










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.12 n.1 Bogotá jan./jun. 2010
ARTÍCULOS CORTOS
Efecto de adición de iones hierro y zinc sobre la producción de etanol de dos cepas recombinantes de Saccharomyces cerevisiae
The effect of adding iron and zinc ions to ethanol production from two recombinant Saccharomyces cerevisiae strains
Carolina Peña Serna1 , Rafael Arango Isaza2 , Luis Fernando Restrepo Betancur3
1Ing, de Procesos, MSc. Facultad de Ciencias Agrarias, Universidad de Antioquia. Corporación para Investigaciones Biológicas. Carrera 75#65-87. cpenaser@gmail.com
2Médico, PhD. Corporación para Investigaciones Biológicas. Carrera 72A#78B-141. rarango@cib.org.co
3Estadístico, Esp. Facultad de Ciencias Agrarias, Universidad de Antioquia. Carrera 75#65-87. fercho7474@hotmail.com
Recibido: Recibido: febrero 12 de 2010 Aprobado: junio 23 de 2010
Resumen
La producción de etanol por fermentación es influenciada por la presencia de iones metálicos como hierro y zinc dado que son cofactores de la enzima alcohol deshidrogenasa. El estudio de este efecto permitiría identificar el comportamiento de los microorganismos fermentadores en sustratos industriales que contienen altas concentraciones de este tipo de iones. Este trabajo evaluó la producción de biomasa, los azúcares residuales y la producción de etanol por fermentación de tres cepas de S. cerevisiae, CBS8066, recombinantes GG570-CIBI y GG570-CIBII, bajo el efecto de la adición de hierro a 0, 50 y 150 µM, y zinc a 0 y 50 µM. Las cepas presentaron inhibición en la producción de biomasa y etanol bajo efecto de iones de hierro y zinc, siendo dicha inhibición mayor al estar en presencia de zinc o alta concentración de hierro. GG570-CIBI mostró disminución en producción de biomasa de 4 g/L y una caída en producción de etanol de 40% en el tratamiento 150 µM hierro-50 µM zinc (con respecto al tratamiento basal). GG570-CIBII fue la menos afectada con inhibición en la producción de etanol inferior a 11% a las 20 h de fermentación. Adicionalmente, presentó la mayor producción de etanol cuando hubo adición de 150 µM Fe con o sin adición de zinc, siendo dicha producción entre un 9 y 14% superior a la de las cepas CBS8066 y GG570-CIBI respectivamente, bajo las mismas condiciones. Posteriormente, GG570-CIBII será evaluada en sustratos industriales debido a su menor inhibición en la producción de etanol, permitiendo así obtener mejores rendimientos.
Palabras clave: gen pdc, gen adhII, Zymomonas mobilis, ión ferroso, ión zinc.
Abstract
Ethanol production by fermentation is influenced by the presence of metallic ions such as iron and zinc because these are alcohol dehydrogenase enzyme cofactors. Studying this effect leads to identifying the behaviour of microorganisms in industrial substrates containing high concentrations of these kinds of ions. This work therefore evaluated biomass production, residual sugars and ethanol production by fermenting three S. cerevisiae strains (CBS8066, recombinants GG570-CIBI and GG570-CIBII) under the effect of adding ferrous ion at 0, 50 and 150 µM and zinc ion at 0 and 50 µM. The strains had inhibited biomass and ethanol production when zinc and ferrous ions were added; such inhibition was greater in the presence of zinc or iron having higher concentrations. GG570-CIBI had 4 g/L reduction in biomass production and ethanol production dropped by 40% in the 150 µM iron–50 µM zinc treatment (compared to base treatment). GG570-CIBII was the less affected, having less than 11% inhibition of ethanol production at 20 h fermentation. GG570-CIBII presented the greatest ethanol production when 150 µM iron was added to the culture medium with or without adding zinc; in this case, production was 9% and 14% greater than CBS8066 and GG570-CIBI ethanol production, respectively, in the same conditions. GG570-CIBII will be evaluated later on using industrial substrates due to its lower ethanol production inhibition, thereby leading to better yields being obtained.
Key words: pdc gene, adhII gene, Zymomonas mobilis, ferrous ion, zinc ion.
Introducción
El bioetanol es un producto derivado de la fermentación alcohólica de los azúcares presentes en materiales renovables como caña de azúcar, remolacha azucarera, madera, maíz, entre otros (Federación Nacional de Biocombustibles, 2007; Soliclima, 2006), dicha fermentación es realizada por microorganismos como Saccharomyces cerevisiae o Zymomonas mobilis.
Industrialmente, el bioetanol se produce utilizando S. cerevisiae como microorganismo fermentador debido a la capacidad para metabolizar un espectro amplio de azúcares (Ertola et al., 2007) y tolerar minerales presentes en sustratos de uso industrial (Gunasekaran y Chandra,1999). Sin embargo, Z. mobilis ha mostrado tener un mayor rendimiento en la producción de etanol (Gunasekaran y Chandra, 1999; Glick y Pasternak, 2003; Davis et al., 2006), posiblemente como consecuencia de la mayor afinidad de las enzimas piruvato descarboxilasa (PDC) EC 4.1.1.1 y alcohol deshidrogenasa (ADH) EC 1.1.1.1 sobre sus respectivos sustratos (Brenda database; Hoppner y Doelle, 1983).
Según lo reportado en Brenda database y Gunasekaran y Chandra (1999), las enzimas ADH de S. cerevisiae requieren ión zinc como cofactor enzimático, mientras la enzima ADHII de Z. mobilis requiere ión ferroso (Mackenzie et al., 1989; Gunasekaran y Chandra, 1999).
De acuerdo con lo reportado por Mackenzie et al. (1989), la adición de iones ferroso en concentraciones entre 2-50 µM y zinc en concentraciones menores a 50µ M incrementan la actividad catalítica de las enzimas ADHI y ADHII de Z. mobilis, potenciando principalmente la actividad catalítica de la ADHII.
Sustratos industriales usados para la producción de bioetanol como la melaza de caña de azúcar contienen altas concentraciones de hierro lo cual podría afectar de forma directa la producción de etanol, y cuyo efecto no ha sido evaluado hasta el momento.
Por esta razón, en este trabajo se evaluó la adición de iones zinc y ferroso sobre la producción de etanol de tres cepas de S. cerevisiae: CBS8066 (parental, utilizada como control) y dos recombinantes, GG570-CIBI y GG570-CIBII desarrolladas en la Unidad de Biotecnología Vegetal de la Corporación para Investigaciones Biológicas (CIB).
Materiales y métodos
Microorganismo de estudio
Se utilizaron tres cepas de Saccharomyces cerevisiae: cepa CBS8066 (parental) usada como control obtenida de Centraalbureau voor SchimMelcultures (Utrecht, Holanda), la cepa GG570-CIBI que contiene el gen pdc de Z. mobilis bajo el control del promotor y terminador de la fosfoglicerato kinasa (PGK) de S. cerevisiae, y la cepa GG570-CIBII que contiene el gen pdc con el promotor y terminador de PGK y el gen adhII de Z. mobilis, bajo el control del promotor y terminador de la alcohol deshidrogenasa I de S. cerevisiae (Vásquez et al., 2007).
Las cepas de S. cerevisiae fueron mantenidas en agar YPD (1% extracto de levadura, 2% peptona y 2% glucosa) a 4° C (Suresh, et al., 1999; Blanco et al., 2006).
Medio de cultivo
Se utilizó el medio de cultivo YPD modificado con la siguiente composición: glucosa 100 g/L, KH2PO4 1 g/L, MgSO4*7H2O 0.5 g/L, peptona 10 g/L y extracto de levadura 5 g/L. De acuerdo con los tratamientos por evaluar, se adicionó hierro y zinc a través de FeSO4*7H2O y ZnSO4*7H2O, respectivamente.
Adicionalmente, al medio YPD modificado se le realizó un análisis químico por absorción atómica para determinar la concentración inicial de hierro y zinc.
Experimentos fermentativos
Se preparó un preinóculo con una colonia de S. cerevisiae aislada del agar YPD, se cultivó en 50 mL de medio de cultivo líquido YPD modificado 10% glucosa por 24 h, luego se midió DO660 (Davis et al., 2006; Govindaswamy y Vane, 2007) hasta alcanzar 1,6 y posteriormente se inocularon los frascos de fermentación con 2% v/v del preinóculo (Mojovic et al., 2006; Govindaswamy y Vane, 2007).
Se realizaron fermentaciones anaeróbicas por duplicado durante un periodo de 20 h, en frasco de 250 mL con 100 mL de medio de cultivo, en agitador orbital bajo condiciones controladas de temperatura 30 °C (Converti et al., 1985; Govindaswamy y Vane, 2007), velocidad de agitación 150 rpm (Suresh et al., 1999; Karimi et al., 2006) y pH inicial 5,0 ± 0,1 (Converti et al., 1985; Mojovic et al., 2006), evaluando las cepas CBS8066, GG570-CIBI y GG570-CIBII bajo el efecto de las concentraciones del ión ferroso (FeSO4*7H20) 0, 50 (Mackenzie et al., 1989) y 150 µM, y las concentraciones del ión zinc (ZnSO4*7H20) 0 y 50 µM (Mackenzie et al., 1989). La toma de muestras se realizó a las 0, 16 y 20 h por duplicado. Estos tiempos fueron establecidos a partir de un trabajo previo realizado por Vásquez et al. (2007) en el cual se determinó que los picos máximos de producción de etanol de la cepa GG570-CIBII se presentaron a las 16 y 20 h de fermentación.
Evaluación de la biomasa celular
Se hizo seguimiento a la biomasa por el método de peso seco. De cada fermentación se tomó 1 mL de suspensión celular en un tubo eppendorff de 1,5 mL, se sometió a centrifugación a 12000 rpm y 4° C durante 6 min, luego se descartó el sobrenadante y se procedió a resuspender el pellet en agua destilada para lavarlo y eliminar azúcares residuales. Posteriormente se volvió a centrifugar, se descartó el sobrenadante y se sometió el pellet húmedo a secado en horno a 80° C por 24 h o hasta que el peso fuera constante. Dicho procedimiento se realizó para cada una de las muestras tomadas.
Evaluación de los azúcares residuales
El contenido de azúcares residuales de la fermentación se determinó por el método del DNS (Miller, 1959; Mojovic et al., 2006, Zhu et al., 2006).
Evaluación de la concentración de etanol
Se cuantificó por cromatografía de gases (Zhu et al., 2006; Govindaswamy y Vane, 2007; Plessas et al., 2007) en un equipo marca Agilent modelo 6890 con detector de masas y automuestreador.
Análisis estadístico
El experimento fue realizado con un diseño de clasificación en bloques aleatorizados, con arreglo factorial asimétrico 2x3 balanceado, de efecto fijo en el tiempo e intratiempo, con submuestreo, dos repeticiones y medidas repetidas a las 0, 16 y 20 h; donde las variables de respuesta fueron producción de biomasa, azúcares residuales y producción de etanol.
Dicho análisis se articuló a la estructura Manova con contraste canónico ortogonal, completándose con análisis Anova individuales en cada variable de respuesta, y contrastándose con la prueba de Tukey a un nivel de significancia de 5%. Adicionalmente, se complementó con estadística descriptiva unidimensional para cada tiempo, tratamiento y niveles de cofactores. Para realizar los análisis mencionados se utilizó el paquete estadístico SAS versión 9.1.tabla 1
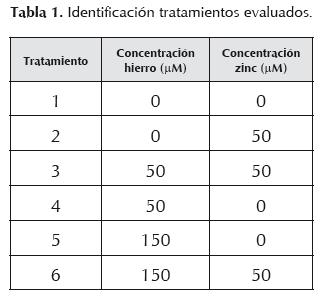
Resultados
La concentración basal o inicial de los iones metálicos del medio YPD modificado (tratamiento 1) arrojó un valor <1 ppm para hierro y 0,004% para zinc.
Evaluación de la biomasa celular
En la figura 1 se puede observar que las cepas de S. cerevisiae, cepa CBS8066, la cepa GG570-CIBI y la cepa GG570-CIBII produjeron menor cantidad de biomasa cuando hubo adición de iones de hierro y zinc.
De acuerdo con el análisis estadístico Anova, se determinó que el factor cepa tiene efecto estadísticamente significativo (p<0,05) sobre la producción de biomasa y, además, hubo diferencias altamente significativas (p<0,001) entre el tratamiento 1 con respecto a los tratamientos 2, 3, 5 y 6. De igual forma, el tratamiento 4 fue estadísticamente diferente a los tratamientos 2 y 6.
También, el tratamiento 3 presentó diferencia con respecto al tratamiento 2; siendo el tratamiento 6 (150 µM hierro-50 µM zinc) el menos productor de biomasa y el tratamiento 1 (0 µM hierro-0 µM zinc) el más productor, tal como se había mostrado anteriormente.
Evaluación de los azúcares residuales
En la figura 2 se puede observar que los azúcares residuales presentes en el medio de fermentación utilizado para las cepas de S. cerevisiae fueron similares bajo todos los tratamientos evaluados, sin embargo, la cepa recombinante GG570-CIBI presentó diferencias estadísticamente significativas (p<0,05) con respecto a la cepa control y la recombinante GG570-CIBII.
Evaluación de la producción de etanol
Como se puede observar en la figura 3, hubo disminución en la producción de etanol cuando se adicionaron iones metálicos al medio de cultivo (con respecto al tratamiento basal) para las cepas CBS8066 (parental) y recombinantes GG570-CIBI y GG570-CIBII, de igual forma a lo sucedido con la producción de biomasa.
Se encontraron diferencias estadísticas altamente significativas (p<0,001) entre el tratamiento 1 y los tratamientos 2, 3, 5 y 6. De igual forma, los tratamientos 3 y 4 fueron estadísticamente diferentes a los tratamientos 5, 2 y 6.
Adicionalmente, las cepas control (CBS8066) y recombinante GG570-CIBII mostraron diferencia estadística altamente significativa (p < 0,001) con respecto a la cepa recombinante GG570-CIBI.
Finalmente, de acuerdo con el Anova, la cepa tuvo efecto estadísticamente significativo (p<0,05) sobre la producción de etanol.
Discusión
En este trabajo se evaluó el efecto de la adición de iones zinc y ferroso sobre la producción de biomasa, los azúcares residuales y la producción de etanol de tres cepas de Saccharomyces cerevisiae, CBS8066 (control), G570- CIBI (recombinante con gen pdc de Z. mobilis) y GG570-CIBII (recombinante con genes pdc y adhII de Z. mobilis).
Evaluación de la producción de biomasa
Durante el proceso fermentativo las cepas de S. cerevisiae mostraron un comportamiento normal para microorganismos en el cual, a mayor tiempo de cultivo, mayor crecimiento celular (Converti et al., 1985; Thatipamala et al., 1992; Blanco et al., 2006), sin embargo, se pudo observar una disminución de la biomasa con la adición de iones metálicos.
De acuerdo con los resultados mostrados en la figura 1, la cepa control fue la menos afectada por la adición de iones, con una reducción de la biomasa inferior a 1 g/L, exceptuando el tratamiento con 150 µM hierro-50 µM zinc (figura 1f), donde se observó disminución de 1,3 y 2,0 g/L a las 16 y 20 h respectivamente.
La cepa GG570-CIBI presentó la mayor reducción en biomasa celular cuando se encontró bajo el efecto de la adición de los iones metálicos, disminuyendo el crecimiento en 3 y 4 g/L a las 20 h de fermentación en los tratamientos 150 µM hierro-0 µM zinc (figura 1e) y 0 µM hierro-50 µM zinc (figura 1b) respectivamente.
De igual forma, la cepa recombinante GG570-CIBII presentó disminución en la biomasa alcanzando una reducción máxima de 2,6 g/L a las 20 h en el tratamiento 150 µM hierro-50 µM zinc (figura 1f) con respecto al tratamiento sin adición de iones (figura 1a).
En el estudio realizado por Mackenzie et al. (1989) se reportó la disminución en el crecimiento celular de Z. mobilis bajo el efecto de la adición de zinc, la cual fue mayor que cuando hubo adición del ión ferroso, de igual forma a como se observó en el presente estudio con la cepa GG570-CIBII de S. cerevisiae que también codifica para la enzima ADHII.
Asimismo, se puede observar que cuando se incrementa la concentración del ión ferroso, ya sea en presencia o ausencia de zinc, las cepas reducen la biomasa celular, aun cuando dicha reducción es mayor en presencia de zinc.
De acuerdo con los resultados anteriores, se podría pensar en una disminución en el crecimiento celular por parte de S. cerevisiae cultivada en medios complejos como melaza de caña de azúcar o banano de rechazo (Hahn et al., 2005) debido a la presencia de iones metálicos como hierro y zinc, que en el caso de melaza se encuentran alrededor de 54,3-138 ppm y 0-7 ppm respectivamente, dependiendo de la procedencia de la melaza.
Evaluación de la producción de etanol
Según los resultados, la adición de zinc (figura 3b) podría tener un efecto inhibitorio sobre la producción de etanol, afectando en mayor proporción la cepa recombinante GG570-CIBI seguida de la cepa CBS8066. Por ello, se considera que la adición del ión zinc no es recomendable para incrementar la producción de etanol como se esperaba, al ser este ión un cofactor enzimático de las isoenzimas alcohol deshidrogenasa, propias de S. cerevisiae.
Mackenzie et al. (1989) reportaron disminución en la actividad catalítica alcohol deshidrogenasa de Z. mobilis al estar bajo el efecto del ión zinc, y aunque no reportaron producción de etanol, ésta tuvo que haber disminuido dado que la enzima ADH tiene un efecto directo sobre la producción de etanol.
Por otro lado, cuando hubo adición de zinc y además del ión hierro a baja concentración (figura 3d), se pudo observar que hay una recuperación de las cepas en cuanto a la producción de etanol. De igual forma, Mackenzie et al. (1989) reportaron incremento en la actividad catalítica de la enzima ADH de Z. mobilis cuando se adicionan ambos cofactores a la vez.
Se encontró que la adición del ión ferroso a una alta concentración, ya sea con o sin adición de zinc (figura 3e, 3f ) genera una disminución en la producción de etanol de las cepas de S. cerevisiae. Sin embargo, se puede ver claramente que cuando hay una concentración de 150 µM de ión ferroso, la cepa recombinante GG570-CIBII es la que presenta la mayor producción de etanol, siendo por tanto la menos inhibida y superando la producción de etanol de la cepa control y la recombinante GG570- CIBI en 9 y 14% respectivamente.
De acuerdo con lo reportado por Mackenzie et al. (1989) y Gunasekaran y Chandra (1999), el ión hierro es un cofactor de la enzima alcohol deshidrogenasa II (ADHII) presente en la bacteria Z. mobilis, que le permite incrementar la actividad enzimática mejorando así la producción de etanol. Dado que la cepa recombinante GG570-CIBII de S. cerevisiae tiene insertado el gen adhII que codifica para dicha enzima, el resultado obtenido muestra una ventaja metabólica adquirida por esta cepa al permitirle, bajo condiciones de alta concentración de hierro, producir la mayor concentración de etanol.
Adicionalmente, los sustratos utilizados para la fermentación alcohólica a nivel industrial como melaza de caña de azúcar tienen altas concentraciones de hierro, por lo cual la utilización de esta cepa permitiría mantener una buena producción de etanol con respecto a otras cepas de S. cerevisiae.
En la tabla 2, se muestran las reducciones en la producción de etanol de cada una de las cepas de S. cerevisiae bajo los tratamientos de adición de iones metálicos con respecto al tratamiento control que no tenía ninguna adición.
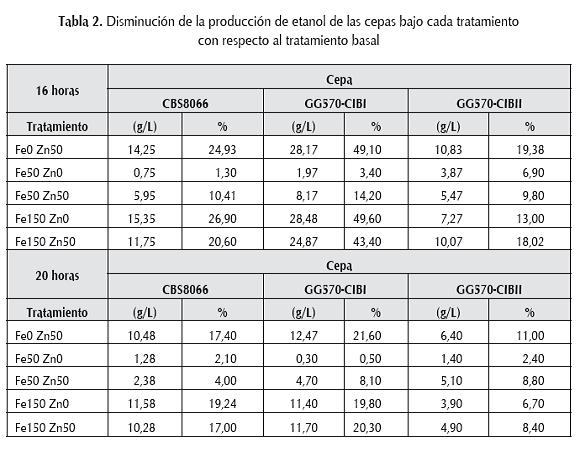
En general, se estableció que hubo mayor inhibición con los tratamientos 150 µM hierro- 0 µM zinc seguido por 0 µM hierro-50 µM zinc. Adicionalmente, la cepa que más disminuyó la producción de etanol fue la recombinante GG570-CIBI a las 16 horas de fermentación.
La cepa que es menos afectada por la adición de iones metálicos durante todo el tiempo de fermentación es la recombinante GG570-CIBII y, además, el tratamiento que más efecto negativo le generó fue 0 µM hierro-50 µM zinc. De acuerdo con esto, la adición de zinc al medio de fermentación podría causar inhibición de la enzima ADHII dado que todos los tratamientos con adición de este mineral (0 µM hierro-50 µM zinc, 50 µM hierro-50 µM zinc y 150 µM hierro-50 µM zinc) generaron una reducción mayor en las concentraciones de etanol que las encontradas en los tratamientos sin adición de éste (50 µM hierro-0 µM zinc y 150 µM hierro-0 µM zinc), igual a como sucede con Z. mobilis (Mackenzie et al., 1989).
En general, cuando el medio fermentativo contiene 50 µM hierro-50 µM zinc, las cepas disminuyeron en mayor proporción la producción de etanol con respecto al tratamiento sin adición de zinc (50 µM hiero–0 µM zinc), en el cual no se presentó una disminución de etanol tan grande, esto podría mostrar que el ión zinc tiene un efecto negativo sobre la producción de etanol.
Sin embargo, cuando la concentración de hierro se incrementó hasta 150 µM, el efecto inhibitorio de la adición de zinc se reduce aun cuando la disminución persiste pero principalmente como consecuencia de la alta concentración de hierro.
Conclusiones
Se logró determinar que la adición de iones metálicos como hierro y zinc que actúan como cofactores enzimáticos de las enzimas alcohol deshidrogenasa presentes en las cepas de S. cerevisiae produjeron una disminución en el crecimiento y la producción de etanol.
La cepa recombinante GG570-CIBII sufrió menor inhibición con respecto a la cepa CBS8066 (control) y GG570-CIBI cuando se adicionaron los iones metálicos. Asimismo, cuando se incrementó la concentración del ión ferroso dicha cepa presentó un aumento en la producción de etanol con respecto a la cepa CBS8066. Esto muestra que la cepa recombinante GG570-CIBII podría ser usada en fermentación de sustratos industriales (con altos contenidos de hierro) dado que la producción de etanol no sería reducida tan fuertemente.
También se estableció que la adición del ión zinc tuvo un efecto mayor en la inhibición de la producción de etanol por parte de las cepas de S. cerevisiae evaluadas.
Referencias bibliográficas
1 Blanco, M., Peinado, A., Mas, J. 2006. Monitoring alcoholic fermentation by joint use of soft and hard modelling methods. Analytica Chimica Acta 556. pp. 364-373. [ Links ]
2 Brenda–the comprehensive enzyme information system. Disponible en: http://www.brenda.uni-koeln.de Fecha de consulta: 27 de agosto de 2009. [ Links ]
3 Cazetta, M., Celligoi, M., Buzato, J., Scarmino, J. 2007. Fermentation of molasses by Zymomonas mobilis: Effects of temperature and sugar concentration on ethanol production. Bioresource Technology 98: 2824-2828. [ Links ]
4 Converti, A., Perego, P., Lodi, A., Parisi, F., Del Borghi, M. 1985. A Kinetic Study of Saccharomyces strains: Performance at high sugar concentrations. Biotechnology and Bioengineering 27: 1108-1114. [ Links ]
5 Davis, L., Rogers, P., Pearce, J., Peiris, P. 2006. Evaluation of Zymomonas based ethanol production from a hydrolysed waste starch stream. Biomass and Bioenergy 30: 809-814. [ Links ]
6 Ertola, R., Yantorno, O., Mignone, C. Programa Regional de Desarrollo Científico y Tecnológico de la OEA. 2007. Disponible en: http://fai.unne.edu.ar/biologia/microind/levaduras.htm Fecha de consulta: 7 de junio de 2007. [ Links ]
7 Federación Nacional de Biocombustibles. ABC de los Alcoholes Carburantes. 2007. Disponible en: http://www.minminas.gov.co/minminas/sectores.nsf/2a84e89f4d73f130052567be0052c75a/ffdff1b298ac766a052571720071cfad?OpenDocument. Fecha de consulta: 27 de mayo de 2007. [ Links ]
8 Glick, B., Pasternak. 2003. J. Molecular Biotechnology. 3 edición. Washintong D.C.: ASM Press. [ Links ]
9 Govindaswamy, S., Vane L. 2007. Kinetics of growth and ethanol production on different carbon substrates using genetically engineered xylose-fermenting yeast. Bioresource Technology 98: 677-685. [ Links ]
10 Gunasekaran, P., Chandra, R. K. 1999. Ethanol fermentation technology – Zymomonas mobilis. Current Science 77: 56-68. [ Links ]
11 Hahn, B., Karhumaa, K., Larsson, U., Gorwa, M., Görgens, J. van Zyl, W. 2005. Role of cultivation media in the development of yeast strains for large scale industrial use. Microbial Cell Factories 4: 1-16. [ Links ]
12 Hoppner, T., Doelle, H. 1983. Purification and kinetic characteristics of pyruvate decarboxylase and ethanol dehydrogenase from Zymomonas mobilis in relation to ethanol production. European Journal of Applied Microbiology and Biotechnology 17: 152-157. [ Links ]
13 Karimi, K., Emtiazi, G., Taherzadeh, M. 2006. Ethanol production from dilute-acid pretreated rice straw by simultaneous saccharification and fermentation with Mucor indicus, Rhizopus oryzae and Saccharomyces cerevisiae. Enzyme and Microbial Technology 40: 138-144. [ Links ]
14 Mackenzie, K., Eddy, C., Ingram, L. 1989. Modulation of alcohol dehydrogenase isoenzyme levels in Zymomonas mobilis by iron and zinc. Journal of Bacteriology 171: 1063-1067. [ Links ]
15 Miller, G. L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31: 426- 428. [ Links ]
16 Mojovic, L., Nikolic, S., Rakin, M., Vukasinovic, M. 2006. Production of bioethanol from corn meal hydrolyzates. Fuel 85: 1750-1755. [ Links ]
17 Plessas, S., Bekatorou, A., Koutinas, A., Soupioni, M., Banat, I., Marchant, R. 2007. Use of Saccharomyces cerevisiae cells iµMobilized on orange peel as biocatalyst for alcoholic fermentation. Bioresource Technology 98: 860-865. [ Links ]
18 Soliclima Energía Solar. El etanol como biocombustible. 2006. Disponible en: http://news.soliclima.com/modules.phpname=Content&pa=showpage & pid=118. Fecha de consulta: 20 de mayo de 2007. [ Links ]
19 Suresh, K., Kiran, N., Venkateswer, L. 1999. Utilization of damaged sorghum and rice grains for ethanol production by simultaneous saccharification and fermentation. Bioresource Technology 68: 301-304. [ Links ]
20 Thatipamala, R., Rohani, S., Hill, G. A. 1992. Effects of high product and substrate inhibition on the kinetics and biomass and products yields during ethanol batch fermentation. Biotechnology and Bioengineering 40. p. 289-297. [ Links ]
21 Vásquez, J., Castaño, H, Marín, P., Rodríguez, E., Arango, R. 2007. Ingeniería genética en rutas metabólicas de Saccharomyces cerevisiae para incrementar la productividad de etanol. Memorias 6° Simposio de Alcoholes y Levaduras 2007-Sipal 07. Bogotá, junio de 2007. [ Links ]
22 Zhu, S., Wu, Y., Yu, Z., Zhang, X., Wang, C., Yu, F., Jin, S. 2006. Production of ethanol from microwave-assisted alkali pretreated wheat straw. Process Biochemistry 41: 869-873. [ Links ]

