 2005. Caracterización morfológica en árboles nativos de guayaba del valle del Cauca. Acta Agronómica 54 (4): 1-8. [ Links ] 31 Sanabria, H., M. A. García, J. E. Muñoz, H. A. Díaz. 2006. Caracterización molecular con marcadores RAM de árboles nativos de Psidium guajava (guayaba) en el Valle del Cauca. Acta Agronómica. 55 (1): 1-8. [ Links ] 32 Sharma, A. S., S. K. Sehrawat, R. S. Singhrot, K. S. Boora. 2005. Assessment of genetic diversity and relationship among Psidium spp. through RAPD analysis. In Pathak R. K. Singh, G., Kishun, R., Chandra E. (eds.). Proceedings of the First International Guava Symposium, Lucknow, India, December 5-8, 2005. Louvain: ISHS Belgique, p. 216-224. International Guava Symposium. 1, 2005-12-05/2005-12-08, Lucknow, Inde. [ Links ] 33 Subramanyan, M. D., C. P. A. Iyer. 1993. Improvement of Guava. In Advances in Horticulture. Vol. 1-Fruits Crops Part 1. New Delhi: Malhotra Publishing House. p. 337-347. [ Links ] 34 Tapia, E., M. A. Gutiérrez, M. Warburton, A. Santacruz, á. Villegas. 2005. Characterization of madarin (Citrus spp.) using morphological and AFLP markers. INCI 30 (11): 1-23. [ Links ] 35Valadez, M. E., K. Günter. 2000. Huellas de AND en genomas de plantas. Teoría y protocolos de Laboratorio. Mundi-Prensa México, S.A de C.V. 60 p. [ Links ] 36 Valdés-Infante, J. 2009. Utilización de caracteres morfoagronómicos y de marcadores de ADN para el desarrollo de una metodología que contribuya al mejoramiento genético del guayabo (Psidium guajava L.) en Cuba. Tesis presentada en opción al Grado Científico de Doctor en Ciencias Biológicas. 120 p. [ Links ] 37 Valdés-Infante, J., N. N. Rodríguez. 2003. Marcadores moleculares de ADN y su aplicación en los frutales tropicales. En: fao.cubasi.cu. Sitio Web de la FAO. 25 p. [ Links ] 38 Valdés-Infante, J., D. Becker, N. Rodríguez; B. Velásquez, G. González, D. Sourd et al. 2003. Molecular characterization of Cuban accessions of guava (Psidium guajava L.), establishment of a first molecular linkage map and mapping of QTLs for vegetative characters. J Genet and Breed 57: 349-358. [ Links ] 39 Vasco, N. L., J. S. Padilla, J. Toro. 2003. Composición nutrimental de la guayaba y sus semillas. Primer Simposio Internacional de la guayaba. p. 116-123. [ Links ] 

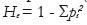 donde pi es la frecuencia de alelos en el imo alelo y la media aritmética de la heterocigosidad del loci polimórfico:
donde pi es la frecuencia de alelos en el imo alelo y la media aritmética de la heterocigosidad del loci polimórfico: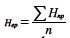 , donde n es el número de marcadores analizados.
, donde n es el número de marcadores analizados.